










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
En la actualidad los nuevos avances para el estudio de la biología molecular y celular han permitido dilucidar la base estructural, funcional y los mecanismos de regulación de las actividades básicas de la célula.
Se ha realizado una serie de estudio relacionado con un complejo proteico grande presente en todas las células llamado proteasoma, el cual interviene en el procesamiento de los antígenos que son procesados por la vía citosólica y presentados por moléculas del Sistema Principal de Histocompatibilidad (MHC, de sus siglas del inglés) clase -I clásicas y no clásicas.1),2,3
Aunque recientemente, algunos investigadores le han llamado sistema ubiquitina-proteasoma o vía ubiquitina-proteasoma.4,5 Debido a que esta vía juega un rol clave en la proteólisis, ya que es la ruta principal de degradación selectiva de las proteínas en las células eucariontes usando como marcador la ubiquitina para la proteólisis rápida de las proteínas endógenas y exógenas. A su vez, presenta una gran implicación en las múltiples funciones celulares, tales como: el control del ciclo celular, la reparación del ADN dañado y la apoptosis.6-7 Así, este manuscrito tiene como objetivo realizar una revisión bibliográfica del proteasoma, los tipos de proteasomas y sus implicaciones en la tolerancia central de los linfocitos T. Asimismo, se realizó la revisión de los artículos más citados o de interés en la base de datos MedLine.
DESARROLLO
VÍA UBIQUITINA-PROTEASOMA
La vía o sistema ubiquitina-proteasoma fue descubierta en los años 70 de la centuria pasada y ha estado sujeta a diferentes y constantes investigaciones.7 Es el principal sistema de degradación de las proteínas dependientes de adenosín trifosfato (ATP), el cual se encuentra en todas las células.6 Este sistema es esencial en el mantenimiento de la homeostasia proteica celular, la eliminación de proteínas mal plegadas durante su proceso de síntesis o aquellas que se han desnaturalizado.8,9 Así, esta ruta es esencial en la regulación de otros procesos celulares, tales como: la diferenciación celular, la progresión del ciclo celular, el desarrollo y la apoptosis celular; así como en la ontogenia y la respuesta inmune.10
No obstante, para que una proteína sea degradada en el proteasoma debe ser marcada selectivamente con otra proteína llamada ubiquitina, la cual está presente en todas las células eucariontes. Por cual, no solo dio el origen de su nombre al ser ubicua, sino por ser una de las proteínas mejor conservadas entre las especies.
Asimismo, permite que las proteínas que se encuentran en el citosol mal plegadas, exógenas provenientes de microorganismos intracelulares, del proceso de recambio proteico celular (llamado turnover del inglés) o para la obtención de aminoácidos (aa) por medio del reciclaje; sufran el proceso de ubiquitinación (o unión de la ubiquitina) para que sean dirigidos al proteasoma donde serán degradadas en péptidos pequeños.13,14
La ubiquitinación es un mecanismo que implica numerosos factores entre los cuales se incluyen la enzima activadora de la ubiquitina (E1), enzima portadora de ubiquitina o la enzima conjugante de ubiquitina (E2) y la ubiquitina proteína ligasa (E3).15,16
Ubiquitina es un polipéptido o proteína pequeña de 76 aminoácidos, que se encuentra desde el hialoplasma de la levadura hasta en el ser humano, de forma globular, rígida y muy estable con un peso molecular de 8,5 kilodaltons (kDa), que interviene como un marcador selectivo de la degradación de las proteínas.17,18
Aunque, en años recientes en las levaduras se ha observado que no todas las proteínas que están ubiquitinadas son eliminadas, por lo que no se ha podido aclarar con exactitud cómo la maquinaria intracelular puede diferenciar entre diferentes señales de ubiquitinación.19 Por tanto, cabría hacerse la pregunta hasta qué punto tiene un rol regulador en el ciclo celular. Por lo que sería oportuno realizar investigaciones acerca de las implicaciones de los mecanismos epigenéticos sobre los genes que codifican la ubiquitina, E1, E2 y E3; y su repercusión en las funciones biológicas de las mismas en el ciclo celular en células normales o mutadas.
Así, en su extremo carboxilo terminal (C-terminal) presenta dos residuos del aminoácido de glicina (Gly, de sus siglas del inglés) contiguos y el último (Gly76) se une a cadenas laterales del residuo de lisina (Lys, de sus siglas del inglés) mediante un enlace isopeptídico. Además, presenta siete residuos internos de Lys, de los cuales los más importantes ocupan la posición Lys48 y Lys63, aunque el más estudiado es el Lys48.20 Asimismo, al grupo amino (NH2) de esta Lys se le puede unir a una segunda molécula de ubiquitina y a esta, una tercera y así sucesivamente hasta formar una cadena ubiquitinada o poliubiquitinada. Empero, con cuatro polipéptidos de ubiquitinas se establece la señal requerida para que la proteína ubiquitinada entre a la vía de degradación citosólica a través del blanco Lys48, el cual es el sustrato para la degradación proteasomal.
Sin embargo, el proceso de ubiquitinación ocurre en tres pasos mediante tres enzimas, las cuales se mencionaron anteriormente. El primer paso es llevado a cabo cuando la ubiquitina se une a un grupo sulfihidrilo presente en un residuo de cisteína (Cys, de sus siglas del inglés) del centro activo de la E1 con el requerimiento de ATP y Mg2+ formando un enlace tioéster entre el extremo C-terminal por adelinación del residuo de la Gly76 de la ubiquitina. Así, el genoma humano solo codifica para una proteína E1.21,22
El segundo paso, se realiza cuando la ubiquitina se transfiere de la E1 hacia la E2 mediante un enlace tioéster. Esta enzima es más numerosa que la E1, ya que el genoma humano codifica alrededor de 40 proteínas E2.21,22
Así, el tercer paso acontece cuando la E3 se une a la ubiquitina cedida por la E2 a la proteína diana, a través de un residuo de Lys de la misma. Por lo que, la proteína diana queda monoubiquitinada con enlace isopeptídico entre el grupo amino ε del residuo de Lys y el grupo carboxilo α del residuo de Gly. Así, por sucesivos efectos de E1, E2 y E3 sobre el residuo de Lys48 de la última ubiquitina unida a la proteína añaden una cadena de moléculas de ubiquitina en cada ciclo sucesivo.23
La E3 es la enzima del sistema ubiquitina-proteasoma más numerosa ya que se estima que en el ser humano existan alrededor de varias centenas.22 Sin embargo, existen dos tipos principales: la que posee un motivo carboxilo terminal homólogo de la proteína E6 (HECT, de sus siglas del inglés), la cual se encontró por primera vez en la región carboxilo-terminal de la proteína de expresión temprana E6 asociada al virus del papiloma humano.24,25 En la misma se encuentra la Cys conservada que actúa como aceptor de la ubiquitina desde la E2 y se transfiere al sustrato. La otra enzima principal presenta el motivo de nuevo gen realmente interesante (RING, de sus siglas del inglés) en la cual se observa la presencia de Cys e histidinas (His, de sus siglas del inglés) conservadas, que forman una estructura como de "grapa cruzada", las cuales pueden ser simples o complejas.26,27,28
Las simples pueden ser monómeros u homodímeros, mientras que las complejas forman complejos multiproteícos como el complejo promotor del anafase, el SCF y el VBC. Estas enzimas transfieren la ubiquitina directamente desde la E2 hacia el sustrato. Por lo tanto, la E3 es determinante en la especificidad del sustrato, lo cual explicaría su elevado número.21,26,27
Por lo tanto, una vez que una proteína entra a la vía de ubiquitina-proteasoma por ubiquitinación, por lo general es dirigida al proteasoma donde será escindida en pequeños péptidos que serán transportados al retículo endoplásmico donde se podrán unir a moléculas de MHC-I recién sintetizadas. También, estos péptidos pueden sufrir la acción de aminopeptidasas que lo escinden por los enlaces peptídicos dando como resultado aa libres que formarán parte del pool de aa.
PROTEASOMAS
En la década de los 70 del pasado siglo, se realizó el descubrimiento del proteasoma o también llamado proteosoma, el cual es tan grande como un ribosoma.18 Pero actualmente, un sinnúmero de estudios ha permitido determinar que existen varios tipos de proteasomas (Fig.1).2,29,30,31
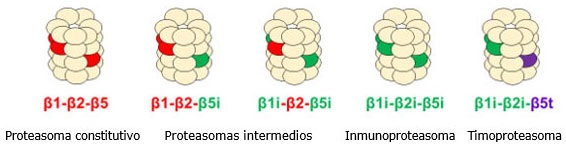
Así, la comprensión de la estructura y funcionamiento de las subunidades del proteasoma ha permitido comprender su asociación con enfermedades cardiovasculares, diabetes, enfermedades neurológicas y cáncer.32,33,34,35,36,37 El gen que codifica el proteosoma se localiza en el cromosoma 6 cerca de la región en donde se codifican las moléculas del MHC-II.38
Proteasoma constitutivo
El proteasoma 26S es llamado el proteasoma constitutivo o clásico por ser una estructura grande macromolecular multiproteica con carácter enzimático que presenta un peso molecular de 2,5 Mega Daltons. Este se encuentra en el núcleo y citoplasma de todas las células eucariotas, pero fundamentalmente en el citosol en los seres humanos con una disposición tubular de unos 50x20 nm.29,39
El proteasoma clásico como se encuentran en el citosol de todas células del cuerpo humano. Así, es el complejo multicatalítico principal que utiliza la célula para la proteólisis. A su vez, es la misma vía que usa la célula para la presentación de péptidos en la superficie en el contexto de moléculas MHC-I clásicas y no clásicas.
El proteasoma 26S está armado por dos complejos el 20S y 19S.40 El complejo 19S con un peso molecular de alrededor de 900 kDa, es llamado la partícula reguladora o subunidad reguladora en la cual se observan dos estructuras: la base y la pestaña.41 La base está formada por seis subunidades con actividad de ATPasa (Rpt1-Rpt6) pertenecientes a la superfamilia AAA+, y tres sin actividad de ATPasa (Rpn1, Rpn2 y Rpn10).42-44
Las ATPasas forman un anillo hexagonal que es común en este tipo de enzimas, las cuales están en contacto con las subunidades α del componente central del complejo 20S. La pestaña con un peso molecular de 400 kDa se asemeja a un disco que puede separarse y unirse al resto de la partícula, la cual está formada por subunidades sin actividad de ATPasa (Rpn3-9,11-12). Las regiones amino-terminal de las subunidades α ocluyen el canal central por lo que funcionan como una compuerta.7,19,45
La partícula central o complejo o proteasoma 20S tiene una disposición cilíndrica con un peso molecular de cerca de los 700 kDa, constituido por cuatro anillos heptaméricos superpuestos que se asemejan a las cuencas de un collar de perlas. Cada anillo comprende siete subunidades proteicas que están formadas por dos anillos β internos (contiene de la subunidad β1-β7) y dos anillos α externos (contiene de la subunidad α1-α7) para un total de 28 subunidades.2, ,29,46
Las proteínas de los anillos α externos son estructurales y carecen de actividad proteolítica. En cambio en los anillos β internos, tres de sus subunidades es donde se lleva a cabo la actividad catalítica de una proteína o polipéptido sustrato una vez que se encuentra en el proteasoma, las cuales son β1 (también llamado como PSMB6, Y y δ), β2 (también conocido como PSMB7, Z y MC14) y β5 (también se le conoce como PSMB5, X, MB1 y ε).47
Estas tres subunidades β1, β2 y β5 han sido identificadas como enzimas tipo caspasas (escindiendo los residuos de aa ácidos), tipo tripsina (las dianas son los aa residuos básicos: arginina o lisina) y tipo quimotripsina (las dianas son los extremos carboxilo terminal de los residuos de aa hidrofóbicos: tirosina, triptófano y fenilalanina), respectivamente.18,47,48 Esta combinación de las subunidades con función proteolítica produce péptidos con una longitud de entre tres y veintidós residuos de aa.49
En la actualidad, se conoce que el proteasoma reconoce los sustratos de tres modos diferentes: el modo dependiente de ubiquitina, el modo mediado por un adaptador y el modo independiente de ubiquitina. Así, en todas las formas en que el proteasoma reconoce el sustrato requiere de ATPasas. Pero, el modo dependiente de ubiquitina es la vía más utilizada por todas las células eucariotas.50,51
El modo mediado por un adaptador fue descrito recientemente por Barthelme y Sauer. Los cuales encontraron que la chaperona citosólica Cdc48 (llamada en los animales p97 o VCP) puede también formar un complejo con la partícula central 20S y contribuir a la degradación del sustrato proteico. Aunque estos mecanismos están muy bien descritos.52,53
El modo independiente de ubiquitina se ha descrito en la degradación de la ornitina descarboxilasa (ODC), pero, se ha observado que otros sustratos pudiesen sufrir la proteólisis en el proteasoma a través de este modo, como: p21/Cip1, c-Jun, c-Fos, p53, p73 IκBα, la cadena α del receptor de células T (TCR, de sus siglas del inglés), Fra-1 y Hif-1a. Más, los mecanismos de este proceso no se comprenden del todo aún.54,55,56,57
Así, el modo dependiente de ubiquitina es la forma más estudiada, ya que una vez que la proteína sustrato sufre el proceso de ubiquitinación puede ser reconocida por el complejo 19S debido a que la Rpn10 se asocia con la poliubiquitina mientras las Rpn1-Rpn2 se unen a la proteína.
Las enzimas desubiquitinizantes separan las ubiquitinas y las subunidades con actividad de ATPasa, utilizando la energía liberada por la hidrólisis del ATP, lo que producen el desplegamiento de la proteína, la cual va pasando hacia la cámara interior del componente central del proteasoma mediante cambios de conformación de las subunidades α que obstruyen la entrada.2,7,19
A medida que la proteína va atravesando la cámara, se produce la hidrólisis de los enlaces peptídicos y las cadenas pequeñas de los péptidos que se forman son liberados a través de la partícula reguladora. Los péptidos formados tienen una vida media muy corta, pues son atacados rápidamente por peptidasas citosólicas permitiendo el reciclaje de aminoácidos o son transportados al retículo endoplásmico.2,7,19
Así, estos péptidos generados pasan al retículo endoplásmico por la acción de un transportador especializado llamado transportador asociado al procesamiento del antígeno (TAP, de sus siglas del inglés), el cual que es miembro de la familia de proteínas transportadoras ABC dependiente de ATP.
Por lo cual, se realiza un transporte activo de forma óptima con péptidos de ocho a dieciséis residuos de aminoácidos que en su extremo presenta grupos carboxilos terminales de aa básicos o hidrófobos, fundamentalmente; hacia la membrana del retículo endoplásmico.46,58,59,60
Una vez en el lumen del mismo, la proteína TAP se asocia a una proteína llamada tapasina que presenta afinidad por moléculas MHC-I recién sintetizadas vacías, las cuales están compuestas por dos cadenas: la microglobulina β2 y la cadena α de la clase I. Así, cada molécula de MHC-I con el péptido antigénico forma un trímero, el cual sufrirá modificaciones postraduccionales tanto el retículo endoplásmico como en el aparato de Golgi. Luego, a través de vesículas de transporte pueden ser expresadas en la superficie celular donde son reconocidas por las células del sistema inmune: linfocitos T CD8+ citotóxicos (CTL, de sus siglas del inglés) y asesinas naturales.46,58,61
Así, estas células son las principales centinelas en el reconocimiento y eliminación de las células que expresen como un marcador celular péptidos originados de la proteólisis llevada a cabo en el proteasoma de proteínas endógenas (mutadas o tumorales) o exógenas (virus, bacterias intracelulares o protozoos).1
Sin embargo, los linfocitos T CD8+ para que puedan ser activados y realizar su función efectora deben reconocer su antígeno cognado en el contexto de MHC-I presentado por la célula dendrítica y, además, debe producirse una cooperación de los linfocitos T CD4+ cooperadores tipo 1 (o Linfocito CD4+ TH1) antes de que maduren a CTL. Una vez activados pueden reconocer los antígenos expresados en las moléculas MHC-I en la superficie de las células dianas.
Inmunoproteasoma
A inicio de la década de 1990, se identificó una forma alternativa de proteasoma, el cual fue observado en células WEHI-3 tratadas con IFNγ. Esta observación originalmente fue referida como un complejo polipéptido de bajo peso molecular (LMP, de sus siglas del inglés), el cual estructuralmente es muy similar al proteasoma clásico. En la actualidad, este complejo es llamado inmunoproteasoma.62
El inmunoproteasoma es expresado en altos niveles de forma constitutiva en las células dendríticas de la médula del timo y en las células epiteliales tímicas medulares (mTEC, de sus siglas del inglés). Además, estas células presentadoras de antígenos presentan la catepsina S que solo es expresada en el timo y en los testículos, la cual es crucial para el procesamiento de la cadena invariante, al córtala en péptidos pequeños por lugares específicos y además, presentando especificidad por el sustrato.63
Asimismo, las mTEC presentan características únicas ya que expresan el factor de transcripción llamado regulador autoinmune (AIRE, de sus siglas del inglés), que les permite la expresión ectópica de un gran número de transcritos que codifican proteínas expresadas habitualmente en la periferia.64 Además, realizan la macroautofagia, lo cual permite que no solo los autopéptidos procesados en los inmunoproteasomas serán presentados en moléculas MHC-I sino también en las moléculas MHC-II.65,66 Este fenómeno de presentación cruzada permitiría educar con autoantígenos a los linfocitos T CD4+ contribuyendo a que se lleve a cabo eficientemente el mecanismo de selección negativa de los mismos. Por lo que, a su vez, contribuye y moldea en el repertorio del TCR de los linfocitos T CD8+ y CD4+.61
Además, el inmunoproteasoma en las células que son y no son de origen hematopoyético es expresado constitutivamente y bajo el efecto del interferón gamma (INFγ), respectivamente. Estas últimas son células blanco del sistema inmune por encontrarse infectadas por microorganismos intracelulares o en tejidos donde está ocurriendo un proceso inflamatorio.67
Así, durante un proceso inflamatorio en el cual se libera citocinas pro-inflamatorias como: el factor de necrosis tumoral alfa (TNFα, de sus siglas del inglés), los interferones tipo II e interferones tipo I en menor medida (INFα e INFβ), las cuales inducen la expresión e incorporación de nuevas subunidades catalíticas alternativas en el proteasoma 20S en las células del sistema inmune.67
Estas subunidades inducidas llamadas β1i (también conocido como PSMB9 y LMP2), β2i (también llamado como PSMB10, LMP10 y MECL-1) y β5i (también conocido por PSMB8 y LMP7) sustituyen las subunidades constitutivas β1, β2 y β5, para formar otra partícula central del complejo 20S.61,68
A pesar de que las subunidades β1i y β5i se encuentran codificadas en la región de las moléculas del MHC-II inmediatamente a corriente arriba de cada uno de los genes de la TAP-1 y TAP2 en el cromosoma 6 (6p21.3), la subunidad β2i se encuentra fuera de esta región.61,62
No obstante, en estudios donde se ha medido la actividad del inmunoproteasoma con péptidos fluorogénicos se ha observado una baja actividad tipo caspasas y una alta actividad tipo tripsina y quimotripsina. Estas diferencias en sus sitios catalíticos en comparación con el proteasoma clásico han permitido determinar que el inmunoproteasoma produce diferentes tipos de péptidos. Esto es debido a que el inmunoproteasoma muestra una tendencia aumentada para escindir detrás de residuos de aminoácidos básicos o hidrofóbicos.49,69
Por lo cual, esto permite predecir que se producirán de forma más eficiente péptidos antigénicos con una alta afinidad para las moléculas MHC-I. Por lo tanto, estos resultados muestran que el inmunoproteasoma juega un rol muy importante en la presentación de antígenos, ya que optimiza la unión de fragmentos peptídicos a moléculas MHC-I.49,67,69,70
Al mismo tiempo, el inmunoproteasoma contribuye a optimizar la respuesta efectora de manera más específica y diversa hacia determinados antígenos realizando una activación más selectiva de clones de linfocitos CD8+, ya que el reconocimiento de estos antígenos está restringido por los alelos de la clase I del MHC propio de cada ser humano.69 Este hecho es llamado fenómeno de restricción por el MHC-I.
Es importante señalar que el estudio del inmunoproteasoma ha permitido comprender los mecanismos de producción de enfermedades autoinmunes, cáncer y cómo el sistema inmune responde antes infecciones por microorganismos intracelulares como virus y bacterias intracelulares, fundamentalmente.46,68
Proteasomas intermedios
Estudios recientes han descrito dos formas alternativas de proteasoma, los cuales son intermedios entre el proteasoma constitutivo y el inmunoproteasoma. Estos proteasomas intermedios se caracterizan por presentar una y dos de las tres subunidades catalíticas del inmunoproteasoma, la β5i y la β1i-β5i, respectivamente.71 Así, estas dos formas de proteasomas intermedios son llamados de igual forma por la presencia de las subunidades de inmunoproteasoma que presentan y además, son simétricos.72
Los proteasomas intermedios β5i y β1i-β5i representan el 10 % y 20 % del total de los proteasomas observados en tumores de los cuales del 30 % al 50 % de ellos se encuentran en el hígado, riñón, intestino delgado, colon y células dendríticas. Asimismo, los proteasomas intermedios β5i y β1i-β5i exhiben actividad tipo tripsina y quimotripsina de forma intermedia entre el proteasoma constitutivo y el inmunoproteasoma. La actividad tipo caspasas del proteasoma intermedio β5i es similar al proteasoma constitutivo, mientras que es baja en el proteasoma intermedio β1i-β5i. Esto es fácilmente explicado por el hecho de que la actividad tipo caspasas esta generalmente asociada con la subunidad β1, la cual está presente en el proteasoma intermedio β5i y no en el proteasoma intermedio β1i-β5i.71
Se ha reportado que las células dendríticas contienen inmunoproteasomas y proteasomas intermedios. Por otro parte, como las células tumorales presentan proteasomas constitutivos y proteasomas intermedios por lo que este hallazgo podría representar una herramienta novedosa en la inmunoterapia del cáncer.49
Timoproteasoma
El timo es un órgano vital en la ontogenia de los linfocitos T, los mismos en esta estructura se les llama timocitos.73 En lo inicial, estos timocitos son doble positivos (CD4+CD8+), los cuales serán sometidos a mecanismos de selección positiva y negativa en la corteza y médula tímica, respectivamente; lo cual finalmente dará como resultado linfocitos T simple positivos CD8+CD4- y CD4+CD8- que saldrán a la periferia con este fenotipo. Sin embargo, primeramente, esto se produce ya que los linfocitos T interactúan con células epiteliales tímicas corticales (cTEC de sus siglas del inglés). Las mismas les presentan una gran variedad de péptidos propios en el contexto de moléculas MHC-I restringido para CD8 y MHC-II restringido para CD4, los cuales son los ligandos para los TCR presente en los linfocitos T. De esta forma, permite que se lleve a cabo el proceso de tolerancia central.74,75
No obstante, para que se realice de forma óptima el mecanismo de selección positiva en los linfocitos CD8+ requiere que las cTEC expresen en su proteasoma la subunidad β5t (también llamado PSMB11), por lo cual se le llama timoproteasoma. Así, el timoproteasoma contiene dos subunidades similares al inmunoproteasoma, la β1i y β2i, pero se diferencia en la subunidad β5t. Empero, cómo el timoproteasoma regula la selección positiva no está del todo claro.31,61
Es sorprendente que los péptidos asociados a las moléculas MHC-I, los cuales fueron procesados por el timoproteasoma son ligandos de baja afinidad cuando son presentados a los TCR en el contexto de CD8+. Por lo cual, estos ligandos son mucho menos inmunogénicos en comparación con los que se enfrentarán posteriormente los linfocitos una vez que salgan a la periferia. Por lo cual, el timoproteasoma genera de forma constitutiva ligandos de péptidos alterados durante el mecanismo de selección positiva, lo cual jugará un rol importante en el proceso de la tolerancia inmune.76,77
Esto se debe, a las diferencias de las propiedades catalíticas de las unidades β5 y β5i en comparación con la unidad β5t, ya que los mismos presentan una mayor afinidad para los residuos de aa hidrofóbicos en sus bolsillos y a su vez, presentan una mejor actividad tipo quimotripsina. Mientras que en el bolsillo de la β5t es más afín para los residuos de aa hidrofílicos por los cual los residuos de aa hidrofóbicos en los péptidos procesados accederán con mayor dificultad en el bolsillo de los mismos.31
Asimismo, este fenómeno permitirá una eficientemente educación en el proceso de ontogenia en la generación de los linfocitos T CD8+ funcionalmente competentes, ya que solo serán seleccionados aquellos clones de linfocitos que sean capaces de reconocer con cierta afinidad los ligandos peptídicos alterados con residuos de aa hidrofílicos. Sin embargo, a los mismos tiempos estos clones tendrán una alta afinidad por péptidos con residuos de aa hidrofóbicos, los cuales a su vez son muy inmunogénicos, ni tampoco fueron educados eficientemente.76,77
Este mismo hecho sucede de cierta forma en el proceso de ontogenia de los futuros linfocitos T CD4+ ya que las cTEC también presentan características únicas como las observadas en las mTEC. Las cTEC presentan una elevada tasa la macroautofagia de forma constitutiva, lo cual permitirá que las moléculas MHC-II carguen autopéptidos procesados en el timoproteasoma o proteínas endógenas las cuales le serán presentadas a los timocitos durante la selección positiva. También, presentan la catepsina L (L en los murinos y catepsina V en los humanos) y la proteasa serin específica del timo (TSSP, de sus siglas del inglés) donde la deficiencia de estas proteasas da como resultado un daño en la selección del repertorio de linfocitos T CD4+.65 Además, esta maquinaria celular presente en los cTEC garantizará que los precursores de los futuros linfocitos T CD4+ sean educados durante el mecanismo de selección positiva con ligandos péptidos alterados con baja afinidad donde su naturaleza son los residuos de aa hidrofílicos. Por los que presentarán una alta afinidad por péptidos con residuos de aa hidrofóbicos presentados en el contexto de moléculas MHC-II.
No obstante, si aún el repertorio de clones de linfocitos T sometidos a este proceso presenta una alta afinidad, los mismos continuarán siendo educados en su proceso de maduración a nivel de la médula tímica por el mecanismo de selección negativa en el timo y por los mecanismos de tolerancia periférica en los órganos linfoides secundarios.
Asimismo, las células en los tejidos periféricos generalmente no expresan hacia al medio extracelular proteínas con residuos de aa hidrofóbicos, solo péptidos o proteínas con residuos de aa hidrofílicos sin señales de peligro. Por lo contrario, las células que expresan péptidos o proteínas con residuos de aa hidrofóbicos son aquellas que se encuentran senescentes, mutadas o infectadas por microorganismos intracelulares. No obstante, se ha observado que en personas con desordenes de inmunoregulación son capaces de desencadenar una respuesta inmune a autoantígenos con una alta inmunodominancia donde su naturaleza peptídica son los residuos de aa hidrofóbicos expuestos en un microambiente inflamatorio o por múltiples factores desde un origen genético hasta ambiental.78,79
Luego, las cTEC solo presentarán autoantígenos a los precursores de los linfocitos T CD8+ y CD4+; y que una vez que culminen su proceso de maduración en el timo y salgan a la periferia no reconocerán o sí serán capaces de reconocer autoantígenos en un contexto de señales de peligro. Mientras que la presentación de antígenos extraños o neo-autoantígenos desencadenará una respuesta efectora por las células de la inmunidad adaptativa con la eliminación de la noxa. Por lo que, las características intrínsecas de las cTEC tanto la presencia del timoproteasoma, la macroautofagia, la catepsina L y TSSP jugarán un rol muy esencial en la educación de los futuros linfocitos T tanto CD4+ y CD8+ en la discriminación de lo propio y no propio.31,75
Por lo que se debería considerar que la tolerancia central no está únicamente definida por el mecanismo de selección negativa, sino que a su vez la selección positiva juega también un papel muy importante en la definición del repertorio de clones de linfocitos T no autorreactivos.
CONSIDERACIONES FINALE
El proteasoma juega un rol muy importante en la homeostasis proteica de la célula, incluyendo la transducción de señales, expresión de genes, ciclo celular, respuesta celular ante el estrés, ontogenia y el proceso de tolerancia central de linfocitos T. Además, está implicado en la respuesta inmune. A su vez, los subtipos de proteasoma permiten realizar un funcionamiento óptimo de la célula según su relación estructura-función. Su estudio no solo permitirá comprender aún más los mecanismos de tolerancia central sino sus implicaciones en las enfermedades oncohematológicas y en los desórdenes autoinmunitarios, brinda nuevas herramientas en el campo de la inmunoterapia.

