










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Investigaciones Biomédicas
versión impresa ISSN 0864-0300
Rev Cubana Invest Bioméd vol.32 no.2 Ciudad de la Habana abr.-jun. 2013
ARTÍCULO ORIGINAL
Diferencias del estado redox entre espermatozoides y plasma seminal en adultos jóvenes
Difference of redux status between spermatozoa and seminal plasma in young adults
MSc. Yanet Hernández-Matos,I MSc. Liván Delgado Roche,I Gregorio Martínez-SánchezII
ICentro de Estudios para las Investigaciones y Evaluaciones Biológicas, Instituto de Farmacia y Alimentos, Universidad de la Habana, La Habana, Cuba.
IIMediNat Srl. Ancona, Italia.
RESUMEN
Introducción: Las especies reactivas del oxígeno y el estrés oxidativo desempeñan un papel fundamental en la infertilidad masculina. Sin embargo, estas especies oxidantes también han sido asociadas con los procesos de capacitación de los gametos masculinos cuando son generadas a bajos niveles y de manera controlada. Actualmente los biomarcadores redox son introducidos en el diagnóstico clínico de la infertilidad masculina en el mundo, como una herramienta complementaria a los parámetros del espermiograma. Sin embargo, en Cuba, esta metodología aún no se encuentra extendida a los servicios de salud.
Objetivo: El objetivo del presente trabajo fue caracterizar el estado redox de espermatozoides y el líquido seminal de sujetos aparentemente sanos, a través de la determinación de una serie de biomarcadores de estrés oxidativo. Los niveles de malonildialdehído, la actividad de las enzimas antioxidantes superóxido dismutasa y catalasa, así como los niveles de glutatión reducido, el potencial de peroxidación y la capacidad reductora fueron determinados mediante técnicas espectrofotométricas.
Métodos: Se analizaron 40 muestras de semen de sujetos aparentemente sanos, las cuales fueron obtenidas mediante masturbación sin el empleo de lubricantes y con al menos tres días de abstinencia eyaculatoria. En el estudio se incluyeron sujetos de 20 a 35 años, aparentemente sanos según exámenes de laboratorio clínico y con paternidad probada.
Resultados: Los resultados demostraron que existen diferencias significativas (p< 0,05) entre los marcadores evaluados en el líquido seminal con respecto al espermatozoide, lo cual sugiere la existencia de un estado redox diferenciado entre ambos compartimentos.
Conclusiones: Estos hallazgos demuestran la necesidad de evaluar el nivel de estrés oxidativo tanto en las células sexuales como en el fluido que las contiene. Ello contribuirá, sin lugar a dudas, a un diagnóstico más eficaz e integral de la capacidad fértil del hombre.
Palabras clave: especies reactivas del oxígeno, estrés oxidativo, espermatozoide, líquido seminal, estado redox diferenciado.
ABSTRACT
Background: Reactive oxygen species and oxidative stress play a fundamental role in male infertility. However, these oxidizing species have also been associated with the process of capability of male gametes when they are generated at low levels and in a controlled manner. Currently, redox biomarkers are introduced in the clinical diagnosis of male infertility in the world as a complementary tool to the spermiogram parameters. However, in Cuba, this methodology is not yet extended to health services.
Objective: The objective of this study was to characterize the redox status of spermatozoa and seminal fluid of apparently healthy subjects through the identification of a number of biomarkers of oxidative stress.
Methods: 40 samples of semen of apparently healthy subjects were analyzed, which were obtained by masturbation without the use of lubricants and with at least 3 days of ejaculatory abstinence. The study included subjects from 20 to 35 years of age, who were apparently healthy according to both laboratory tests and paternity test results.
Results: The results showed that there are significant differences (p<0, 05) between the markers evaluated in the seminal fluid and the spermatozoon which suggests the existence of a differentiated redux status between the two compartments.
Conclusions: These findings demonstrate the need to evaluate the level of oxidative stress in both sexual cells and the fluid that contains them. It will contribute, with no doubt, to a more effective and comprehensive diagnosis of man's fertility.
Key words: oxygen-reactive species, oxidative stress, spermatozoon, seminal fluid, differenciated redux status.
INTRODUCCIÓN
La infertilidad masculina se ha incrementado notablemente a nivel mundial en los últimos años. Diversas son las causas que la condicionan, algunas de origen genético y otras asociadas a la exposición a agentes nocivos. A nivel global, 1 de cada 20 hombres es infértil.1
La adquisición de la capacidad fértil del espermatozoide eyaculado es un proceso complejo que aún no está del todo esclarecido y que se conoce como capacitación.2 Las especies reactivas del oxígeno (ERO) juegan un papel decisivo en este proceso. Se ha podido comprobar que la producción, bajo condiciones fisiológicas y controladas de estas, es de vital importancia para que se produzca la capacitación del espermatozoide.3 Además, intervienen otros mecanismos que están regulados por ERO como es el influjo de calcio, el aumento de monofosfato cíclico de adenosina (AMPc), la fosforilación de proteínas, entre otros.4
Sin embargo, una sobreproducción de ERO incide negativamente sobre la fertilidad masculina. Estas moléculas reactivas provocan daños a la membrana espermática, lo cual da lugar a una reducción de la motilidad y habilidad del espermatozoide de fusionarse con el óvulo. Por otra parte, dañan directamente el ADN espermático, a través del cual afectan la genómica paternal del embrión.1
La Organización Mundial de la Salud (OMS), ha establecido una serie de parámetros del espermiograma para evaluar la fertilidad en el hombre.5 Sin embargo, consideramos que la determinación clínica de una serie de marcadores del estado redox, constituye una herramienta de gran utilidad que puede contribuir a la interpretación de los procesos fisiológicos y fisiopatológicos que están asociados a la capacidad fértil del hombre. Por esta razón, en el presente estudio se caracterizó el estado redox del líquido seminal y espermatozoides de sujetos sanos, con el objetivo de determinar si existen diferencias en cuanto al nivel de estrés oxidativo (EO) entre ambos compartimentos.
MÉTODOS
Aspectos éticos de la investigación
El protocolo del estudio clínico fue revisado y aprobado por los Comités de ética e investigación en humanos de ambas partes participantes en el estudio, el Instituto de Farmacia y Alimentos de la Universidad de La Habana y el Hospital "Hermanos Ameijeiras". Todos los procedimientos llevados a cabo estuvieron en correspondencia con los principios promulgados por la Declaración de Helsinki (1997). Todos los pacientes incluidos en el estudio ofrecieron su consentimiento informado de forma oral y escrita, luego de recibir toda la información concerniente al estudio, así como todas las obligaciones y derechos una vez incluidos en el mismo.
Muestra poblacional
En el estudio se incluyeron sujetos de 20 a 35 años, aparentemente sanos según exámenes de laboratorio clínico y con paternidad probada. Se tuvo en cuenta, además, que los parámetros del espermiograma (motilidad, viabilidad, infiltrado de leucocitos, pH, concentración espermática) estuvieran en correspondencia con los valores establecidos como normales. Se excluyeron aquellos hombres que presentaran antecedentes de alguna enfermedad aguda o crónica (urogenital, hepática, metabólico-endocrina, cardiovascular, neurológica, respiratoria, del sistema inmunológico, entre otras); antecedentes de alcoholismo, tabaquismo u otras adicciones tóxicas. También fueron excluidos los que estuviesen bajo tratamiento farmacológico o consumo de algún suplemento nutricional en fase de investigación.
Obtención de las muestras biológicas
Se analizaron 40 muestras de semen de sujetos aparentemente sanos, las cuales fueron obtenidas mediante masturbación sin el empleo de lubricantes y con al menos tres días de abstinencia eyaculatoria. Las muestras fueron centrifugadas a 800 g × 10 min a 4 ºC. El sobrenadante (líquido seminal) fue separado del sedimento (espermatozoides). Este último fue lavado en una solución de cloruro de sodio 0,9 % y centrifugado nuevamente. Una vez culminado este segundo proceso de centrifugación se separó el sobrenadante y el sedimento (espermatozoides) fue resuspendido en agua desionizada para lisar las células por shock osmótico6. Tanto el líquido seminal como el lisado de espermatozoides fueron almacenados a -80 ºC hasta el momento de las determinaciones bioquímicas.
espermiograma
Las variables analizadas fueron la movilidad, viabilidad, concentración espermática, pH e infiltrado de leucocitos, además de los parámetros físicos.
Movilidad espermática
La movilidad espermática fue determinada según la clasificación sugerida por la OMS.7
Viabilidad espermática
Con el propósito de determinar la viabilidad espermática se utilizó el método de exclusión de la eosina.8 Se añadieron 50 µL del colorante a una dilución 1:2 de semen en solución amortiguadora fosfato. Tras agitarse suavemente por unos segundos se tomaron 50 µL de la muestra y se extendieron sobre un portaobjetos. Tras la incubación a 37 ºC durante 30 min, se fijó la preparación y se cuantificaron los espermatozoides que excluían la eosina. A estos se les consideró vivos en el momento de la fijación, ya que los espermatozoides muertos exhiben una coloración rosada al no excluir a la eosina. Se contabilizaron al microscopio óptico con una magnificación de 100X.
Determinación de leucocitos
La concentración de los leucocitos se determinó a través del procedimiento para la cuantificación de células peroxidasa positivas.9 Se utilizó una solución de benzidina al 0,012 % y H2O2 al 0,038 % diluidos en etanol al 50 %. A 20 µL de esta solución se le añadieron 20 µL de semen, se colocaron en un hemocitómetro y se contaron las células que adquirieron una tonalidad parda (células peroxidasa positivas).
Concentración espermática y pH del semen
El análisis de estos parámetros fue realizado según los procedimientos descritos en el manual publicado por la OMS, 1999.7
Determinaciones de biomarcadores redox
Todos los biomarcadores redox fueron determinados a través de técnicas espectrofotométricas con la utilización de un espectrofotómetro Pharmacia 1 000 (Pharmacia LKB, Uppsala, Suiza) y un lector de placas de ELISA (SUMA, Centro de Inmunoensayos, La Habana, Cuba).
Proteínas totales
Las proteínas del medio reaccionan con el reactivo Azul de Coommasie cuyo producto es detectable espectrofotométricamente y tiene un máximo de absorción a una longitud de onda de 595 nm. Esta técnica es válida para la detección de las proteínas que se encuentren en un intervalo de 0,01-0,05 mg.10
Actividad antioxidante enzimática
La actividad de la enzima superóxido dismutasa (SOD) fue evaluada mediante la utilización de un kit comercial Cat. No. SD125 (Diamond Road, Crumlin, Reino Unido). El método emplea xantina y xantina oxidasa para generar radical O2•-, el cual reacciona con el reactivo 2-(4-iodofenil)-3-(4-nitrofenol)-5-feniltetrazolio cloruro (INT) y forma un complejo coloreado rojo detectable a 420 nm. La actividad enzimática fue medida a través de la inhibición de esta reacción.11
La actividad de la enzima catalasa fue determinada a través del seguimiento de la descomposición del H2O2 a 240 nm en 1 min.11
Indicadores de daño biomoléculas
Para cuantificar la concentración de hidroperóxidos totales (ROOH) se utilizó el kit comercial Bioxytech H2O2-560 (Oxis International Inc., Portland, OR, EU). El ensayo se basa en la oxidación de Fe2+ a Fe3+ por los ROOH bajo condiciones de acidez. Los iones Fe3+ se unen al indicador xilenol naranja (3,3-bis(N,N-di(carboximetil)-aminometil)-o-cresolsulfona-ftalein, sal sódica) (Sigma St. Louis, MO, EU), para formar un complejo coloreado estable que puede ser detectado a una longitud de onda de 560 nm.
La concentración de malonildialdehído (MDA) fue medida mediante la utilización del kit comercial LPO-586 obtenido de la firma Calbiochem (La Jolla, CA, EU). El ensayo se basa en la formación de un cromóforo estable luego de la incubación de la muestra con el reactivo durante 40 min a 45 °C detectable a 586 nm. Para la generación de la curva de calibración se utilizó un patrón de referencia de malonildialdehído bis [dimetil acetal] (Sigma St Louis, MO, EU) y se procesó bajo las mismas condiciones experimentales.12
Los PAOP fueron cuantificados como se describe en la literatura.13 Fueron tomados 100 µL de suero y se diluyeron en 1 mL de solución amortiguadora fosfato, posteriormente se añadieron 50 µL de yoduro de potasio 1,16 M y 100 µL de ácido acético glacial. La absorbancia de la reacción fue medida a 340 nm contra blanco reactivo. La concentración de PAOP fue expresada en µM de cloramina T (Sigma St. Louis, MO, EU), sustancia patrón que fue utilizada para generar la curva de calibración.
Antioxidantes de bajo peso molecular
La concentración de glutatión (GSH) fue determinada según se describe en la literatura,14 el cual utiliza el reactivo de Ellman (5,5`dithiobis-2-nitrobenzoic acid) 10-2 M (Sigma St. Louis, MO, EU), la absorbancia fue medida a una longitud de onda de 412 nm. Se utilizó un patrón de GSH (Sigma St Louis, MO, EU) para generar la curva de calibración.
Susceptibilidad a la peroxidación lipídica
Para evaluar la susceptibilidad a la peroxidación lipídica (POL) se calculó el potencial de peroxidación (PP). Las muestras de suero fueron incubadas con una solución de sulfato de cobre II (2 mM) a 37 °C durante 24 h. Posteriormente, se determinó la concentración de MDA a la cual se le restó la determinada sin incubación con cobre, la diferencia resultante se considera como PP.
Estado antioxidante total
La concentración de agentes reductores (medida a través de FRAP) fue determinada con el objetivo de conocer el estado antioxidante total.15 La técnica se basa en la reducción del Fe3+ por parte de los agentes reductores presentes en la muestra, para lo cual se midió la densidad óptica del complejo Fe2+-TPTZ (2, 4, 6-tripiridil-s-triazina) (Sigma St. Louis, MO, EU) el cual absorbe a una longitud de onda de 593 nm.
Procesamiento estadístico
Inicialmente se realizó un análisis exploratorio de los datos para la detección de valores aberrantes. Posteriormente se llevó a cabo el análisis de homogeneidad de varianza (Bartlett-Box). La comparación de medias entre los grupos fue realizada a través de la utilización de la prueba t de student de muestras no pareadas. Los datos experimentales fueron expresados como la media ± la desviación estándar (DE). El nivel de significación estadística fue asumido como p< 0,05. El procesamiento estadístico se realizó mediante el programa Statistic para Windows, versión 6.0.
RESULTADOS
En la tabla 1 se muestran los valores de los diferentes indicadores del espermiograma. En su mayoría se encontraron dentro del intervalo de referencia establecido por el laboratorio clínico del hospital "Hermanos Ameijeiras". Es de señalar que en la mayoría de las muestras se pudo observar que los espermatozoides mostraban una movilidad progresiva lineal rápida (A) y una movilidad progresiva lineal lenta, fundamentalmente (B). Además, la concentración espermática fue superior al 70 % y la viabilidad superior al 60 %, así como una concentración de leucocitos en el rango establecido como normal. El espermiograma demuestra que los individuos incluidos en el estudio poseían capacidad fértil, lo cual fue corroborado con la paternidad probada de cada uno de ellos.
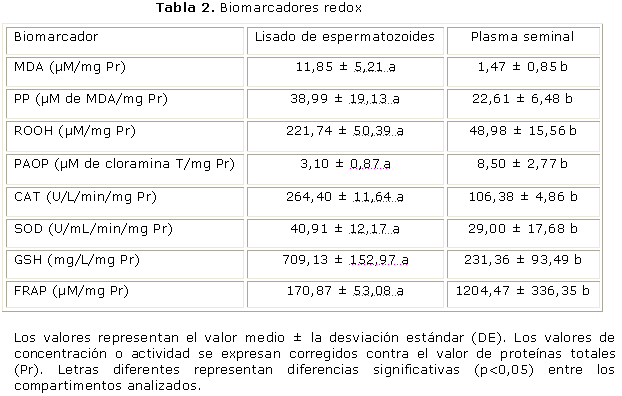
En la tabla 2 se muestra el comportamiento de los biomarcadores redox. En la población estudiada, las concentraciones de MDA y ROOH son significativamente mayores (p< 0,05) en el lisado de espermatozoides que en el plasma seminal. En tanto, los niveles de PAOP son significativamente (p< 0,05) mayores en el plasma seminal que en lisado. En cuanto a la actividad de las enzimas antioxidantes se observó un incremento significativo (p< 0,05) de ambas en el lisado de espermatozoides en comparación con el plasma seminal. Por otra parte, los niveles de GSH fueron significativamente mayores (p< 0,05) en el lisado de espermatozoides que en el plasma seminal, mientras que la susceptibilidad a la POL fue significativamente (p< 0,05) mayor en espermatozoides y la capacidad antioxidante total mostró un incremento significativo (p< 0,05) en plasma seminal con respecto al lisado de espermatozoides.
DISCUSIÓN
La espermatogénesis es un proceso multifactorial que se produce en los testículos durante la vida sexual activa, comienza alrededor de los 13 años como consecuencia de la estimulación de las hormonas gonadotrópicas de la adenohipófisis y continúa por el resto de la vida.1 La capacidad fértil se relaciona estrechamente al proceso espermatogénico y el inicio del mismo no es exactamente igual en todos los hombres. Se conoce que aproximadamente a partir de los 17 años, la capacidad de fecundar es elevada a no ser que exista alguna anomalía. El promedio de edades de los sujetos incluidos en el estudio fue de 30,5 años, aspecto que se corresponde con lo descrito en la literatura como una de las edades de mayor actividad reproductora.16 Este fue un elemento fundamental a la hora de incluir los individuos en este estudio clínico, además de tener en cuenta de que tuvieran al menos un hijo como criterio de paternidad probada.
El proceso de fertilización en mamíferos involucra la interacción directa entre el espermatozoide y el óvulo, la fusión entre sus membranas y la unión del genoma contenido en los gametos masculinos y femeninos.17 El éxito de este fenómeno está estrechamente relacionado con varios parámetros que durante más de 25 años la OMS ha estandarizado para estudiar el potencial de fertilidad del semen. Estas técnicas se basan en la observación y medición de parámetros tales como las propiedades físicas del semen eyaculado (color, viscosidad, volumen), la cuantificación del contenido celular (tanto de espermatozoides como de leucocitos), la viabilidad espermática, así como la morfología y la motilidad de las células sexuales masculinas.18
En el presente estudio, la mayoría de los indicadores del espermiograma se encontraron en los intervalos de referencia establecidos; sólo en el caso de la clasificación D para la movilidad de los espermatozoides se observó un ligero aumento con respecto a los valores de referencia, pues en dos de los pacientes se observaron células sexuales con estas características, lo que pudiera estar asociado con algún proceso patológico desconocido por estos individuos. De manera general los resultados del espermiograma demuestran la capacidad fecundativa de los individuos incluidos en el estudio.
Con el devenir del tiempo y el surgimiento de nuevos conocimientos acerca de los procesos involucrados en la capacidad de fertilización de las células sexuales masculinas, han emergidos nuevos parámetros a tener en cuenta a la hora de evaluar sus potencialidades en el humano.5 La OMS ha plateado que indicadores como la fragmentación del ADN, la apoptosis de células sexuales y el EO deben ser necesariamente tomados en consideración. Todos ellos son el reflejo de la existencia de factores de riesgo o procesos fisiopatológicos que inciden notablemente sobre un problema de salud como es la infertilidad masculina. Su interpretación puede influir decisivamente sobre el diagnóstico precoz de dicha problemática.19,20
La evaluación clínica de marcadores del sistema redox constituye una metodología innovadora utilizada en los últimos años para interpretar algunos de los procesos que ocurren en el organismo, la espermatogénesis y la infertilidad masculina no quedan excluidos de esto. En el presente estudio se determinaron una serie de marcadores redox en muestras de semen de sujetos aparentemente sanos, con el objetivo de conocer el comportamiento del estado de óxido-reducción, tanto en las células sexuales masculinas como en el líquido seminal que las contienen.
Los lípidos de la membrana del espermatozoide constituyen componentes de gran importancia desde el punto de vista estructural y funcional. Los fosfolípidos (FL) de las células espermáticas son mayoritariamente ácidos grasos poliinsaturados (AGPI) de cadena larga pertenecientes a las series n-3 y n-6 cuyas proporciones varían según la especie.16 El alto grado de insaturación de los FL contribuye al movimiento celular y metabolismo lipídico, además confiere suficiente fluidez a la membrana plasmática del espermatozoide necesaria para los procesos de fusión con el óvulo.22 Sin embargo, altas concentraciones de AGPI incrementan la susceptibilidad de la célula de sufrir un daño oxidativo inducido por ERO, lo cual se considera una causa de infertilidad masculina.23 Por otra parte, en concentraciones moderadas de ERO juegan un papel importante en la regulación de varias funciones espermáticas tales como la capacitación y la reacción acrosomal.24
A pesar de que es conocida la toxicidad del MDA sobre las células, cabe destacar que durante la capacitación existe una generación fisiológica de ERO, la cual puede conducir a la formación de pequeñas cantidades de este mediador de la POL. Esto puede dar explicación al hecho de que los niveles de MDA fueron mayores en el lisado de espermatozoides que en el plasma seminal. Bajo condiciones fisiológicas, los niveles de ERO pueden ser neutralizados por las defensas antioxidantes.25 De igual forma, la generación de ROOH fue mayor en espermatozoides; consecuencia de los daños oxidativos a FL y proteínas de membrana que ocurren bajo condiciones fisiológicas.
Los niveles de PAOP detectados en el análisis son relativamente bajos, lo cual pudiera relacionarse con la ausencia de procesos inflamatorios en el sistema urogenital de los individuos incluidos en el estudio. Este indicador se asocia directamente a estos procesos y su comportamiento se corresponde con los valores del infiltrado de leucocitos en el espermiograma, los cuales estaban dentro del rango aceptado como normal. Se conoce que el semen eyaculado es rico en fibrinógeno, que por la acción de enzimas presentes en el mismo es convertido a fibrina, la cual le brinda la consistencia necesaria al semen para que este pueda transportar los espermatozoides hacia el sitio de la fecundación. Además existen otras proteínas como la albúmina, la carnitina y algunos aminoácidos que también son susceptibles a la oxidación. Adicionalmente la fuente generadora principal de ERO en el semen son los leucocitos, de esta forma las proteínas del plasma seminal están más expuestas a los procesos oxidativos que las que forman parte del espermatozoide.26
El organismo humano ha desarrollado una serie de estrategias antioxidantes para protegerse contra el daño oxidativo mediado por ERO. Esto permite que el metabolismo oxidativo transcurra sin que se produzcan daños que afecten la vida celular y por otra parte que las ERO puedan ejercer su papel fisiológico en mecanismos de señalización y control.27 La determinación de la actividad de enzimas antioxidantes constituye otro biomarcador de importancia en el estudio del ambiente redox celular.25
Las enzimas SOD y catalasa constituyen parte del arsenal antioxidante endógeno.25 El hecho de que en el lisado de espermatozoides la actividad de SOD sea mayor que en el plasma seminal puede deberse a que fue determinada la actividad de la SOD total. Se conoce que, desde el punto de vista filogenético, existen dos familias de esta enzima: la Cu/Zn-SOD y la Fe/Mn-SOD. La primera de ellas se localiza en el interior celular al igual que la manganeso dependiente de localización mitocondrial, mientras que existe sólo una isoforma que se localiza en la membrana de células eucariotas, responsable de la dismutación del radical O2•- en los fluidos extracelulares.25
Un comportamiento similar se observó en la actividad de la catalasa, el cual puede estar asociado a la localización, fundamentalmente intracelular de esta enzima. Ella se localiza en la membrana de los peroxisomas y en la mitocondria, aunque se ha observado en el citoplasma de algunos tipos celulares como el eritrocito.25
Otros de los componentes del sistema de defensa del organismo lo constituyen los antioxidantes de bajo peso molecular, los cuales son moléculas de naturaleza hidrofílica o lipofílica que pueden concentrarse en lugares específicos donde tiene lugar el EO25. Uno de estos antioxidantes es el GSH (L-ã-glutamil-L-cisteinil-glicina) que constituye el tiol intracelular más abundante, el cual se encuentra en el orden de los milimolares en todas las células.28 El GSH está involucrado en varias funciones celulares, como la defensa antioxidante, la cual puede llevarla a cabo de forma directa a través de la interacción con ERO o indirectamente mediante su función como cofactor de enzimas detoxificadoras como la glutatión peroxidasa.29
El PP constituye una expresión del balance entre la generación de metabolitos oxidados y su inactivación por parte de antioxidantes lipofílicos.30 Los resultados del presente trabajo sugieren que existe una mayor susceptibilidad a la POL en espermatozoides que en el plasma seminal. El a- tocoferol es el antioxidante más importante en las membranas biológicas. Este es fundamental en el mantenimiento de la integralidad y estabilidad de las mismas al proteger los AGPI de la peroxidación.25
En tanto, el ácido ascórbico, los uratos y flavonoides contenidos en el plasma seminal son la principal defensa antioxidante que se encuentra en los fluidos extracelulares de los testículos y el epidídimo una vez que el espermatozoide es eyaculado.1 La proporción de estos agentes antioxidantes pueden condicionar una mayor capacidad reductora en el plasma seminal que en el lisado celular.
Teniendo en cuenta los resultados de la presente investigación se puede concluir que en el semen de adultos jóvenes sanos existen niveles de daño a biomoléculas y de defensa antioxidante que reflejan la existencia de un EO basal. Además se concluye que existe una diferenciación entre el estado redox entre el plasma seminal y los espermatozoides, a pesar de que ambos compartimentos están en estrecha relación. A través de la implementación del diagnóstico redox personalizado se puede contribuir a la evaluación de las potencialidades de fertilidad del semen, así como la detección de alguna alteración que puede impactar negativamente sobre la capacidad reproductora del hombre.
REFERENCIAS BIBLIOGRÁFICAS
1. Tremellen K. Oxidative stress and male infertility-a clinical perspective. Hum Reprod Upd. 2008;4:1-16.
2. O´Flaherty C, de Lamirande E, Gagnon C. Positive role of reactive oxygen species in mammalian sperm capacitation: triggering and modulation of phosphorylation events. Free Radic Biol Med. 2006;41:528-40.
3. De Lamirande E, Lamothe G. Reactive oxygen-induced reactive oxygen formation during human sperm capacitation. Free Radic Biol Med. 2009;46:502-10.
4. De Jonge C. Biological basis for human capacitation. Hum Reprod. 2005;11:205-14.
5. Aziz N, Agarwal A. Evaluation of sperm damage: beyond the World Health Organization criteria. Fertil Steril. 2008;90:484-5.
6. Kierszenbaum AL. Apoptosis during spermatogenesis: the thrill of being alive. Mol Reprod Dev. 2001;58:1.
7. World Health Organization. WHO Laboratory manual for the examination of human semen and semen-cervical mucus interaction. Cambridge, UK: Cambridge University Press; 1999.
8. Marmor D, Grob-Menéndez F. Male infertility due to asthenozoospermia and flagallar anomaly: Detection in routine semen analysis. Int J Androl. 1991;14:108-16.
9. Politch JA, Wolff H, Hill JA, Anderson DJ. Comparison of methods to enumerate white blood cell in semen. Fertil Steril. 1993;60:372-5.
10. Bradford, MM. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal Biochem. 1976;72:248-54.
11. Boehringer Mannheim. Biochemica Information. A revised biochemical reference source. Enzymes for routine. 1st ed., Berlin, Germany: Boehringer Mannheim; 1987. p. 15-6.
12. Esterbauer H, Cheeseman, KH. Determination of aldehydic lipid peroxidation product: Malonaldehyde and 4-hydroxynonenal. Method Enzymol. 1990;186:407-21.
13. Witko-Sarsat V, Friedlander M, Nguyen-Khoa T, Capellére-Bladin C, Nguyen A, Canteloup S, et al. Advanced oxidation protein products as novel mediators of Inflammation and monocytes activation in chronic renal failure. J. Immunol. 1998;161:2524-32.
14. Sedlak J, Lindsay RH. Estimation of total, protein-bound, and non-protein sulfhydril groups in tissue with Ellman´s reagent. Anal Biochem. 1968;25:192-205.
15. Benzie I, Strain JJ. The ferric reducing abillity of plasma (FRAP) as a measure of antioxidant power: The FRAP Assay. Anal Biochem. 1996;239:70-6.
16. Gómez E, Aitken J. Impact of in vitro fertilization media or peroxidative damage to human espermatozoa. Fertil Esteril. 1996;65:880-2.
17. O´Brien J, Zini A. Sperm DNA integrity and male infertility. Urology. 2005;65:16-22.
18. Morrice P, Josset P, Chapron C, Dubuisson JB. History of infertility. Hum Reprod Update. 1995;1:497-504.
19. Agarwal A, Allamaneni SS. Sperm DNA damage assessment: a test whose time has come. Fertil Steril. 2005;84:850-3.
20. Aziz N, Said T, Paasch U, Agarwal A. The relationship between human sperm apoptosis, morphology and the sperm deformity index. Hum Reprod. 2007;22:1413-9.
21. Zalata AA, Christophe AB, Depuydt CE, Schoonjans F, Comhaire FH. The fatty acid composition of phospholipids of spermatozoa from infertile patients. Mol Hum Reprod. 1998;4:111-8.
22. Cerolini S, Gliozzi TM, Pizzi F, Parodi L, Maldjian A, Noble R. Importanz strutturale e funzionale delle componenti lipidichenegli spermatozoi degli animali domestici di interesse domestico. Prog Nutr. 2002;4:151-4.
23. Aitken RJ, Baker HW. Seminal leukocytes: passengers, terrorists or good samaritans? Hum Reprod. 1995;10:1736-9.
24. Gliozzi TM, Zaniboni L, Maldjian A, Luzi F, Maertens L, Cerolini S. Quality and lipid composition of spermatozoa in rabbits fed DHA and vitamin E rich diets. Theriogenology. 2009;71:910-9.
25. León OS, Martínez G, Candelario EJ, García I, Bilbao T, Ledesma L. Balance antioxidante-prooxidante: salud y enfermedad. La Habana: Ed. Ciencias Médicas; 2005. p. 2-84.
26. Agarwal A, Prabakaran SA. Mechanism, measurement, and prevention of oxidative stress in male reproductive physiology. Indian J Exp Biol. 2005;43:963-74.
27. Valko M, Leibfritz D, Moncol J, Cronin MT, Mazur M, Telser J. Free radicals and antioxidants in normal physiological function and human diseases. Int J Biochem Cell Biol. 2007;39:44-84.
28. Dickinson DA, Forman HJ. Cellular glutathione and thiols metabolism. Biochem Pharmacol. 2002;64:1019-26.
29. Circu ML, Aw Y. Glutathione and apoptosis. Free Rad Res. 2008;42:689-706.
30. Martínez G, Popov I, Pérez G, Al Dalaen SM, Horwat R, Giuliani A, et al. Contribution to characterization of oxidative stress in diabetic patients with macroangiopatic complication. Acta Farm Bonaerense. 2005;24:197-203.
Recibido: 14 de noviembre de 2012.
Aprobado: 22 de marzo de 2013.
Liván Delgado Roche. Ave. 23 # 21425 e/ 214 y 222, La Coronela, La Lisa. Ciudad de La Habana, Cuba. Correo electrónico: ldelgado@ifal.uh.cu ldelgadoroche@gmail.com


{kind=link}
{kind=link}