










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPastos y Forrajes
versión impresa ISSN 0864-0394
Pastos y Forrajes v.31 n.3 Matanzas jul.-sep. 2008
ARTÍCULO DE INVESTIGACIÓN
Comportamiento ecofisiológico de Brachiaria decumbens en monocultivo y en asociación con Leucaena leucocephala
Ecophysiological performance of Brachiaria decumbens in monocrop and in association with Leucaena leucocephala
A. Rodríguez Petit1, F. Rada2.y M. Colmenares3
1 Universidad Nacional Experimental Sur del Lago. Santa Bárbara de Zulia, Venezuela
E-mail: arodrigu@ula.ve
2 Instituto de Ciencias Ecológicas y Ambientales. Universidad de Los Andes. Venezuela 3 Universidad Nacional Experimental Francisco de Miranda. Venezuela
RESUMEN
Con el objeto de estudiar las relaciones ecofisiológicas que determinan el crecimiento de B. decumbens en monocultivo (MO) y asociada a L. leucocephala (AS), se realizaron evaluaciones de radiación fotosintéticamente activa (RFA), asimilación (A), conductancia estomática (Gs), transpiración (E), humedad relativa (HR), potencial hídrico de la hoja (Øh), diferencial de presión vapor (DPV) e índice de área foliar (IAF) en ambas condiciones (AS y MO), en una zona de bosque húmedo tropical; estas evaluaciones se hicieron a los 5, 10 y 22 días después del pastoreo en ambas condiciones. Los valores más positivos de Øh se encontraron en AS, al igual que los valores más bajos de RFA, con diferencias significativas (P<0,05) para ambos indicadores en las fechas evaluadas; mientras que no se observaron diferencias para las variables microclimáticas HR y DPV. Las variables de intercambio de gases (E, Gs y A) mostraron valores significativamente mayores (P<0,05) en MO en todas las fechas. Los valores más altos (P<0,05) para IAF se obtuvieron en MO a partir de los 10 y 22 días de evaluación. Debido a la limitación en la capacidad fotosintética que ejerce la disminución de la luz sobre la gramínea, se concluye que la RFA es la variable que más determina el crecimiento y desarrollo de B. decumbens en AS. Palabras clave: Brachiaria decumbens, intercambio de gases, sistemas silvopascícolas
ABSTRACT
In order to study the ecophysiological relations that affect the growth of B. decumbens in monocrop (MO) and associated with L. leucocephala (AS), evaluations of photosynthetic active radiation (PAR), assimilation (A) stomatic conductance (Gs), transpiration (E), relative humidity (RH), water potential (Øh), differential of steam pressure (DSP) and leaf area index (LAI) were made in a tropical humid forest zone. The evaluations were made 5, 10 and 22 days after grazing under both conditions (MO and AS). The most positive values of Øh (P<0,05) were found in AS, as well as the lowest values of PAR (P<0,05) in all the evaluated dates. RH and DSP did not show differences (P 0,05). The gas exchange parameters (E, Gs and A) showed significantly higher values in MO (P<0,05) in all dates. LAI was higher in MO after 10 and 22 days of evaluation. Due to the limitation of the photosynthetic capacity exerted by the light decrease on the grass, that PAR is concluded to be the most important factor that determine growth and development of B. decumbens in AS.
Key words: Silvopastoral systems, gas exchange, B. decumbens
INTRODUCCIÓN
En el trópico de Latinoamérica el desarrollo de los sistemas de pastoreo se ha sustentado casi en su totalidad en la adopción de esquemas tecnológicos basados en la explotación de especies de gramíneas en monocultivo, sin tener en cuenta las consecuencias negativas que este tipo de agricultura produce en el medio ambiente y el ecosistema en general, además de ser sistemas poco estables y de baja sostenibilidad (Iglesias, 1998; Sánchez, 1998). Los problemas que ha generado esta forma de crecimiento agrícola son múltiples, pero su impacto está relacionado principalmente con la limitación de la biodiversidad, la deforestación, la erosión (Detweiler y Hall, 1988; Houghton, Skole y Lefkowitz, 1991; Fearnside y Barbosa, 1998) y la promoción de sistemas de cultivo altamente dependientes de la utilización de agroquímicos. Por otro lado, estos sistemas no han demostrado una alta eficiencia en el trópico, donde la productividad de los rumiantes por superficie es baja y alrededor del 50% de las áreas de pastoreo se encuentran degradadas (Pezo e Ibrahim, 1999), lo que contribuye al aumento del CO2 atmosférico (Hougthon et al., 1991). Los cambios necesarios pueden estar representados por las transformaciones en los modelos tradicionales o por mejoras en la eficiencia de técnicas de uso común.
El uso de los árboles y los arbustos forrajeros (asociaciones, cercas vivas, bancos de forraje, etc.) en los sistemas silvopastoriles (SSP), puede constituir la respuesta de los sistemas de explotación agropecuarios tropicales a los retos de la protección del ambiente. Su introducción ha tomado importancia debido a que, desde el punto de vista productivo, ofrecen forraje con un alto contenido de proteína y minerales esenciales, además de una alta digestibilidad (Rodríguez Petit, Clavero y Razz, 2001); adicionalmente ayudan a la regeneración natural de las pasturas (Oliveira, Oliveira, Ferreira, Alves, Miranda, Vilela, Urquiaga y Boddey, 1999), son hábitat de diversas especies y favorecen la actividad de la fauna del suelo (Sánchez y Milera, 2002; Sileshi y Mafongoya, 2005). Aunque desde el punto de vista agronómico han sido ampliamente estudiados, es escaso el conocimiento acerca de la ecología de estos sistemas asociados, que involucran factores que les confieren a las gramíneas una condición de crecimiento diferente a la del monocultivo. Esta situación se debe a que las gramíneas forrajeras tropicales son predominantemente de ruta fotosintética C4, mientras que las leguminosas son C3, lo que determina tasas de crecimiento diferentes que complican el manejo, ya que bajo este supuesto las gramíneas, al poseer mayor capacidad para aprovechar la radiación solar y fijar CO2, tienen una recuperación posterior al pastoreo más rápida que las leguminosas, con lo cual alcanzan su óptimo forrajero mucho antes que la especie arbórea. Sin embargo, en las asociaciones la reducción de la intensidad de la luz bajo la copa de los árboles forrajeros es ecológicamente importante, no solo por su efecto directo en la asimilación, sino porque afecta también otros factores microclimáticos como la humedad relativa, la humedad del suelo y la temperatura, los cuales, en conjunto, influyen en la respuesta ecofisiológica de la gramínea asociada. El objetivo del presente trabajo fue estudiar las principales relaciones ecofisiológicas en el comportamiento de Brachiaria decumbens en monocultivo y en asociación con Leucaena leucocephala, con el fin de comparar los factores que más influían en su desarrollo en ambas condiciones. MATERIALES Y MÉTODOS Área de estudio El ensayo se realizó en el año 2005 durante los meses de mayo a julio, después del primer pico de precipitación del año que ocurre en abril, en la Hacienda Judibana, propiedad de la Universidad de Los Andes al norte del Estado Mérida, Venezuela, a 68 msnm y ubicada geográficamente entre las coordenadas 8° 37' 10'', 8° 37' 26'' latitud Norte y 71° 41' 00'', 71° 42' 22'' longitud Oeste. Las condiciones climáticas ubican a la zona como Bosque Húmedo Tropical según la clasificación de Holdridge (Ewel y Madriz, 1968), con promedios anuales de precipitación de 1 990 mm, temperatura de 26,8ºC, humedad relativa de 80,74% y 2 776 mm de evaporación. Se compararon dos condiciones de pastoreo: el monocultivo (MO) de B. decumbens (Gramineae, Poaceae), y la asociación (AS) en sistema silvopastoril de esta gramínea y L. leucocephala (Leguminoseae, Mimosoideae) con una arquitectura de siembra en hileras, distancia entre estas de 6 m y árboles continuos dentro de la hilera. Ambas condiciones se desarrollan próximas una a la otra, se encontraban completamente establecidas y en plena producción desde hace unos 15 años, y eran manejadas bajo pastoreo rotativo según el caso. Las mediciones se hicieron después del pastoreo, y coincidió el inicio de estas el mismo día en ambas condiciones. Variables evaluadas Índice de área foliar. En cada condición en estudio (AS y MO) se determinó el índice de área foliar (IAF), mediante el medidor portátil de área foliar LAI-2000 Plant Canopy Analizer (LiCor®). Se realizaron tres mediciones de IAF en cada condición, a los 5, 22 y 30 días después del pastoreo. Intercambio de gases. Se tomaron registros de radiación fotosintéticamente activa (RFA), asimilación (A), conductancia estomática (Gs) y transpiración (E), con el uso del equipo portátil de intercambio de gases LCA-4 (ADC®); se hicieron 11 repeticiones de la medición en cada fecha y condición en estudio. Relaciones hídricas. Se registró la humedad relativa (HR) mediante un medidor infrarrojo de humedad y la temperatura del aire y de la hoja mediante termocuplas, a fin de estimar los cambios en el diferencial de presión vapor (DPV). También se determinó el potencial hídrico de la hoja (Øh) mediante el método de la bomba de presión de Scholander. Se hicieron nueve repeticiones de la medición en cada fecha y condición evaluada. Las mediciones para intercambio de gases y relaciones hídricas se realizaron a los 5, 10 y 22 días después del pastoreo en ambas condiciones. En el análisis de los resultados se utilizó el paquete estadístico SAS (Statistical Analysis System, 1990). Para el análisis de varianza se utilizó el procedimiento PROC GLM, con un modelo de bloques completamente al azar. Las medias se compararon con la prueba de mínima diferencia significativa con el procedimiento LS MEANS. Las pruebas de correlación de Pearson se realizaron con el procedimiento PROC CORR. RESULTADOS Y DISCUSIÓN La tabla 1 muestra los datos registrados para las variables microclimáticas medidas en ambas condiciones durante el ensayo. En la HR y el DPV no existieron diferencias significativas en las fechas evaluadas para ambas condiciones. Sin embargo, en el DPV se observó en el monocultivo una ligera tendencia a registrar valores más altos, lo que pudo deberse a una mayor demanda de agua en esta condición entre la hoja y el ambiente, relacionada con las altas temperaturas. En términos generales las plantas en ambas condiciones presentaron valores de Øh que están alejados del punto de pérdida de turgor para estas especies, lo cual es explicable debido a las condiciones subhúmedas de la zona. Guenni, Gil, Baruch, Márquez y Núñez (2006) encontraron que los síntomas de marchitez en las especies del género Brachiaria (incluida B. decumbens) se presentan con valores de Øh de 2.0 MPa; mientras que en el presente estudio los valores más bajos estuvieron cercanos a 1.0 MPa. 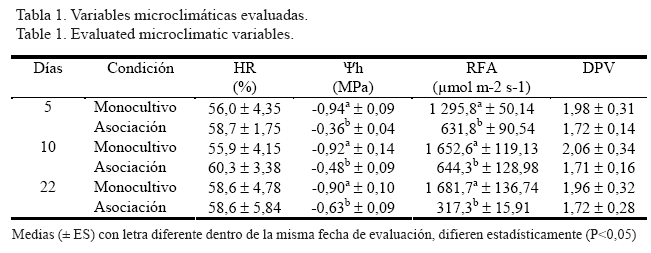
Sin embargo, los resultados mostraron que, aun bajo estas condiciones, las plantas que se encontraban en la AS registraron valores de Øh significativamente mayores (P<0,05) a los observados en las plantas de MO. Se ha sugerido que la protección bajo la copa de los árboles provoca una disminución en la pérdida de agua de las plantas (Baruch y Fisher, 1991) y, por otro lado, esta protección también disminuye la temperatura del suelo, por lo que la pérdida de agua por evaporación es menor (Wilson y Wild, 1991), de modo que el complejo suelo - planta presenta más disponibilidad de humedad. Existió diferencia significativa (P<0,05) en todas las fechas para la RFA entre las condiciones evaluadas. En AS se observó una disminución progresiva de la RFA a lo largo de las mediciones, en comparación con los valores registrados en MO. En términos porcentuales esta disminución de la luz incidente fue de 51,3% a los cinco días; 61,1% a los 10 días y 81,2% a los 22 días de evaluación. Este comportamiento se debió a que la luz incidente fue inicialmente interceptada por la planta arbórea (L. leucocephala), y esta intercepción aumentó progresivamente a medida que la planta recuperó su biomasa aérea después de la defoliación. En plantaciones de cualquier leñosa perenne, la transmisión de luz hacia el estrato herbáceo tiende a declinar con el tiempo; el grado de esta disminución responde principalmente a la relación entre la edad y la morfología de la leñosa (Wilson y Ludlow, 1991). Los valores significativamente más altos de E, Gs y A se registraron en MO (tabla 2). Los estomas desempeñan un papel determinante tanto en la difusión del CO2 como en la transpiración. Su funcionamiento está regulado por la fotosíntesis, en función de cambios en la presión parcial interna de CO2, y por la luz disponible; por otro lado, también está regulado por la diferencia entre el Øh y el del aire (ÄØh hojaaire). Como se muestra en la tabla 1 los valores más bajos de Øh se registraron en MO, lo cual podría ejercer una señal de cierre estomático en función de evitar la pérdida de agua (E). Sin embargo, como se observa en la tabla 2 la mayor Gs se registró para MO, lo que parece estar asociado a la necesidad de una alta difusión de CO2 al aparato fotosintético de la planta, debido a que los valores más altos de A igualmente se hallaron en esta condición. En AS se observaron los valores significativamente más bajos de Gs y E (P<0,05). En condiciones hídricas adecuadas (bajo DPV y adecuado Øh en la planta), las plantas de sombra pueden experimentar conductancias estomáticas que permiten una adecuada suplencia de CO2 al mesofilo, lo cual aumenta la capacidad de las gramíneas C4 asociadas de responder a la luz difusa y a los haces o flecos de luz (sunflecks). Lambers, Chapin y Pons (1998) sugieren que las plantas de sombra en condiciones hídricas favorables, son capaces de mantener mejores ganancias de carbono que cuando presentan algún grado de estrés hídrico. El comportamiento descrito para Gs y E, en MO y AS, estuvo en concordancia con la asimilación de CO2 (A) en ambas condiciones. La A más alta (P<0,05) se registró en MO en todas las fechas evaluadas y el mayor valor se encontró a los cinco días, lo cual puede coincidir con un comportamiento compensatorio posterior a la defoliación. Por su parte, los valores de A en AS disminuyeron progresivamente en la medida que trascurrió el período de descanso, hasta casi la mitad, de los cinco a los 22 días. En MO la adecuada suplencia de luz estimuló las altas tasas de A. Estas plantas presentaron una Gs más alta, por lo que fueron capaces de absorber suficiente CO2 como para mantener una adecuada ganancia de carbono. En AS las plantas no tenían acceso a una adecuada suplencia de RFA, lo que limitó su respuesta fotosintética (baja A); por lo tanto, mantener Gs bajas fue suficiente para permitir el suministro adecuado de CO2 a su aparato fotosintético. 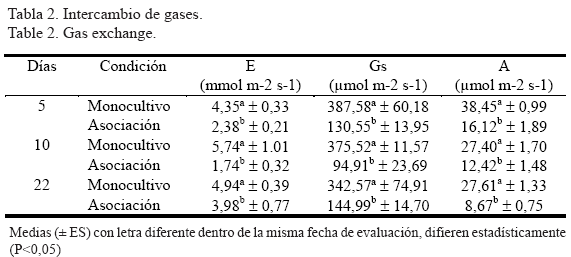
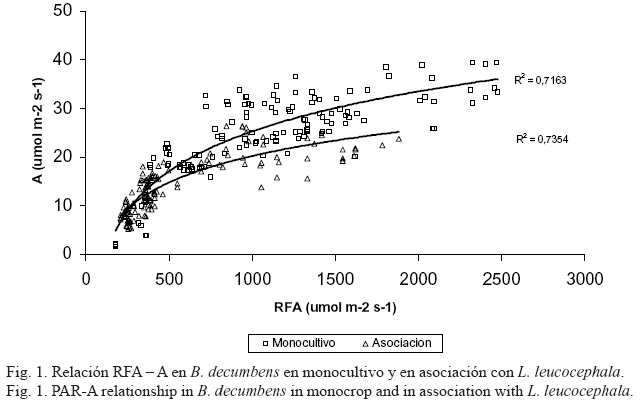
El análisis de correlación practicado a la relación RFA-A muestra que existe dependencia de A con RFA, tanto en MO como en AS (r= 0,893, p<0,001 y r=AS: 0,752, p<0,001; para MO y AS, respectivamente). Las curvas RFA-A que se muestran en la figura 1, describen una fuerte relación exponencial de la luz sobre la asimilación.
A iguales tasas de RFA las plantas de MO presentaron una mayor capacidad asimilatoria que las de AS, situación descrita anteriormente por Horne y Blair (1991) y que se atribuye a los cambios estructurales y bioquímicos de la planta para adaptarse a las condiciones de luz limitada. Las plantas de sombra poseen menos cloroplastos, menos componentes de la cadena transportadora de electrones, menos Rubisco (Ribulosa 1,5-bifosfato carboxilasa/oxigenasa) y menos nitrógeno por unidad de área. Esto, en conjunto con otros factores morfológicos y fisiológicos, reduce la capacidad de carboxilación y de transporte de electrones, lo que limita la capacidad fotosintética de estas plantas (Lambers et al., 1998). Los valores registrados para el IAF se muestran en la tabla 3. A los cinco días de pastoreo no había una recuperación de la biomasa defoliada y no se encontró diferencia entre los valores para AS y MO. A los 22 días se halló diferencia significativa (P<0,05) entra ambas condiciones, situación que se mantuvo a los 30 días después del pastoreo. Podría esperarse a los 30 días que el efecto acumulativo de la biomasa recuperada por el estrato arbóreo y el herbáceo en la AS comenzara a igualar los valores de IAF observados en MO en la misma fecha; sin embargo, aparentemente a los 30 días la participación de la biomasa de L. leucocephala en la biomasa total en AS aun era baja, lo cual puede ser explicado por su condición de C3 que le confiere una menor tasa de crecimiento que la de la gramínea. 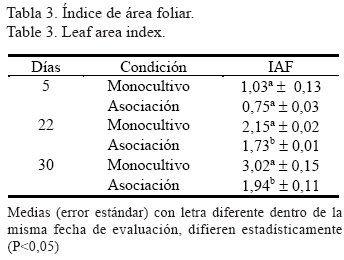
La respuesta en el IAF estuvo en concordancia con lo que ocurrió en RFA y A, lo cual se manifestó en una producción de biomasa más rápida en MO que en AS. En AS una parte de la reducción del IAF es consecuencia de la baja intensidad de luz sobre la fotosíntesis neta; sin embargo, debe considerarse que las alteraciones en el aparato fotosintético, que incluyen la declinación de la actividad de la cadena transportadora de electrones y de las enzimas carboxiladoras, pueden ser también una causa importante de esta disminución (Lin et al., 1999). Pezo e Ibrahim (1999) plantearon que la respuesta a la sombra es diferente entre las especies de pastos tropicales (C4) y existen algunas que pueden tolerar o no el sombreamiento. Dicha tolerancia se expresa principalmente en la disminución de la tasa de crecimiento en una misma especie en condiciones de sombra. Estos autores describen a B. decumbens como tolerante a niveles medios de sombreamiento. Norton, Wilson, Shelton y Hill (1991) sugieren que estas especies resultan de gran importancia para su inclusión en sistemas agroforestales (silvopastoriles). Conclusiones y Recomendaciones De acuerdo con los resultados se puede concluir que las condiciones de luz en el monocultivo favorecieron una mejor respuesta de B. decumbens en términos de fotosíntesis y que se manifestaron en un mejor crecimiento, hasta las edades evaluadas. En la asociación la respuesta de la planta estuvo fuertemente determinada por la RFA que pasa a través de la leguminosa arbórea. Aunque no fue objetivo del trabajo hacer evaluaciones de calidad del forraje en estos sistemas, es conveniente obtener dicha información, ya que se ha reportado variaciones de la calidad por efecto de la sombra, principalmente en lo que se refiere a proteína y digestibilidad (Pezo e Ibrahim, 1999). Esto constituye una información importante desde el punto de vista del uso forrajero. Agradecimientos Los autores desean expresar su agradecimiento al personal de Hda. Judibana de la Universidad de Los Andes por ceder el uso de sus instalaciones y su colaboración en el trabajo de campo. Igualmente, el apoyo del Instituto de Ciencias Ambientales y Ecológicas (ICAE) de la Universidad de Los Andes por facilitar los equipos y la logística en esta investigación. REFERENCIAS BIBLIOGRÁFICAS 1. Baruch, Z. & Fisher, M. 1991. Factores climáticos y de competencia que afectan el crecimiento de la planta en el establecimiento de pasturas. En: Establecimiento y renovación de pasturas: conceptos, experiencias y enfoques de investigación. (Eds. C. Lascano y J. Spain). 6ta. Reunión Comité Asesor RIEPT. Centro Internacional de Agricultura Tropical (CIAT). Cali, Colombia 2. Detweiler, R. & Hall, C. 1988. Tropical forest and the global carbon cycle. Science. 239:42 3. Durr, P. & Rangel, J. 2000. The response of Panicum maximum to simulated subcanopy environment. 1. Soil x shade interaction. Tropical Grasslands.34:110 4. Ewel, J. & Madriz, A. 1968. Zonas de vida de Venezuela. Dirección de Investigación. Ministerio de Agricultura y Cría. Venezuela. p. 65 5. Fearnside, P. & Barbosa, R. 1998. Soil carbon changes from conversion of forest to pastures in Brazilian Amazonia. Forest Ecology and Management. 80:35 6. Guenni, O.; Gil, J.; Baruch, Z.; Márquez, L. & Nuñez, C. 2006. Respuesta al déficit hídrico en especies forrajeras de Brachiaria (Trin.) Griceb. (Poaceae). Interciencia. 31:505 7. Horne, P. & Blair, G. 1991. Forage tree legumes: Productivity of leucaena/grass mixtures. Australian J. Agric. Res. 42:1231 8. Sánchez, M. 1998. Sistemas agroforestales para intensificar de manera sostenible la producción animal en América Latina. Revista Pecuaria de Nicaragua. 2 (19):12 9. Sánchez, Saray & Milera, Milagros. 2002. Dinámica de la macrofauna edáfica en la sucesión de un sistema de manejo de gramíneas a un sistema con árboles intercalados en el pasto. Pastos y Forrajes. 25:189 10. Sileshi, G. & Mafongoya, P. 2005. Variation in macrofaunal communities under contrasting land use systems in eastern Zambia. Applied Soil Ecology. 33:49 11. Statistical Analysis System (SAS). 1990. SAS/STAT User's guide Version 6. 4th edition. SAS Institute Inc. Cary, NC. 12. Wilson, J. & Ludlow, M. 1991. The environment and potential growth of herbage under plantations. In: Forages for plantation crops. (Eds. M. Shelton and W. Stür). ACIAR Proceedings No. 32. Canberra, Australia. p. 10 13. Wilson, J. & Wild, D. 1991. Improvement of nitrogen nutrition and grass growth under shading. In: Forages for plantation crops. (Eds. M. Shelton and W. Stür). ACIAR Proceedings No. 32. Canberra, Australia. p. 77 Recibido el 20 de julio del 2007
Aceptado el 17 de abril del 2008

{kind=link}