










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Hematología, Inmunología y Hemoterapia
versión On-line ISSN 1561-2996
Rev Cubana Hematol Inmunol Hemoter vol.32 no.1 Ciudad de la Habana ene.-mar. 2016
ARTÍCULO DE REVISIÓN
Regulación del metabolismo del hierro: dos sistemas, un mismo objetivo
Iron metabolism regulation: two systems, one goal
Mariela Forrellat Barrios
Instituto de Hematología e Inmunología. La Habana, Cuba.
RESUMEN
La existencia humana está indisolublemente unida al hierro, que es parte de una amplia variedad de enzimas claves como catalasas, aconitasas, ribonucleótido reductasa, peroxidasas y citocromos, que explotan la flexibilidad de su química redox para ejecutar un elevado número de reacciones esenciales para la vida. El cuerpo humano ha evolucionado para conservar el hierro en diferentes formas, incluido su reciclaje después de la ruptura de los eritrocitos y la retención en ausencia de un mecanismo de excreción. El metabolismo del hierro está balanceado por dos sistemas regulatorios: uno sistémico basado en la hormona hepcidina y la proteína exportadora ferroportina, y el otro que controla el metabolismo celular través de las proteínas reguladoras de hierro (IRP) que se unen a los elementos de respuesta al hierro (IRE) de los ARNm regulados. Estos sistemas funcionan de modo coordinado lo que evita, tanto la deficiencia como el exceso del mineral.
Palabras clave: metabolismo del hierro, hepcidina, ferroportina, sistema IRE/IRP.
ABSTRACT
Human existence is indissolubly linked to iron, which is part of a wide variety of key enzymes such as catalase, aconitases, ribonucleotide reductase, peroxidases and cytochromes, exploiting the flexibility of its redox chemistry to run a large number of reactions essential for life. Human body has evolved to keep iron in different forms, including recycling after rupture of erythrocytes and the retention without excretion mechanism. Iron metabolism is balanced by two regulatory systems: one based on systemic hormone hepcidin protein export and ferroportin, and the other, which controls cell metabolism through the iron regulatory protein (IRP) binding to the mRNAs regulated iron regulatory elements (IRE). These systems work in a coordinated manner avoiding both deficiency and excess.
Keywords: iron metabolism, hepcidin, ferroportin, IRE/IRP system.
INTRODUCCIÓN
La importancia del hierro (Fe) en la salud humana es conocida desde la antigüedad. Los primeros reportes de su uso medicinal datan de las antiguas civilizaciones egipcia, hindú, griega y romana. En el siglo XVII fue usado para el tratamiento de la clorosis, enfermedad resultante de su deficiencia; pero no fue hasta 1932 que la importancia del Fe fue demostrada al probarse que el Fe inorgánico es imprescindible para la síntesis de la hemoglobina.1
Durante años, el interés nutricional en este mineral se focalizó en su importancia para la síntesis de hemoglobina y el transporte de oxígeno. Hoy se conoce que es esencial para muchos procesos bioquímicos, en especial para la producción del grupo hemo y de los centros Fe/S presentes en un sinnúmero de proteínas y enzimas vinculadas a procesos vitales.1,2
Los últimos años se han caracterizado por la intensa actividad de investigación en el campo del metabolismo férrico. Como resultado, se han producido importantes avances en el esclarecimiento de los mecanismos de control de la homeostasia, sistémica y celular, del mineral.
PARADOJAS DEL Fe
El Fe es un elemento que resulta paradójico. Aunque es un mineral muy abundante y un componente esencial para los organismos vivos, frecuentemente resulta un factor limitante en el medio, debido a que al entrar en contacto con el oxígeno forma óxidos insolubles que limitan su disponibilidad para ser utilizado por los organismos. Es por ello que evolutivamente se han desarrollado variados mecanismos celulares para captarlo en su forma biológicamente activa.1
La clave de la utilidad biológica del Fe es su habilidad de existir e interconvertirse en dos estados de oxidación: ferroso (Fe2+) y férrico (Fe3+). Esta característica le permite actuar como un agente catalítico redox al aceptar y donar electrones reversiblemente; un excelente ejemplo son las proteínas que componen la cadena transportadora de electrones. Sin embargo, esta misma propiedad lo convierte en un elemento peligroso, pues es la base de su toxicidad, que puede reaccionar con las especies reactivas del oxígeno formadas como resultado de la respiración celular y provocar la peroxidación de proteínas, del ADN y de los lípidos de membrana; estos efectos deletéreos se exacerban cuando hay sobrecarga del mineral pues solo se producen cuando el Fe está “libre” o en una forma anormal dentro de la célula.3
En el hombre el Fe se encuentra fundamentalmente unido a proteínas en forma de complejos, como las hemoproteínas (hemoglobina, mioglobina, las enzimas que contienen hemo) o los compuestos no hemo (flavin Fe enzimas, transferrina y ferritina). Esta forma de quelación es un mecanismo de controlar su reactividad y permiten su adquisición, transporte y almacenamiento intracelular.1,3
METABOLISMO DEL Fe Y SÍNTESIS DEL HEMO
El Fe es imprescindible para la síntesis de una multitud de proteínas y enzimas de las que es parte componente o que lo utilizan como cofactor. Casi las dos terceras partes del Fe corporal se encuentran en la hemoglobina; el 25 % está contenido en las reservas movilizables y el resto está unido a la mioglobina. Una fracción pequeña, pero significativa, se encuentra formando parte de la amplia variedad de enzimas relacionadas con el metabolismo oxidativo y otras funciones celulares.1
El Fe es continuamente reciclado y estrictamente conservado por el organismo. La clave del suministro y homeostasia sistémica del Fe radica en la regulación de los niveles plasmáticos del mineral. Así, en la deficiencia de Fe se afectan la capacidad de transportar electrones y el metabolismo energético. Clínicamente, la deficiencia puede llegar a causar anemia y afectación del neurodesarrollo. Contrariamente, el exceso de Fe provoca complicaciones como son los desórdenes endocrinos, cirrosis hepática y disfunción cardiaca.3-6
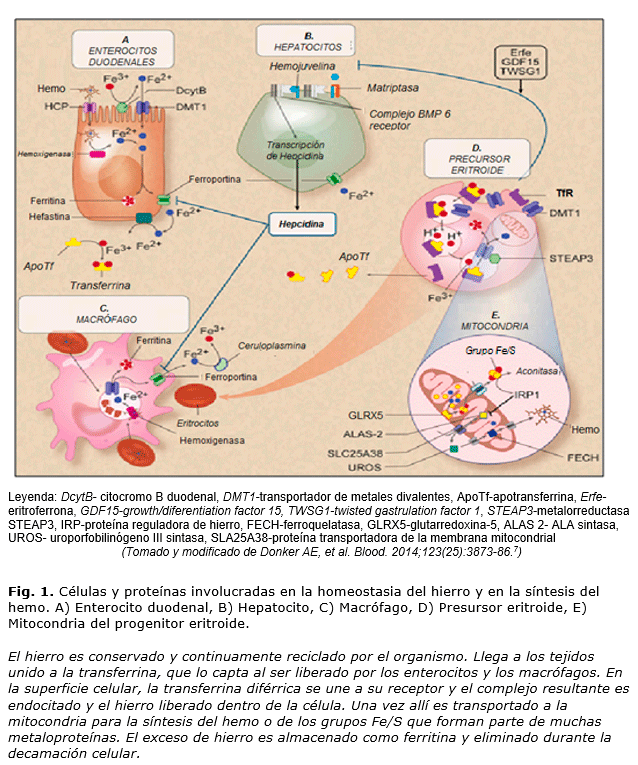
Esta es la razón de la estricta regulación de la homeostasia del hierro corporal en la cual están implicadas un número importantes de proteínas, muchas de las cuales han sido descritas en las últimas décadas. Las alteraciones en estas proteínas conducen a desórdenes del metabolismo del Fe que se caracterizan por la sobrecarga, la deficiencia o la mala distribución del mineral.2 Las células involucradas en la homeostasia del Fe son los enterocitos duodenales, los hepatocitos, los macrófagos y los precursores eritroides (figura 1).7
- Enterocito. El Fe entra al organismo a través de la dieta como Fe3+ y es absorbido fundamentalmente en el duodeno y yeyuno proximal. Este proceso ocurre en cuatro fases: en la luminal el Fe es solubilizado y convertido a Fe2+ por el citocromo B duodenal ( DcytB). Durante la fase mucosa, el Fe hemo es liberado por digestión enzimática de la hemoglobina y la mioglobina y entra al enterocito a través de la proteína transportadora de hemo (HCP); el Fe inorgánico se une al borde en cepillo y transportado dentro de la célula mucosa por el transportador de metales divalentes 1 (DMT1). En la fase celular, el hemo es degradado por la hemoxigenasa y el Fe es liberado. Una vez dentro de la célula, es almacenado en forma de ferritina celular, o transportado directamente a la cara opuesta de la célula mucosa. En la última fase, el Fe2+ es liberado a la circulación portal a través de la ferroportina (Fpn ), exportador celular basolateral, paso en que se requiere de la hefastina, una oxidasa multicobre homóloga a la ceruloplasmina (Cp), que oxida el Fe2+ a Fe3+ para ser unido y transportado por la apotransferrina. Este eflujo celular es inhibido por la unión de la hormona peptídica hepcidina a la Fpn y la subsecuente degradación del complejo Fpn-hepcidina.7-9
- El Hepatocito sirve como el principal sitio de almacenamiento de Fe (gran parte del Fe está presente en los enterocitos y macrófagos). Además, es el principal productor de hepcidina, por lo que está fuertemente implicado en la regulación sistémica del mineral. La señal de transducción parte de la membrana al núcleo e implica a una serie de proteínas que son elementos claves en la regulación de la hepcidina, como la proteína morfogénica del hueso (BMP) y su receptor, la proteína de membrana hemojuvelina (HJV), la proteína de la hemocromatosis (HFE), los receptores de transferrina (TfR)1 y 2 y la matriptasa 2 (TMPRSS6). A través de mecanismos intracelulares se da la señal de la transcripción de la hepcidina. En la deficiencia de Fe la proteasa de membrana, matriptasa-2, bloquea la transcripción al escindir a la HJV.2,5,7,10
- El macrófago pertenece al grupo de células retículoendoteliales que destruyen los glóbulos rojos senescentes. Durante este proceso, el Fe es liberado del hemo y puede ser almacenado intracelularmente como ferritina o hemosiderina, o ser entregado a los progenitores eritroides para la formación de nuevos eritrocitos. La Fpn es responsable del eflujo del Fe2+ a la circulación, para lo cual requiere la acción de la Cp, que lo oxida a Fe3+, para que sea unido a la apotransferrina, al igual que ocurre en los hepatocitos. 7,9,11
- En los progenitores eritroides la transferrina diférrica (Tf-Fe2, o sea cargada con dos átomos de Fe) es endocitada víaTfR 1, en el endosoma el Fe es liberado de la transferrina (Tf), convertido de Fe3+ a Fe2+ por la ferroreductasa STEAP3 y transportado al citosol por el DMT1, donde queda disponible, fundamentalmente para la síntesis del hemo. Se ha reportado que la eritropoyesis se comunica con el hepatocito a través de las proteínas TWSG1, GDF15 y eritroferrona (Erfe), que inhiben la señal de la hepcidina.2,7,12,13
En la mitocondria de los progenitores eritroides tiene lugar la síntesis del hemo y de los grupos Fe/S. Primero se sintetiza el ácido 5-amino levulínico (ALA), a partir de glicina y succinil coenzima A, por la enzima ALA sintasa 2 (ALAS2), en la matriz mitocondrial. La proteína SLC25A38, de la membrana mitocondrial, es probablemente la encargada de importar glicina a la mitocondria y también de exportar el ALA al citosol, donde continúa la síntesis del hemo. La uroporfobilinógeno III sintasa (UROS) citosólica, cuarta enzima de la ruta, es responsable de la conversión de hidroximetilbilano (HMB) a uroporfobilinógeno III, uno de los precursores del hemo. En el último paso, la ferroquelatasa (FECH), localizada en el espacio intermembrana mitocondrial, es la responsable de la incorporación del Fe2+ a la protoporfirina IX para formar el hemo. La expresión de la UROS y la ALAS2 en los eritroblastos es regulada por la proteína GATA1, que es crítica para la eritropoyesis normal, la expresión de los genes de la globina y el desarrollo megacariocítico. La enzima glutarredoxin-5 (GLRX5) tiene una función en la síntesis de los grupos Fe/S que son transportados al citoplasma. La alteración de cualquiera de las enzimas de la ruta de síntesis del hemo provoca enfermedades como las porfirias, en las que se produce la acumulación de intermediarios que resultan tóxicos y provocan afectaciones a diferentes órganos y sistemas.4,7,12
Este sistema homeostático mantiene la saturación de la transferrina a los niveles fisiológicos en respuesta a las señales provenientes de las vías que consumen Fe, como la eritropoyesis, y envía señales a las células que liberan Fe al torrente sanguíneo. El hierro es liberado a la circulación a partir de los enterocitos duodenales y de los macrófagos. Los hepatocitos tienen una doble función en el metabolismo sistémico del hierro, son el principal sitio de reserva y secretan la hormona reguladora hepcidina, que coordina los flujos sistémicos del mineral y controla los niveles de hierro por unión con la ferroportina. Las alteraciones congénitas o adquiridas que alteran la producción de hepcidina causan deficiencia de hierro (altos niveles de hepcidina) y sobrecarga del mineral (deficiencia de hepcidina).4,5
ABSORCIÓN DE Fe: PUNTO CRÍTICO DEL METABOLISMO
El Fe corporal total de un adulto promedio es entre 4 y 5 mg. Para mantener la homeostasia sistémica debe existir un estricto balance entre el Fe incorporado y el que se pierde. Hasta el presente no se ha demostrado la existencia de ningún mecanismo regulatorio de la excreción por lo que la absorción es el proceso críticamente controlado. Normalmente solo son absorbidos de 1 a 2 mg de Fe de la dieta para compensar las pérdidas diarias (0,5 - 1 mg) del mineral, debido fundamentalmente a la decamación de la piel y las mucosas.1-3
La fracción de Fe absorbida es generalmente baja en comparación con la cantidad ingerida, puede oscilar entre el 5 % y el 35 %, en dependencia de las circunstancias y el tipo de hierro. La absorción puede aumentar cuando hay incremento en las necesidades, como ocurre en el embarazo o con el aumento de la eritropoyesis; y disminuir cuando hay sobrecarga del mineral. Existe un mecanismo de retroalimentación que favorece el aumento de la absorción en personas con deficiencia de Fe; sin embargo, en personas con sobrecarga la absorción disminuye por acción de la hepcidina. 1-3,14
El estado físico del Fe es una determinante de su absorción. Al pH fisiológico, el Fe2+ es rápidamente oxidado a Fe 3+, que es insoluble y precipita, por lo que no puede ser captado por la célula mucosa. El ácido gástrico contribuye a disminuir el pH en el duodeno proximal con lo que favorece la solubilización y captación del Fe. Cuando la producción de ácido gástrico está afectada por ingestión de medicamentos como los bloqueadores H2, de la bomba ácida o por cirugías, la absorción se ve sustancialmente disminuida.1,3,14
Existe una serie de factores dietéticos que influyen en la absorción de Fe. El ascorbato y el citrato favorecen la absorción debido a que solubilizan el mineral en el duodeno mediante la formación de quelatos débiles, de esta forma el Fe es fácilmente transferido al epitelio absortivo. Por su parte, los fitatos, el salvado, los polifenoles y los taninos inhiben la absorción de Fe pues forman quelatos insolubles que impiden su captación por la célula mucosa. La absorción del Fe hemo es un proceso más eficiente e independiente del pH duodenal y es poco afectado por los componentes de la dieta. Consecuentemente, las carnes son consideradas excelentes fuentes de hierro.3
Biodisponibilidad
El Fe se presenta en la dieta como Fe hemo u orgánico y como Fe no hemo o inorgánico. Las principales fuentes de hemo son la hemoglobina y la mioglobina provenientes del consumo de proteína animal (carnes, aves y pescados). Este tipo de hierro es muy absorbible (15 - 35 %) y poco afectado por la composición dietética. Por su parte, el Fe inorgánico es obtenido a partir de los cereales, las legumbres, los frutos y los vegetales; su absorción es mucho menor (2 - 20 %) y está muy influenciado por la composición de la dieta, pero a diferencia del Fe hemo, su presencia y por tanto su contribución a la nutrición, es mucho mayor a pesar de su pobre biodisponibilidad.1,14
Los elementos favorecedores incluyen al ácido ascórbico y al tejido muscular que pueden reducir el Fe3+ y unirlo en complejos solubles que facilitan la absorción. El efecto favorecedor del ácido ascórbico es dependiente de la dosis y puede revertir el efecto negativo de todos los inhibidores y facilitar la absorción del Fe natural y adicionado. La cocción, el procesamiento industrial y el almacenamiento destruyen el ácido ascórbico y por tanto, eliminan su efecto positivo. Las carnes, aves y pescados además del Fe hemo, aportan ácido ascórbico. Se plantea que 30 g de tejido muscular equivalen a 25 mg de este agente reductor y que su presencia en la dieta aumenta la absorción de Fe inorgánico entre 2 y 3 veces.1,14
Los principales inhibidores de la absorción de Fe son el ácido fítico, los polifenoles, el calcio y los péptidos formados por la digestión parcial de las proteínas. El efecto inhibidor del fitato depende de la dosis y comienza a concentraciones muy bajas (2 - 10 mg/comida). Los polifenoles son ampliamente consumidos pues se encuentran en los vegetales, las frutas, los cereales, las legumbres, así como en el té, el café y el vino.
El efecto negativo del calcio afecta, tanto la absorción del Fe hemo como del no hemo; también depende de la dosis. Las proteínas animales de alto peso molecular, como la de la leche, el huevo y la albúmina, también disminuyen la absorción de Fe. También se ha planteado que algunos metales pesados como el plomo, el manganeso, el cobalto y el zinc, pueden competir con el Fe por su vía de absorción, con la correspondiente afectación del proceso.1,14
Un aparte merece el consumo de leche de vaca, que puede contribuir al desarrollo de un estado deficitario por varios mecanismos. Primeramente, el contenido de Fe en la leche es bajo con una biodisponibilidad menor que la de la leche materna. Además, su consumo tiende a remplazar a los alimentos ricos en Fe. Adicionalmente, en la composición de la leche de vaca se encuentra calcio y caseína, que pueden interferir directamente con la absorción de Fe. La leche entera también contiene proteínas que pueden irritar el epitelio del tracto gastrointestinal del lactante y, aunque en bajo grado pero de forma crónica, pueden provocar hemorragias que predisponen a la deficiencia de Fe. El crecimiento neonatal requiere de una gran cantidad de Fe que se contrapone a las desventajas del consumo de leche de vaca. Por estas razones, las recomendaciones internacionales plantean excluir la leche de vaca de la dieta en el primer año de vida y limitar su consumo posterior a no más de 24 onzas diarias. En el caso de los niños que no pueden ser lactados se recomienda el empleo de fórmulas fortificadas y de suplementación con el mineral.3
TRANSPORTE INTERCELULAR DE Fe
Puesto que solo una pequeña fracción del total de Fe corporal entra y abandona el organismo diariamente, el transporte intercelular resulta cuantitativamente más importante, especialmente si se tiene en cuenta que la fracción de Fe que circula en el pool intercambiable es solo el 0,1 % o 4 mg del Fe total, y lo hace unido a la Tf, que es el principal suministrador fisiológico del mineral. De hecho, la Tf plasmática se encarga de la entrega de Fe a casi todos los tejidos. En estas circunstancias, la Tf tiene tres propósitos: 1) mantener el Fe en estado soluble en las condiciones fisiológicas; 2) prevenir la toxicidad por radicales libres mediada por Fe y 3) facilitar el transporte a las células. Además, se ha planteado que la concentración de Tf – Fe2 modula la expresión de la hepcidina. Es por ello que su producción está regulada a múltiples niveles por factores nucleares y estímulos, como los niveles de Fe, las hormonas y la inflamación. Así, por ejemplo, en la deficiencia de hierro los niveles de Tf sérica aumentan sustancialmente como resultado de la estimulación de la síntesis de su ARN mensajero (ARNm) por el hígado.2,3,8
HOMEOSTASIA CELULAR: EL SISTEMA IRE/IRP
El mantenimiento de la homeostasia del Fe por las células implica acciones similares a las del nivel sistémico, como coordinar la incorporación, utilización y almacenamiento del metal para asegurar la disponibilidad de un suministro adecuado y prevenir la toxicidad. Sin embargo, la maquinaria y los mecanismos son enteramente diferentes pues, en contraste con el metabolismo sistémico, el tráfico celular involucra la excreción regulada del mineral.2,4
La Tf-Fe2 es la principal fuente de Fe para las células, que lo adquieren vía TfR1 de elevada afinidad. El complejo Tf-Fe 2/TfR es internalizado por endocitosis dependiente de clatrina. La acidificación del endosoma provoca cambios conformacionales en la Tf y en su receptor que permiten la liberación del Fe, que una vez liberado es reducido a Fe2+ para su paso al citosol. La apotransferrina y el TfR1 son reciclados a la superficie celular.3,4
Aunque el ciclo de la Tf es la principal vía de adquisición de Fe por las células, existen evidencias de que algunas células pueden adquirir Fe independientemente de este ciclo. Este hecho es soportado por estudios bioquímicos y genéticos que evidencian la existencia de rutas de incorporación de Fe independientes de la Tf, como el DMT1 en los hepatocitos, canales de calcio en los cardiomiocitos y células neuronales, así como endocitosis mediada por receptores de otras formas de Fe unido a proteínas como la lipocalina 2; es decir, las células especializadas son capaces de adquirir Fe en forma de hemo.4
La exportación de Fe ocurre en muchas células, incluidas las neuronales y las eritroides, pero es particularmente importante en las que mantienen los niveles de Fe plasmático, los macrófagos y los enterecitos duodenales, y en desarrollo fetal es mediado por las células del endodermo visceral extraembrionario y por los sincitiotrofoblastos placentarios. Estas células expresan niveles relativamente altos de Fpn, que como se mencionó, transporta el Fe2+ en coordinación con una ferroxidasa, la hefastina en los enterocitos y la Cp en los otros tipos celulares, que facilitan la extracción del Fe del canal de la Fpn y su entrega a la Tf plasmática. Adicionalmente, las células parecen ser capaces de exportar Fe hemo, pero la función fisiológica de este mecanismo, así como el de la salida de Fe unido a ferritina, no están bien establecidas.4
El Fe que queda sin utilizar en el citoplasma es almacenado en forma de ferritina. La ferritina provee a la célula con un mecanismo de atrapamiento del exceso de Fe en una forma no reactiva para prevenir el daño celular y tisular mediado por Fe; además, constituye la reserva que puede movilizarse por degradación proteosomal y lisosomal de la proteína.3,4.
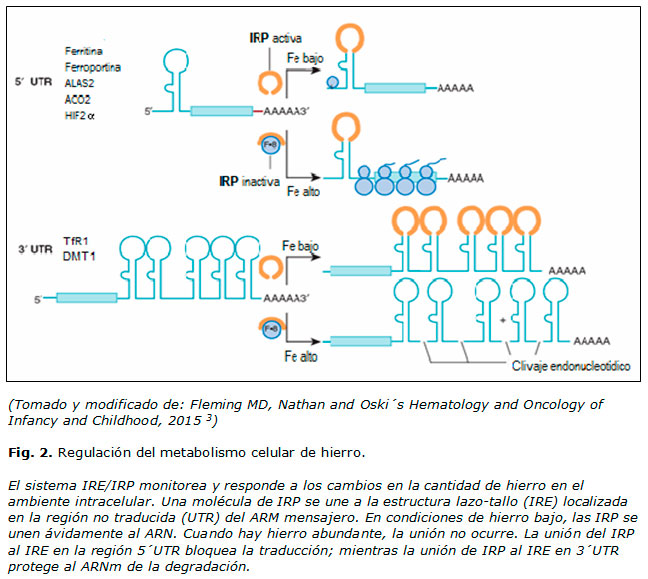
Regulación del metabolismo celular del Fe
Mientras que los aspectos claves del metabolismo sistémico son regulados a nivel transcripcional (expresión de la hepcidina) y postraduccional (Fpn por acción de la hepcidina), la homeostasia celular es coordinada postranscripcionalmente por las proteínas reguladoras de Fe, IRP 1 y 2 (figura 2), que interactúan con regiones conservadas de los ARNm, de estructura lazo-tallo, conocidas como IRE (elemento de respuesta al Fe), que están presentes en las regiones 5´ o 3´ no traducidas (UTR) de los ARNm. Cualquiera de las IRP inhibe la iniciación de la traducción cuando se une al IRE simple en la región 5´ UTR de los ARNm de la ferritina, Fpn, aconitasa mitocondrial o factor inducible de hipoxia 2a (HIF2α), mientras que la unión a los múltiples IRE dentro de la región 3´ UTR del ARNm de TfR1 previene su clivaje endonucleolítico y subsecuente degradación. Las IRP también parecen regular positivamente la expresión del ARNm del DMT1 a través de un IRE simple en 3´ UTR. 2-4,8,12
La unión de IRP al IRE responde a los niveles de Fe celular. En células deficientes de Fe, IRP 1 o 2 se unen a losIRE presentes en las UTR de los ARNm que codifican proteínas involucradas en el transporte y almacenamiento. La unión a los IRE simples de la región 5´ UTR inhibe la traducción, mientras que la unión a los múltiples IRE en el transcripto del TfR aumenta su estabilidad. Como consecuencia, aumenta la captación de Fe mediada por el TfR1, mientras que el almacenamiento en la ferritina y la exportación vía Fpn disminuyen. En las células repletas de Fe se produce la ubiquitinación y degradación proteosómica de las IRP. Las IRP también modulan la traducción de los ARNm que codifican la enzimas especifica de la síntesis del hemo ALAS2, la aconitasa mitocondrial y el HIF2α. 2-4,8,12,15
INTERACCIÓN ENTRE REGULACIÓN CELULAR Y SISTÉMICA
La homeostasia celular y sistémica son mantenidas por sistemas de control independientes, IRE/IRP y hepcidina/Fpn, pero entre los que hay una estrecha coordinación, al menos se han identificado tres interconexiones.4,12
1. La conexión de Fpn: la expresión de esta proteína, critica para los niveles de Fe plasmáticos, está sujeta a la regulación por ambos sistemas. Así, el estado de Fe sistémico es comunicado postraduccionalmente vía hepcidina, mientras que la disponibilidad de Fe celular regula la síntesis de Fpn vía IRE en 5´UTR del ARNm de la Fpn. El sistema IRE/IRP protege a las células de exportadoras de Fe contra las pérdidas perjudiciales, mientras que la hepcidina protege al organismo contra la sobrecarga sistémica.
2. La conexión HIF2α: el ARNm del HIF2α es una diana de la IRP y el factor de transcripción codificado regula la expresión del DMT1 en la superficie apical de los enterocitos y la transcripción de hepcidina en respuesta a la hipoxia o la deficiencia de hierro.
3. La conexión TfR: la expresión de hepcidina es regulada por la señal de la TfR 2 y la HFE que también se une al TfR 1 en competencia con la Tf-Fe2. La expresión de TfR1 es promovida por la alta actividad de las IRP. Se piensa que el equilibrio entre la cantidad de Fe plasmático “sentido” por el TfR 1 y “señalizado” por TfR 2 es importante para la activación de la hepcidina, y la actividad IRP puede indirectamente afectar la expresión de la hepcidina por regular los niveles de TfR 1 en los hepatocitos.2-4,8,12
El metabolismo del hierro está balanceado por dos sistemas regulatorios, uno sistémico basado en la hormona hepcidina y la proteína exportadora Fpn; y el otro, que controla el metabolismo celular a través de las IRP que se unen a los IRE de los ARNm regulados. Estos sistemas funcionan de modo coordinado como una pareja de bailarines que ejecutan sus rutinas con extrema precisión.
Los avances de las últimas décadas han permitido mejorar sustancialmente el entendimiento del metabolismo de este mineral y ya se vislumbran aplicaciones diagnósticas y terapéuticas de los nuevos conocimientos, pero aún quedan múltiples aspectos sobre los que investigar, ese es el reto que impone este metal, no tan brillante como el oro y la plata pero sin dudas muy valioso para vida.
REFERENCIAS BIBLIOGRAFICAS
1. Abbaspour N, Hurrell R, Kelishadi R. Review on iron and its importance for human health. J Res Med Sci. 2014 Feb;19(2):164–74.
2. Finberg KE. Unraveling mechanisms regulating systematic iron homeostasis. Hematology Am Soc Hematol Educ Program. 2011;2011: 532–537. doi:10.1182/asheducation-2011.1.532.
3. Fleming MD. Disorders of Iron and Copper Metabolism, the Sideroblastic Anemias, and Lead Toxicity. In: Orkin SH, Fisher DE, Ginsburg D, Look T, Lux SE, Nathan DG. Nathan and Oski´s Hematology and Oncology of Infancy and Childhood. 8th ed. Philadelphia: Elsevier Saunders; 2015.
4. Hentze MW, Muckenthaler MU, Galy B, Camaschella C. Two to Tango: Regulation of Mammalian Iron Metabolism. Cell. 2010 Jul;142(1):24-38.
5. Camaschela C. Iron Deficiency Anemia. New Engl J Med. 2015 May;372(19):1832-43.
6. Fleming RE, Pomka R. Iron overload in human disease. New Engl J Med. 2012;366(4):348-59.
7. Donker AE, Raymakers RAP, Vlasveld LT, van Barneveld T, Terink, R, Dors N, et al. Practice guidelines for the diagnosis and management of microcytic anemias due to genetic disorders of iron metabolism or heme synthesis. Blood. 2014;123(25):3873-86. DOI 10.1182/blood-2014-01-548776.
8. Wang J, Pantopoulos K. Regulation of cellular iron metabolism. Biochem J. 2011;434:365–81.
9. Theil EC, Chen H, Miranda C, Janser H, Elsenhans B, Núñez MT, et al. Absorption of iron from ferritin is independent of heme iron and ferrous salts in women and rat intestinal segments. J Nutr. 2012;142:478–83.
10. Meynard D, Babitt JL, Lin HY. The liver: conductor of systemic iron balance. Blood. 2014 Ene; 123(2): 168-75. DOI 10.1182/blood-2013-06-427757.
11. Camaschella C. Iron and hepcidin: a story of recycling and balance. Hematology Am Soc Hematol Educ Program. 2013;2013:1-8. doi: 10.1182/asheducation-2013.1.1.
12. Ganz T, Nemeth E. Iron Metabolism: Interactions with Normal and Disordered Erythropoiesis. Cold Spring Harb Perspect Med. 2012;2:a011668.
13. Kautz L, Jung G, Valore EV, Rivella S, Nemeth E, Ganz T. Identification of erythroferrone as an erythroid regulator of iron metabolism. Nat Genet. 2014 July;46(7):678–684. doi:10.1038/ng.2996.
14. Hurrell R, Egli I. Iron bioavailability and dietary reference values. Am J Clin Nutr. 2010;91:1461–7S.
15. Mastrogiannaki M, Matak P, Peyssonnaux C. The gut in iron homeostasis: role of HIF-2 under normal and pathological conditions. Blood. 2013 Aug;122(6):885-91. doi:10.1182/blood-2012-11-427765.
Recibido: agosto 03, 2015.
Aceptado: agosto 17, 2015.
Mariela Forrellat Barrios . Instituto de Hematología e Inmunología. Apartado 8070, La Habana, CP 10800, CUBA. Tel (537) 643 8695, 8268. Email: rchematologia@infomed.sld.cu


{kind=link}
{kind=link}