










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Investigaciones Biomédicas
versión On-line ISSN 1561-3011
Rev Cubana Invest Bioméd vol.34 no.4 Ciudad de la Habana oct.-dic. 2015
Rev Cubana de Investigaciones Biomédicas. 2015;34(4)
ARTÍCULO ORIGINAL
Efecto de una combinación nutritiva y selectiva con sustratos cromogénicos para diagnóstico de cocos grampositivos
Effect of a selective nutrient combination of chromogenic substrates for diagnosis of Grampositive cocci
MSc. Dennis Someillan Iglesias, DrC. Raisa Zhurbenko, DrC. Claudio Rodríguez Martínez
Centro Nacional de Biopreparados (BioCen). Bejucal. Mayabeque, Cuba.
RESUMEN
Introducción: la reemergencia de infecciones por bacterias grampositivas y el aumento de su patogenicidad, requiere de un diagnóstico microbiológico rápido y certero. En BioCen se desarrolló una composición cromogénica para el aislamiento, cultivo y diferenciación rápida y presuntiva de microorganismos grampositivos por medio de reacciones cromogénicas específicas, donde las bacterias gramnegativas se encuentran inhibidas de manera parcial o total.
Objetivo: evaluar el efecto de la combinación de bases nutritivas, inhibidores selectivos y sustratos cromogénicos para aumentar la selectivad y capacidad diferencial para especies de los géneros Enterococcus, Streptococcus y Staphylococcus de importancia clínica.
Métodos: se evaluaron 21 cepas microbianas de la American Type Culture Collection y 24 aislamientos clínicos de Streptococcus, Enterococcus y Staphylococcus y otros microorganismos gramnegativos. Se evaluaron diferentes combinaciones de bases nutritivas, acetato de talio, ácido nalidíxico y sustratos cromogénicos para la promoción del crecimiento y diferenciación de las bacterias grampositivas. Se evaluó la funcionalidad microbiológica y se le determinaron los parámetros de calidad diagnóstica.
Resultados: la combinación de bases nutritivas permitió el desarrollo de los microorganismos grampositivos, en 24 h y su diferenciación por reacciones cromogénicas específicas. El crecimiento de los microorganismos gramnegativos fue inhibido por la acción del acetato de talio (0,014 g·L-1) y ácido nalidíxico (0,008 g·L-1), excepto Proteus mirabilis y Pseudomonas aeruginosa, cuyas características morfológicas no interfieren en la diferenciación de los microorganismos diana. La sensibilidad, especificidad y exactitud diagnósticas fueron del 100 %.
Conclusión: la combinación de las bases nutritivas, los inhibidores selectivos y los sustratos cromogénicos permitió el desarrollo y diferenciación de especies de los microorganismos evaluados. La inoculación en el medio cromogénico de microorganismos diana y no diana y la diferenciación de aquellas cepas donde se detectó color similar de las colonias por medio de pruebas complementarias rápidas, le confirió al medio elevadas sensibilidad, especificidad y exactitud diagnóstica.
Palabras clave: composición cromogénica, microorganismos grampositivos, bases nutritivas, inhibidores selectivos.
ABSTRACT
Introduction: reemergence of Grampositive bacteria infections and the rise of their pathogenicity require a quick and accurate microbiological diagnosis. BioCen has developed a chromogenic composition for isolation, culturing and rapid and presumptive differentiation of gram-positive microorganisms through specific chromogenic reactions in which the inhibition of gramnegative bacteria is partial or total.
Objective: to evaluate the effect of a combination of nutrient bases, selective inhibitors and chromogenic substrates to increase the selectivity and differential capacity to detect Enterococcus, Streptococcus and Staphylococcus species of clinical importance.
Methods: twenty one microbial strains from the American Type Culture Collection and 24 clinical isolates of Enterococcus, Streptococcus and Staphylococcus and of other gramnegative microorganisms were evaluated. Various combinations of nutrient bases, thallium acetate, nalidixic acid and chromogenic substrates were also assessed for the promotion, growth and differentiation of grampositive bacteria. The microbiological functionality was evaluated whereas the diagnostic quality parameters were determined.
Results: the combination of nutrient bases allowed the development of grampositive microorganisms in 24 hours and their differentiation through specific chromogenic reactions. The growth of gramnegative microorganisms was inhibited by the thallium acetate (0.014 g·L-1) and nalidixic acid (0,008 g·L-1) except for Proteus mirabilis and Pseudomonas aeruginosa whose morphological characteristics do not interfere with differentiation of target microorganisms. Sensitivity, specificity and accuracy for diagnosis were 100 %.
Conclusions: the combination of nutrient bases, selective inhibitors and chromogenic substrates allowed the development and differentiation of the evaluated microorganism species. The inoculation of target and non-target microorganisms in the chromogenic medium and the differentiation of those strains where a similar color of the colonies was detected by means of supplementary rapid tests provided the medium with high diagnostic sensitivity, specificity and accuracy.
Keywords: chromogenic composition, grampositive microorganisms, nutrient bases, selective inhibitors.
INTRODUCCIÓN
En los últimos años se observa un incremento de las infecciones por bacterias grampositivas, con una alta incidencia y prevalencia en disímiles afecciones de origen comunitario y nosocomial, con serias implicaciones sociales y económicas.1-8
Disímiles factores han originado cepas con alto potencial patogénico dentro de este grupo donde los cambios en la virulencia y en la resistencia a los antimicrobianos, derivados del uso indiscriminado de antibióticos provocan una situación alarmante a nivel mundial producto de la reemergencia de infecciones graves producidas por estos agentes y una limitada eficacia en el tratamiento.9-13
Esta situación condiciona la investigación y desarrollo de composiciones nutricionales que favorezcan el cultivo e identificación de los microorganismos grampositivos mediante el diagnóstico rápido y certero que, a su vez, posean altos indicadores de sensibilidad y especificidad.14
Los métodos convencionales han demostrado su factibilidad a la hora de aislar e identificar microorganismos grampositivos de disímiles muestras clínicas como ambientes ecológicos, pero no los exentes de ser laboriosos y de consumir mucho tiempo, puesto que se necesitan de pruebas bioquímicas complementarias o serológicas para la confirmación de estos agentes.
El empleo de medios cromogénicos resulta factible por su sencillez, elevada sensibilidad, especificidad diagnóstica y rápida interpretación de los resultados sobre la base de la morfología y color de las colonias en el medio, a partir de la detección de actividades enzimáticas específicas.5,15,16 Estos medios requieren de menos tiempo de trabajo y logran un resultado presuntivo, específico, y en algunos casos confirmatorio mediante pruebas enzimáticas rápidas complementarias, en un tiempo menor a las 48 h.14,17
En BioCen, en estudios anteriores,17 se desarrolló una composición cromogénica para el aislamiento, cultivo y diferenciación rápida y presuntiva de microorganismos grampositivos, tales como Enterococcus, Streptococcus y Staphylococcus; por medio de reacciones específicas proporcionadas por enzimas y sustratos cromogénicos. En esta composición los microorganismos diana proporcionaban colonias específicas pero de menor tamaño en comparación con las desarrolladas en el medio de referencia agar sangre.
El objetivo de esta investigación consiste en evaluar el efecto de la combinación de bases nutritivas, inhibidores selectivos y sustratos cromogénicos para aumentar su capacidad nutritiva, selectiva y diferencial para especies de los géneros Enterococcus, Streptococcus y Staphylococcus de importancia clínica.
MÉTODOS
Se emplearon bases nutritivas de producción nacional (BioCen, Cuba) como hidrolizado enzimático de caseína (HEC), peptona bacteriológica S (PBS), peptona de soya (PS) y extracto de levadura (EL). El extracto de carne (EC) utilizado fue de Conda (España).
Se utilizó una mezcla de acetato de talio (AT) y de ácido nalidíxico (AN) (Sigma-Aldrich, EE. UU.) como inhibidores selectivos de bacterias gramnegativas y diferentes sustratos cromogénicos como el 5-bromo-4-cloro-3-indoxil-beta-D-glucurónido, sal de ciclohexilamonio (X-gluc), el 6-cloro-3-indoxil-beta-D-glucopiranósido (rosa-glu) y paranitrofenilfosfato sal disódica (PNP) (Apollo, Inglaterra) para diferenciar los microorganismos estudiados a partir del color desarrollado por las colonias luego de 24 h de incubación a 35 ± 2 °C y los cambios ocurridos en la superficie del agar de la composición cromogénica.
La DL-fenilalanina, citrato férrico, D(+)-glucosa (Merck, Alemania) se emplearon como indicadores de reacciones bioquímicas en el medio. Las sales cloruro de sodio, fosfato dipotásico y fosfato disódico procedieron de AppliChem (Alemania) y se utilizaron para garantizar la presión osmótica, estabilizar el pH del medio de cultivo y como disolvente del suplemento líquido estéril (PNP), en orden. La tierra silícea (Merck, Alemania) ejerció papel de sustancia contrastante, mientras que el agar bacteriológico A (AgarMex, México) como agente gelificante.
Evaluación microbiológica
Se empleó un total de 45 cepas bacterianas (tabla 1), de ellas 21 de referencia clasificadas según la American Type Culture Collection (ATCC) y 24 aisladas de muestras clínicas de exudados de pacientes procedentes de la consulta médica de BioCen (tabla 1). Las cepas se almacenaron en el medio agar nutriente a temperatura de 2 a 8 °C. Los microorganismos se inocularon en tubos de cristal con tapa de rosca que contenían caldo triptona soya. Se incubaron de 18-24 h a 35 ± 2 °C en aerobiosis y se procedió a estandarizar el inóculo. La alícuota de la suspensión bacteriana se añadió a un tubo con solución salina estéril al 0,85 % (p/v) y se ajustó la concentración microbiana a DO580 de 0,25 en un espectrofotómetro (T70/T70+UV-VIS, England), que corresponde a 3,0 x 108 UFC·mL-1. Con un asa calibrada 0,1 mL de la solución estandarizada del inóculo, se colocó en la superficie del medio cromogénico experimental (MC) y del medio control agar sangre (AS) (BioCen, Cuba), este último preparado con 5 % de sangre de carnero desfibrinada. La siembra se realizó por estriado por agotamiento del inóculo en la superficie del agar de ambos medios para obtener colonias aisladas. Los medios se incubaron de 18-24 h a 35 ± 2 °C en aerobiosis.
Tabla 1. Microorganismos utilizados en el estudio
| Microorganismo | Procedencia | Cantidad | Corrida experimental | ||||
| Grupo 1 | Grupo 2 | Grupo 3 | |||||
| Streptococcus pyogenes 19615 | ATCC | 1 | x | x | x | ||
| Streptococcus agalactiae 12136 | ATCC | 1 | x | x | x | ||
| Streptococcus canis 43496 | ATCC | 1 | x | - | x | ||
| Streptococcus pneumoniae 49619 | ATCC | 1 | x | x | x | ||
| Staphylococcus aureus 25923 | ATCC | 1 | x | x | x | ||
| Staphylococcus saprophyticus 15305 | ATCC | 1 | x | - | x | ||
| Staphylococcus epidermidis 12228 | ATCC | 1 | x | - | x | ||
| Staphylococcus xylosus 27971 | ATCC | 1 | x | - | x | ||
| Staphylococcus haemolyticus 29770 | ATCC | 1 | x | - | x | ||
| Enterococcus faecalis 29212 | ATCC | 1 | x | x | x | ||
| Enterococcus faecium 19434 | ATCC | 1 | x | - | x | ||
| Enterococcus avium 14025 | ATCC | 1 | x | - | x | ||
| Enterococcus casseliflavus 70327 | ATCC | 1 | x | - | x | ||
| Pseudomonas aeruginosa 27853 | ATCC | 1 | x | - | x | ||
| Proteus mirabilis 12453 | ATCC | 1 | x | - | x | ||
| Proteus vulgaris 13315 | ATCC | 1 | x | - | x | ||
| Escherichia coli 25922 | ATCC | 1 | x | - | x | ||
| Enterobacter aerogenes 13048 | ATCC | 1 | x | - | x | ||
| Klebsiella pneumoniae 13883 | ATCC | 1 | x | - | x | ||
| Salmonella Typhimurium 14028 | ATCC | 1 | x | - | x | ||
| Citrobacter freundii 8090 | ATCC | 1 | x | - | x | ||
| Aislados de muestras clínicas | |||||||
| S. pyogenes | F | 4 | - | - | x | ||
| L | 1 | - | - | x | |||
| S. agalactiae | F | 1 | - | - | x | ||
| V | 1 | - | - | x | |||
| S. aureus | F | 2 | - | - | x | ||
| L | 8 | - | - | x | |||
| N | 1 | - | - | x | |||
| Ó | 1 | - | - | x | |||
| P. aeruginosa | F | 1 | - | - | x | ||
| P. mirabilis | V | 1 | - | - | x | ||
| E. coli | V | 1 | - | - | x | ||
| OR | 2 | - | - | x | |||
| Total | 45 | 21 | 5 | 45 | |||
F: exudado faríngeo; L: exudado de la lesión de piel; V: exudado vaginal; N: exudado nasal; O: exudado ótico; OR: muestras de orina; grupo 1: evaluación de los inhibidores selectivos en la composición cromogénica; grupo 2: evaluación de bases nutritivas combinadas para la promoción de bacterias grampositivas; grupo 3: evaluación de la funcionalidad microbiológica de la composición cromogénica.
Se realizó, en todo el estudio, la inspección visual del crecimiento bacteriano en las placas de los medios de cultivos empleados y de las características culturales de las colonias desarrolladas en la superficie del agar luego de la incubación. Los aspectos analizados fueron: tamaño, forma y color desarrollados según el sustrato cromogénico utilizado por el microorganismo. En el AS se observaron las mismas características de los cultivos más la producción de hemólisis por los microorganismos ensayados.
Para el estudio de promoción de crecimiento microbiano se emplearon curvas de crecimiento construidas a partir de los valores de la densidad óptica obtenida del incremento de la absorbancia de los cultivos a DO640 en un espectrofotómetro (T70/T70 + UV-VIS, England). Las absorbancias se midieron por triplicado al inicio y posterior, cada una hora durante las primeras 10 h y a las 24 h de incubación a 35 ± 2 °C.
Evaluación de los inhibidores selectivos en la composición cromogénica
Se evaluaron distintas concentraciones de los inhibidores selectivos para microorganismos gramnegativos no objeto de estudio: variante uno (V1: AT 0,3 g·L-1 y AN 0,013 g·L-1), variante dos (V2: AT 0,02 g·L-1 y AN 0,008 g·L-1), variante tres (V3: AT 0,016 g·L-1 y AN 0,008 g·L-1) y variante cuatro (V4: AT 0,014 g·L-1 y AN 0,008 g·L -1).
Los restantes ingredientes correspondieron a la composición inicial del medio cromogénico.17 Al medio preparado se le ajustó el pH a 7,3 ± 0,2 y luego de esterilizado se le adicionaron para 1 L del MC 12 mL de una solución estéril al 0,1 % de PNP (p/v) en una solución de fosfato disódico 0,01 M. Se inocularon los microorganismos identificados como Grupo 1 (tabla 1).
Evaluación de bases nutritivas combinadas para la promoción de bacterias grampositivas
Se evaluaron tres combinaciones de bases nutritivas en distintas concentraciones: V1- HEC 9,97 g·L-1, PBS 6,37 g·L-1, PS 3,24 g·L-1, EL 3,24 g·L-1 y EC 2,43 g·L-1; V2- HEC 5,0 g·L-1, PBS 6,0 g·L-1, PS 5,0 g·L-1, EL 5,0 g·L-1 y EC 3,0 g·L-1 y V3- HEC 3,0 g·L-1, PBS 6,0 g·L -1, PS 6,0 g·L-1, EL 6,0 g·L-1 y EC 4,0 g·L-1. A ellas se adicionaron cloruro de sodio (0,5 g·L-1) y fosfato disódico (0,1 g·L-1), con un pH entre 7,0-7,2, según SIGMA.18 En todas se inocularon las cepas de la ATCC descritas en el Grupo 2 (tabla 1) en representación de los microorganismos grampositivos objeto de estudio.
Evaluación de la funcionalidad microbiológica de la composición cromogénica
Se confeccionó una composición final a partir de los resultados obtenidos de la evaluación de los inhibidores selectivos de microorganismos gramnegativos y de las bases nutritivas combinadas para la promoción de bacterias grampositivas. En esta composición se evaluaron las cepas microbianas referidas como Grupo 3 (tabla 1).
Se realizó la tinción de Gram con el juego de reactivos Empresa de Productos Biológicos (EPB) “Carlos J. Finlay, Cuba y se aplicaron pruebas de catalasa (AppliChem, Alemania) y PYR (Oxoid, England) para la diferenciación de los microorganismos desarrollados en el MC con las características cromogénicas similares.
Determinación de los parámetros diagnósticos de calidad
La sensibilidad (S), especificidad (E) y exactitud diagnóstica (ED), expresadas en porcentaje, se calcularon según la norma ISO 2000: 19 S [(VP/VP+FN) x 100], E [(VN/VN+FP) x 100] y ED [(VP+VN/VP+VN+FN+FP) x 100], donde: VP - verdaderos positivos, VN – verdaderos negativos, FP – falsos positivos y FN – falsos negativos.
Análisis estadístico
El procesamiento de los datos durante el estudio de promoción de crecimiento bacteriano a través del aumento de la densidad óptica en el tiempo se realizó según el programa estadístico STATISTICA (versión 8; Copyright, Statsoft, Inc. 2008). Se determinó la existencia de diferencia significativa entre los valores de la densidad óptica correspondientes, a cada combinación de bases nutritivas, se empleó la prueba de Tukey, para un 95 % de confianza.
RESULTADOS
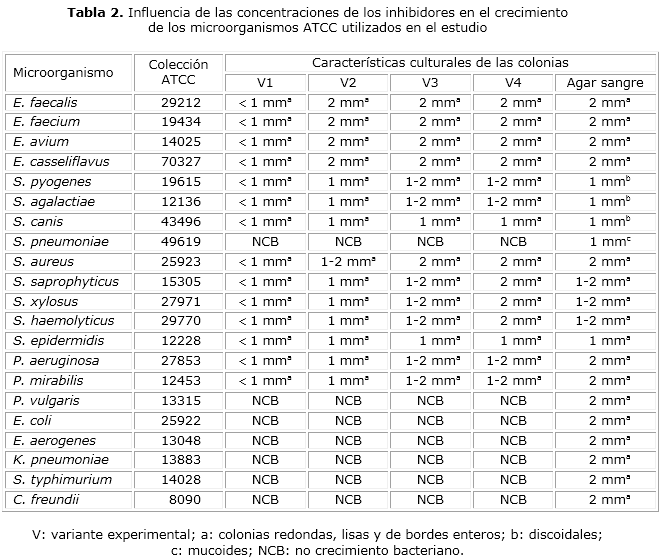
Las colonias de los microorganismos grampositivos de la ATCC, pertenecientes a los géneros Streptococcus, Enterococcus y Staphylococcus durante el estudio de los inhibidores en el MC resultaron de diferentes tamaños (tabla 2). No se evidenció crecimiento de S. pneumoniae 49619 en el MC mientras que en el AS se observó desarrollo con colonias mucoides de 1 mm de diámetro. Tanto P. mirabilis 12453 como P. aeruginosa 27853 mostraron crecimiento con colonias que de menos de 1 mm de diámetro en la V1, pasaron a ser entre 1 y 2 mm en las demás variantes, mientras que el resto de los microorganismos gramnegativos estudiados (P. vulgaris 13315, E. coli 25922, E. aerogenes 13048, K. pneumoniae 13883, S. Typhimurium 14028 y C. freundii 8090) no evidenciaron desarrollo microbiano. En el medio AS la totalidad de los microorganismos gramnegativos mostraron colonias de 2 mm de diámetro (tabla 2).
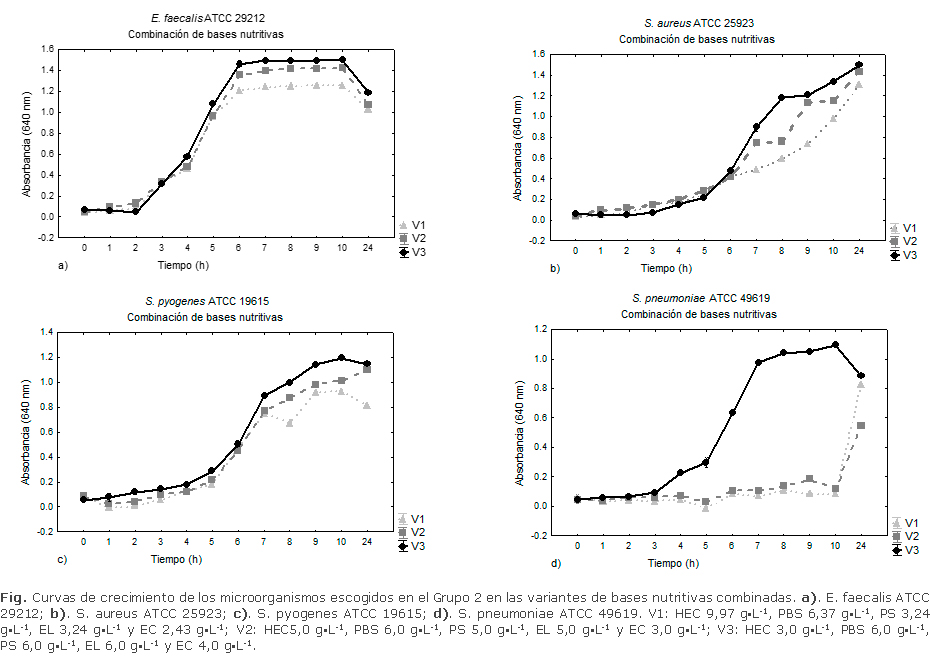
La figura muestra las curvas de crecimiento de los microorganismos diana en diferentes combinaciones de bases nutritivas. E. faecalis ATCC 29212 (Fig. a) no mostró diferencias significativas (p< 0,05) en las tres primeras horas en las diferentes variantes de las bases combinadas. A partir de la cuarta hora el microorganismo comenzó su crecimiento acelerado, alcanzó la fase exponencial hasta la sexta hora. En este tiempo se observaron diferencias significativas (p< 0,05) entre las variantes, mostró la V3 mayor incremento de la biomasa. Desde la séptima hora y hasta las 24 h el microorganismo mantuvo un crecimiento constante en todas las variantes. La V2 manifestó superioridad a la V1 en cuanto al incremento de la biomasa, mientras que V3 resultó superior al resto.
Para S. aureus ATCC 25923 (Fig. b), en las primeras seis horas de cultivo se observó crecimiento lento de este microorganismo, sin existir diferencias significativas (p< 0,05) entre las variantes. El crecimiento acelerado se apreció a partir de la séptima hora y hasta la décima, se mostró diferencias significativas de la densidad óptica entre todas las variantes (p< 0,05), pero con valores superiores en V3.
El crecimiento acelerado de S. pyogenes ATCC 19615 (Fig. c) se evidenció en todas las variantes a partir de la quinta hora. En este tiempo V3 mostró una ligera superioridad, con diferencias significativas (p< 0,05) al resto de las variantes a partir de la sexta hora, mientras que V2 fue superior a V1. Desde la séptima hora hasta las 24 h, se observaron diferencias significativas (p< 0,05) en los valores de absorbancia entre todas las variantes, mostrando superioridad V3.
La curva de crecimiento de S. pneumoniae ATCC 49619 (Fig. d) mostró que en las primeras tres horas el microorganismo se encontró en la fase lag de su desarrollo en todas las variantes. A partir de la tercera hora y hasta la decima, se apreció un desarrollo acelerado del microorganismo en V3, mostró diferencia significativa (p< 0,05) y una superioridad absoluta al resto de las variantes. Tanto V1 como V2 mostraron una desaceleración del crecimiento hasta la décima hora en contraste con V3. Aunque V1 y V2 mostraron un aumento del crecimiento de S. pneumoniae a partir de la décima hora, a las 24 h de incubación se observó el predominio de V3.
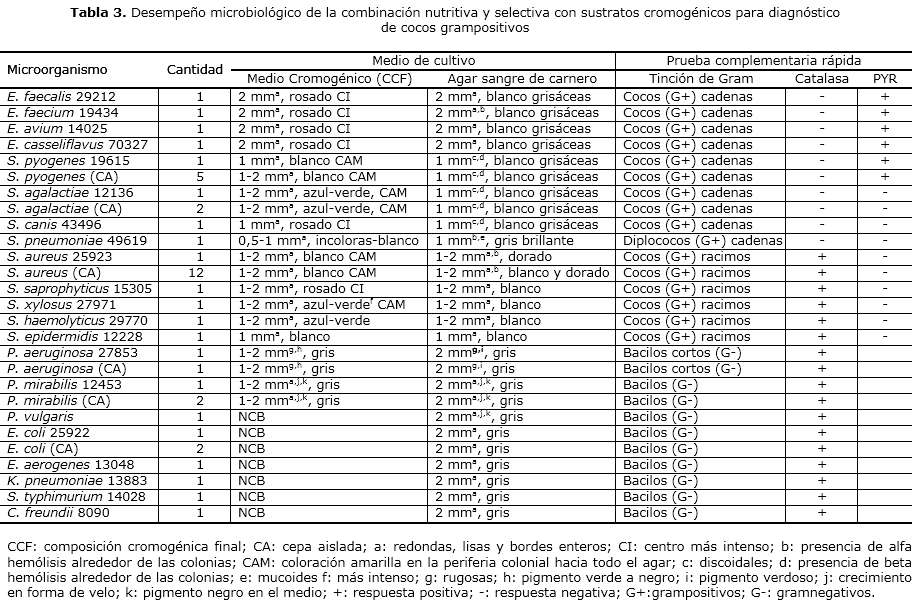
Para todas las especies de los microorganismos grampositivos se obtuvo el tamaño de colonias similar y, en algunos casos, superior al obtenido en el medio control (AS); otras características morfológicas (forma, color y hemólisis) resultaron ser las esperadas para cada uno de los medios de cultivo (MC y AS) (tabla 3).
En el MC las cepas ATCC P. mirabilis 12453 y P. aeruginosa 27853 y las cepas aisladas de estas bacterias mostraron colonias entre 1-2 mm (inferior a las desarrolladas en el AS), redondas, lisas y con bordes enteros. Para el resto de los microorganismos gramnegativos estudiados, no se observó crecimiento bacteriano (tabla 3).
Las características cromogénicas de las especies de los géneros grampositivos y gramnegativos ATCC y aislados de muestras clínicas se observan en la tabla 3. En el MC las colonias de color rosado con centro más intenso y pruebas bioquímicas rápidas correspondieron a las colonias de especies de la ATCC del género Enterococcus (PYR positivo), S. canis 43496 (PYR y catalasa negativos) y S. saprophyticus 15305 (PYR negativo y catalasa positivo).
El desarrollo de un color azul-verde en las colonias se produjo por las siguientes cepas de la ATCC y aisladas, quedó diferenciadas por la producción o no de un color amarillo desde la periferia colonial hacía el medio y pruebas rápidas como catalasa y PYR: S. agalactiae ATCC 12136 (color amarillo y catalasa negativo), S. xylosus 27971 (color amarillo y catalasa positivo) y S. haemolyticus 29770 (catalasa positivo).
Las colonias de color blanco pertenecieron a las siguientes cepas de la ATCC o aisladas, que fueron diferenciadas por el desarrollo o no de color amarillo alrededor del crecimiento bacteriano y la prueba rápida de catalasa: S. pyogenes 19615 y las cinco cepas aisladas de muestras clínicas (color amarillo y catalasa negativo), S. aureus 25923 y 12 aislamientos de especímenes clínicos (color amarillo y catalasa positivo), S. epidermidis 12228 (catalasa positivo) y S. pneumoniae 49619 (de incoloras a blancas y catalasa negativo).
En el MC P. mirabilis ATCC 12453 y las cepas aisladas de esta bacteria mostraron colonias de color gris, con crecimiento en forma de velo y pigmento negro en el medio. En el caso de P. aeruginosa ATCC 27853 y su aislamiento clínico se observaron colonias de similar tamaño y color que P. mirabilis, pero de aspecto rugoso con pigmento verde a negro en las colonias.
El análisis de los parámetros diagnósticos de calidad del MC para las 45 cepas microbianas empleadas en la investigación mostró que 32 resultaron verdaderos positivos y 13 verdaderos negativos. No se obtuvieron falsos positivos ni falsos negativos por lo que la sensibilidad, especificidad y exactitud diagnósticas alcanzó valores del 100 %.
DISCUSIÓN
El empleo del acetato de talio en la formulación del MC respondió a la necesidad de sustituir la azida de sodio debido a sus propiedades cancerígenas y teratogénicas, a pesar de ser esta un inhibidor selectivo de bacterias gramnegativas, de amplio uso en medios de cultivo para el aislamiento de microorganismos grampositivos.20-22
El AT es un reactivo tóxico, pero menos nocivo que el sulfato de talio y que otras sustancias, que son empleadas en diferentes composiciones cromogénicas y fluorogénicas.22 El mismo es efectivo para lograr la inhibición total o parcial de los microorganismos no diana (gramnegativos) debido a que posee la propiedad de acumularse en la célula y competir con el potasio y reemplazarlo, modificando la activación enzimática de la ATPasa de Na+/K+. También interfiere con la estabilidad de los ribosomas y la síntesis de proteínas y con sitios activos de varias enzimas y su interacción con los grupos amino-sulfhidrilo en este grupo bacteriano.23
Kumiko Kohguchi y colaboradores, en 1969,24 plantearon que el empleo de concentraciones de 0,01-0,5 g·L-1 de AT no influye en el crecimiento de los microorganismos grampositivos. En el presente estudio, cantidades superiores a 0,014 g·L-1, de este inhibidor, combinado con el AN afectaron el desarrollo de los microorganismos diana en el MC, expresado en el tamaño de las colonias (tabla 2).
El ácido nalidíxico sal disódica se utilizó en la investigación por ser empleado con frecuencia en medios de cultivo selectivos debido a su poder inhibitorio sobre el crecimiento de bacterias gramnegativas, en especial de enterobacterias y Pseudomonas, por su capacidad de interferir en la síntesis del ADN bacteriano.25 Concentraciones superiores a 0,008 g·L-1 de AN (0,013 g·L-1 en V1-V3) afectan el crecimiento de S. pyogenes y especies de estafilococos; sin embargo, una concentración de AN en 0,008 g·L -1, combinada con 0,014 g·L-1 de AT, proporcionó el tamaño colonial esperado (entre 1-2 mm de diámetro en el MC, similar al medio AS).
La disminución de ambos inhibidores en el MC y el establecimiento de las concentraciones definidas en V4, favoreció el crecimiento de los microorganismos diana. Lo anterior está en plena correspondencia con lo referido por otros autores,26 sobre la efectividad de una correcta combinación de inhibidores de manera general.
El microorganismo más afectado en su crecimiento resultó ser S. pneumoniae. Esta bacteria tiene requerimientos nutricionales especiales, llegando a ser una de las más fastidiosas dentro del grupo ya que requiere de nutrientes muy específicos que garanticen su desarrollo27 y posterior crecimiento en los medios de cultivo. Fue necesario proceder a un estudio pormenorizado con el objetivo de definir las concentraciones adecuadas de bases nutritivas que permitieran su crecimiento satisfactorio.
El aumento de las concentraciones de PS, EL y EC en el MC, y la disminución en las PBS y HEC (V3) incrementaron el desarrollo de los microorganismos ensayados, es el más favorecido, S. pneumoniae (Fig.) debido a la alta calidad de las proteínas de soya y su valor energético, unido al amplio espectro de sustancias nutritivas y el elevado contenido de carbohidratos y vitaminas de la PS, así como el alto contenido en vitaminas del complejo B, aminoácidos libres, péptidos, carbohidratos y minerales del EL.28,29
La presencia del EC en concentraciones superiores a los 2 g·L-1 en la composición cromogénica responde a una compilación de los resultados obtenidos en este estudio en conjunto con investigaciones anteriores realizadas por Durán Vila y Rodríguez Martínez,18 que demuestran una mayor promoción en el crecimiento de bacterias exigentes a una concentración de esta base nutritiva de 4 g·L-1. Esto se justifica en el aporte considerable de esta base en proteínas hidrosolubles, aminoácidos, péptidos, purinas, guanidinas, ácidos orgánicos, vitaminas, minerales y otros compuestos orgánicos, también referidos por Zhurbenko y Rodríguez.30
La poca variabilidad en la concentración de la PBS en las variantes realizadas se debió a que la misma debe ser utilizada en concentraciones altas en los medios, si se desea aislar microorganismos exigentes, según refiere Osmanov, en 1983.31
El HEC (triptona) es una base nutritiva de suma importancia para los microorganismos de lento crecimiento.32 Fue necesario disminuir su concentración hasta 4 g·L-1 debido a que cantidades superiores de la misma, no aportaron al crecimiento de S. pneumoniae. Las combinaciones de la triptona (4 g·L-1), con otras bases nutritivas empleadas en este estudio suplió los requerimientos metabólicos de estos microorganismos nutricionalmente exigentes. Estos resultados concuerdan con los hallazgos obtenidos por otros autores, los cuales demuestran la conveniencia de utilizar las mezclas de peptonas, hidrolizados y extractos en medios de cultivo destinados a este tipo de microorganismos.33
La presencia en la formulación de los sustratos cromogénicos (rosa-glu, x-gluc y PNP) permitió detectar la actividad de diferentes enzimas tales como beta-glucosidasa, beta-glucuronidasa y fosfatasa alcalina en las células microbianas, lo que garantizó la diferenciación de las bacterias grampositivas por el color desarrollado en las colonias y en el medio alrededor de las colonias, luego de 18 a 24 h de incubación (tabla 3). Esta diferenciación se debe a la capacidad que tiene una enzima específica de romper el enlace entre el sustrato específico de la reacción enzimática y un grupo cromóforo unido a dicho sustrato, que provoca un cambio de color en las colonias desarrolladas en la superficie del MC.15
Tanto las especies de enterococos como S. saprophyticus y S. canis poseen actividad beta-glucosidasa capaz de degradar el sustrato rosa-glu, evidenciado en el color rosado adquirido por las colonias en el MC. S. agalactiae, S. xylosus y S. haemolyticus demuestran actividad beta-glucuronidasa por el color azul-verde desarrollado por las colonias al hidrolizar el sustrato x-gluc presente en el medio. Se comprobó la actividad fosfatasa alcalina de S. agalactiae y S. xylosus ya que ambos mostraron una coloración amarilla intensa, alrededor de las colonias y en el resto de la superficie del MC, debido a la utilización del sustrato PNP que tiene la capacidad de producir un pigmento, que difunde por la superficie del agar. La misma actividad fosfatasa posee S. pyogenes y S. aureus al observarse una coloración amarilla similar en el medio. El color blanco de las colonias de ambas especies, así como de S. pneumoniae y S. epidermidis, demostró que estos microorganismos no tienen la capacidad de utilizar los sustratos rosa-glu y x-gluc.
Aunque S. pyogenes y S. aureus mostraron características cromogénicas similares se diferenciaron en la tinción de Gram, ubica al pimero en la familia Streptococcaceae y Micrococcaceae. La prueba de catalasa permitió situarlos en el género Streptococcus y Staphylococcus y la demostración de la enzima PYR, en S. pyogenes fue un ensayo adicional que permitió confirmar de manera presuntiva a este microorganismo y diferenciarlo de S. aureus. Las mismas pruebas, excepto la de PYR, permitieron la diferenciación de S. agalactiae y S. xylosus y la ubicación taxonómica correcta en cuanto a familia y género bacteriano descrita en la literatura.34 Tanto la coloración de Gram como la prueba de catalasa garantizaron la diferenciación de S. saprophyticus de las especies de enterococos y S canis. Estas dos últimas se diferenciaron con la aplicación de la prueba de PYR, fue positiva para las especies de enterococos y negativa para S. canis.
Los indicadores de la calidad diagnósticos resultaron en todos los casos superiores al 90 %, y correspondió a los requisitos establecidos en la norma ISO, 2000.20 El MC fue capaz de detectar los microorganismos diana ATCC inoculados y los que se encontraban presentes en muestras clínicas luego de 24 h de incubación. Los microorganismos no diana no crecieron en el medio excepto las cepas de P. mirabilis y P. aeruginosa, las cuales fueron clasificadas como negativas, lo cual garantiza una exactitud satisfactoria en el diagnóstico de los microorganismos grampositivos.
La combinación en el medio de los inhibidores selectivos, las bases nutritivas y los sustratos cromogénicos permitió el desarrollo y diferenciación de especies de estreptococos, estafilococos y enterococos. La inoculación en el MC de microorganismos diana y no diana y la diferenciación de aquellas cepas donde se detectó color similar de las colonias por medio de pruebas complementarias rápidas, le confirió al medio una sensibilidad, especificidad y exactitud diagnóstica satisfactoria con una alta competitividad.
REFERENCIAS BIBLIOGRÁFICAS
1. Liu C, Bayer A, Cosgrove SE, Daum RS, Fridkin SK, Gorwitz RJ, et al. Clinical Practice Guidelines by the Infectious Diseases Society of America for the treatment of methicillin-resistant Staphylococcus aureus infections in adults and children: Executive summary. Clin Infect Dis. 2011;52(3):285-92.
2. Arias CA, Murray BE. The rise of Enterococcus: beyond vancomycin resistance. Nat Rev Microbiol. 2012;10(4):266-78.
3. Fariña N, Carpinelli L, Samudio M, Guillén R, Laspina F, Sanabria R, et al. Staphylococcus coagulasa-negativa clínicamente significativos. Especies más frecuentes y factores de virulencia. Rev Chil Infectol. 2013;30(5):480-8.
4. Parks T, Barrett L, Jones N. Invasive streptococcal disease: a review for clinicians. British Medical Bulletin. 2015 [citado 10 Jun 2015]; p. 1-13. Disponible en: http://bmb.oxfordjournals.org/
5. Cantón R, Ruiz-Garbajosa P. Infecciones causadas por bacterias grampositivas multiresistentes (Staphylococcus aureus y Enterococcus spp.). Enferm Infecc Microbiol Clin. 2013;31(8):543-51.
6. Pérez Montoya LH, Zurita Villarroel IM, Pérez Rojas N, Patiño Cabrera N, Calvimonte OR. Infecciones Intrahospitalarias: Agentes, manejo actual y prevención. Rev Cient Cienc Méd. 2010;13(2):94-8.
7. Zhang R, Wang F, Kang J, Wang X, Yin D, Dang W, et al. Prevalence of multidrug resistant Gram-positive cocci in a Chinese hospital over an 8-year period. Int J Clin Exp Med. 2015;8(6):9462-9.
8. Hughes GJ, van Hoek AJ, Sriskandan S, Lamagni TL. The costs of invasive group A streptococcal infections in England. Epidemiol Infect. 2015;143:1719-30.
9. Huttner A, Harbarth S, Carlet J, Cosgrove S, Goossens H, Holmes A, et al. Antimicrobial resistance: a global view from the 2013 World Healthcare-Associated Infections Forum. Antimicrobial Resistance and Infection Control. 2013 [citado 7 May 2015];2(1):31. Disponible en: http://www.aricjournal.com/content/2/1/31
10. Centers for Disease Control and Prevention. Antibiotic resistance threats in the United States. CDC Atlanta (EE. UU): GA; 2013.
11. Pujol M, Limón E. Epidemiología general de las infecciones nosocomiales. Sistemas y programas de vigilancia. Enferm Infecc Microbiol Clin. 2013;31(2):108-13.
12. Reglinski M, Sriskandan S. The contribution of group A streptococcal virulence determinants to the pathogenesis of sepsis. Virulence. 2014;5(1):127-36.
13. Khan HA, Ahmad A, Mehboob R. Nosocomial infections and their control strategies. Asian Pac J Trop Biomed. 2015;5(7):509-14.
14. Díaz Pérez M, Zhurbenko R, Fuentes Bárcenas M, Hernández Cortez C, Castro-Escarpulli G, Rodríguez Martínez C, et al. Evaluation of an alternative chromogenic method for the detection and enumeration of enterococci in waters. Afr J Microbiol Res. 2014;8(7):652-8.
15. Manafi M, Kneifel W, Bascomb S. Fluorogenic and chromogenic substrates used in bacterial diagnostics. Microbiol Rev. 1991;55(3):335-48.
16. Orenga S, James AL, Manafi M, Perry JD, Pincus DH. Enzymatic substrates in microbiology. J Microbiol Meth. 2009;79(2):139-55.
17. Durán Vila A, Rodríguez Martínez C. Inventors; National Center of Biological products, assignee. Selective culture medium for the isolation and/or detection of species in the Streptococcus genus. EP patent 1 600 514 B1. 2011 Ago 31.
18. Sigma A. Biochemicals and Reagents for like Science Research. St. Louis (EE. UU.): Sigma-Aldrich; 2000.
19. ISO 13843 (2000). Water quality – Guidance on validation of microbiological methods, ISO/TR 13843. Geneva, Switzerland: International Organization for Standarization; 2000.
20. Reuter G. Selective media for group D-Streptococci. Int J Food Microbiol. 1985;2(1-2):103-14.
21. Koneman EW, Allen SD, Janda WM, Schreckenberger PC, Winn WC Jr. Color atlas and textbook of Diagnostic Microbiology. 5th ed. New York, Philadelphia: Lippincott; 1997.
22. Díaz Pérez M, Rodríguez Martínez C, Zhurbenko R. Enterococcus, medios de cultivo convencionales y cromogénicos. Rev Cubana Hig Epidemiol. 2013;51(1):97-110.
23. Rodríguez JJ, Altamirano MA. Genetic toxicology of thallium: a review. Drug Chem Toxicol 2013;36:369-83.
24. Konguchi K, EDE K-I, Sagara Y, Nakamura M. Effect of thallium acetate on the growth of bacteria. Kurume Med J 1969;16(3):163-8.
25. Calvo J, Martínez-Martínez L. Mecanismos de acción de los antimicrobianos. Enferm Infecc Microbiol Clin. 2009;27(1):44-52.
26. Power DA, McCuen PJ. Manual of BBL Products and Laboratory Procedures. 6th ed. Cockeysville, MD: Becton Dickinson Microbiology Systems; 1988.
27. Gobbetti M, Calasso M. Streptococcus. En Batt CA, Tortolello ML (Ed). Encyclopedia of food microbiology. 2nd ed. Elsevier Ltd. 2014;3:535-54.
28. Hongfei Z, Fengling B, Fang Z, Walczak P, Xiangning J, Bolin Z, et al. Characterization of soybean protein hydrolysates able to promote the proliferation of Streptococcus thermophilus ST. J Food Sci. 2013;78(4):575-81.
29. Zhurbenko R, Rodríguez Martínez C, Díaz Pérez M, Durán Vila A, López Hernández OD, Viera Oramas DR, et al. Caracterización de la peptona de soya para el cultivo de microorganismos. Rev Cubana Med Trop. 2006;58(2):109-18.
30. Zhurbenko R, Rodríguez Martínez C. Bases nutritivas para el cultivo de los microorganismos: Parte 1. Procesos tecnológicos. Salud (i) Ciencia. 2008;16(4):420-5.
31. Osmanov SK. Poluchenie I experimentalnoie izuchenie pitatielnikj sried s gidrlizatami krovi zhivtnikj dlia mikrobiologicheskoi diagnostiki streptkokkovikj infektsii (Tesis doctoral). Moskva: MNIIVSIIIM; 1983.
32. Meli F, Lazzi C, Neviani E, Gatti M. Effect of protein hydrolysates on growth kinetics and aminopeptidase activities of Lactobacillus. Curr Microbiol. 2014;68:82-7.
33. Tsoraeva A, Zhurbenko R. Development and Characterization of a Mixed Nutrient Base for the Culture of a Wide Range of Microorganisms. Rev Latinoam Microbiol. 2000;42:155-61.
34. MacFaddin JF. Pruebas Bioquímicas para la identificación de Bacterias de importancia clínica. Tomo I. 3era edición. Ciudad de La Habana: Ed. Ciencias Médicas; 2006.
Recibido: 2 de agosto de 2015.
Aprobado: 30 de agosto de 2015.
Dennis Someillan Iglesias. Centro Nacional de Biopreparados (BioCen). Apartado 6048. Tel: (047) 68 2441. Fax: (5347) 68 2850. Mayabeque. Cuba.
Correo electrónico: dennis.someillan@biocen.cu


{kind=link}
{kind=link}
{kind=link}