










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPastos y Forrajes
versión impresa ISSN 0864-0394
Pastos y Forrajes v.31 n.3 Matanzas jul.-sep. 2008
ARTÍCULO DE INVESTIGACIÓN
Selección de rizobios adaptados a ecosistemas ganaderos de Sancti Spíritus, Cuba, inoculados en maíz (Zea mays, L.). Fase I: Invernadero
Selection of rhizobia adapted to livestock production ecosystems of Sancti Spiritus, Cuba, inoculated in corn (Zea mays L.). Stage I: Greenhouse
C.J. Bécquer1, Beatriz Salas1, *D. Archambault2, J. Slaski2 y A. Anyia2
1 Estación Experimental de Pastos y Forrajes Sancti Spíritus, Cuba
E-mail: becquer@pastos.yayabo.inf.cu
2 Department Environmental Technologies, Alberta Research Council, Vegreville, AB, Canada
E-mail: slaski@arc.ab.ca
* Dirección actual: Laurentian University, 935 Ramsey Lake Road, Sudbury, ON
E-mail:DArchambault@laurentian.ca
RESUMEN
Se efectuó un ensayo en invernadero con el objetivo de seleccionar cepas de rizobio inoculadas en maíz (Zea mays, L.). Para ello se utilizaron 30 cepas nativas cubanas pertenecientes a Bradyrhizobium sp., así como 12 cepas comerciales, pertenecientes a diferentes géneros y especies de rizobio. Los inóculos se confeccionaron en medio CLM y fueron agitados hasta alcanzar un título de 106-108 cél./mL. Se utilizaron los métodos estándar para la inoculación de cereales. Se evaluaron las variables: peso seco aéreo (PSA), peso seco raíz (PSR), longitud del tallo (LT), diámetro del tallo (DT) y presencia de mazorcas (PM). En las variables PSA, LT y DT se hallaron tratamientos superiores al control absoluto, y en el caso de la LT, dos tratamientos presentaron valores sin diferencias significativas a los del control fertilizado. No se detectaron diferencias estadísticas entre tratamientos en la PM. En el PSR los tratamientos inoculados fueron inferiores al control fertilizado, excepto JI2, TD1, SP7 y SP12. Se obtuvo una agrupación en conglomerados, mediante la cual se seleccionaron las cepas TD, JK, SP, SP, TE, JK, SP, USDA 110, JI, JJ, USDA 76, SP, 25B6, SP, NITRAGIN, ATCC 10317, que se destacaron para su posterior evaluación en condiciones de campo. Los tratamientos seleccionados en el dendrograma constituyeron el 69% para las cepas nativas y el 31% para las comerciales. Al evaluar los tratamientos inoculados en cuanto al grado de dependencia de las diferentes variables con respecto al PSR, se determinó que el índice de regresión fue débil, pero al analizarlos por separado, se observó una regresión fuerte en las variables DT (cepas nativas) y PSA (cepas comerciales). Se recomienda la continuación de estas investigaciones en condiciones de campo. Palabras clave: Bradyrhizobium, inoculación, Zea mays
ABSTRACT
A greenhouse trial was carried out with the objective of selecting rhizobia strains inoculated in corn (Zea mays, L.). Thirty Cuban native strains belonging to Bradyrhizobium sp. were used, as well as 12 commercial strains belonging to different rhizobium genera and species. The inoculi were made in BYM medium and were shaken until reaching a titer of 106-108 cel./mL. The standard methods for cereal inoculation were used. The variables evaluated were: aerial dry weight (ADW), root dry weight (RDW), stem length (SL), stem diameter (SD) and presence of ears (PE). In the variables ADW, SL and SD treatments higher than the absolute control were found, and in the case of SL, two treatments showed values without significant differences with the fertilized control. No statistical differences were found among treatments in PE. In RDW the inoculated treatments were lower than the fertilized control, except JI2, TD1, SP7 and SP12. A grouping in clusters was obtained, by means of which the strains TD, JK, SP, SP, TE, JK, SP, USDA 110, JI, JJ, USDA 76, SP20, 25B6, NITRAGIN, ATCC 10317, were selected, which stood out to be evaluated later under field conditions. The treatments selected in the dendrogram constituted 69% for the native strains and 31% for the commercial ones. When evaluating the inoculated treatments regarding the degree of dependence of the different variables with regards to RDW, the regression index was determined to be weak, but when analyzing them separately, a strong regression was observed in the variables SD (native strains) and ADW (commercial strains). To continue these studies under field conditions is recommended.
Key words: Bradyrhizobium, inoculation, Zea mays
INTRODUCCIÓN
Muchos autores señalan que los rizobios (rizobacterias conocidas comúnmente como bacterias dinitrofijadoras en simbiosis con las leguminosas) pueden colonizar e influir en el crecimiento de las plantas no leguminosas (Antoun, Beauchamp, Goussard, Chabot y Lalande, 1998). Las rizobacterias colonizan de forma agresiva las raíces de las plantas. Aquellas que reciben el nombre de bacterias promotoras del crecimiento vegetal (PGPR, en Inglés) son una pequeña porción (2-5%) de las rizobacterias que promueven el crecimiento de las plantas (Antoun y Kloepper, 2001). Este proceso estimulante puede ser beneficioso, por lo tanto, para aquellas plantas que tradicionalmente han sido sometidas a altas dosis de fertilizantes inorgánicos por su efecto esquilmante en el suelo. Entre éstas, se encuentran los cereales. Existen criterios conservadores, como el de Cocking (2004), quien considera que en la mayoría de los casos, la bacteria coloniza sólo la superficie de la raíz de los cereales y queda vulnerable a la competencia de otros microorganismos. No obstante, Bécquer, Salas, Archambault, Slaski y Anyia (2006) han demostrado los beneficios de fertilizantes inorgánicos por su efecto esquilmante en el suelo. Entre éstas, se encuentran los cereales. Existen criterios conservadores, como el de Cocking (2004), quien considera que en la mayoría de los casos, la bacteria coloniza sólo la superficie de la raíz de los cereales y queda vulnerable a la competencia de otros microorganismos. No obstante, Bécquer, Salas, Archambault, Slaski y Anyia (2006) han demostrado los beneficios de la inoculación de plantas de trigo con cepas nativas de Bradyrhizobium sp., procedentes de ecosistemas ganaderos de Sancti Spiritus, Cuba. El objetivo de este trabajo fue estudiar el efecto de la inoculación de rizobios nativos cubanos y comerciales en maíz, y seleccionar las cepas más promisorias para su posterior evaluación en condiciones de campo.
MATERIALES Y MÉTODOS
Identificación de las cepas de rizobio. Se utilizaron 30 cepas procedentes de leguminosas naturalizadas de Sancti Spíritus, Cuba (Centrosema, Neonotonia y Stylosanthes). Estas cepas fueron confirmadas en trabajos anteriores como rizobios y ubicadas taxonómicamente en el género Bradyrhizobium sp. (Bécquer, 2003). Se utilizaron también 12 cepas comerciales pertenecientes a diferentes géneros y especies de rizobio (tabla 1).
Preparación de los inóculos. Los inóculos se confeccionaron en medio CLM (caldo-levadura-manitol) (Somasegaran y Hoben, 1994), a partir de una asada de cultivos frescos en medio agarizado. Se colocaron en agitación (120 rpm) durante siete días (para las especies de lento crecimiento) o de tres a cuatro días (especies de rápido crecimiento), en una zaranda orbital a 30oC, hasta alcanzar un título UFC (unidad formadora de colonias) de 106-108 cél./mL Ensayos de invernadero. Se escogió la variedad de maíz 2338 F15, donada por el Alberta Research Council, Canadá. Las semillas fueron pregerminadas previamente a su siembra en macetas. Cada maceta contenía 1,0 kg de suelo estándar mezclado con bajo nivel de N: 30 ppm (mg/kg). Se ralearon las macetas a los cuatro días de la siembra hasta dejar dos plantas por maceta. Se utilizaron los métodos estándar para la inoculación de los cereales (Sabry, Saleh, Batchelor, Jones, Jotham, Webster, Kothari, Davey y Cocking, 1997; Biswas, Ladha y Dazzo, 2000), con cambios en la frecuencia y la cantidad de inóculo. El inóculo bacteriano en cantidad de 3 mL/planta, que contenía 106-108 cél./mL, se aplicó a los cinco días de la siembra y, consecutivamente, se realizaron otras tres inoculaciones con el mismo título (6 mL/planta, 12 mL/planta, y 12 mL/planta, respectivamente), a los 20, 30 y 40 días de la siembra, con un total de cuatro inoculaciones por maceta. Se evaluaron las plantas a los 90 días de la siembra, excepto la presencia de mazorcas (PM), a los 60 días de la siembra. El resto de las variables medidas fueron: peso seco aéreo (PSA), peso seco raíz (PSR), longitud del tallo (LT) y diámetro del tallo (DT) datos correspondientes a PM fueron transformados por el coeficiente de Barlett, según Lerch (1976). Diseño experimental y análisis estadístico. Se empleó un diseño en bloques al azar con 44 tratamientos y cuatro réplicas. Se incluyó un control absoluto y uno con un portador nitrogenado (NH4NO3, 150 kg N/ha). Se realizó un ANOVA mediante el paquete estadístico StatGraphics Plus, versión 2.0, 1994-1996, Statistical Graphics Corporation. Las diferencias entre medias se hallaron mediante la prueba de LSD de Fisher y se tomó en consideración la desviación estándar (DE). Se construyó un dendrograma mediante el método de Ward (distancia euclidiana). RESULTADOS Y DISCUSIÓN
En el peso seco aéreo se constataron cuatro tratamientos inoculados con cepas nativas: JI2 (12,75 g/maceta), JK1 (12,44 g/maceta), SP7 (12,69 g/maceta) y SP21 (13,43 g/maceta) que fueron estadísticamente superiores (P<0,05) al control absoluto (tabla 2), lo cual representó un incremento del 25,5% (JI2), 22,4% (JK1), 27,0% (SP7) y 32,2% (SP21). Estos valores fueron superiores a los encontrados por Prévost, Saddiki y Antoun (2000) en un experimento donde se inoculó maíz con cepas comerciales de Bradyrhizobium japonicum, en el cual se obtuvieron incrementos en el PSA de la planta desde un 6,7% hasta 8,7% en comparación con el control absoluto. También fueron superiores a los datos obtenidos por Antoun y Prévost (2000), con la cepa P31 de Rhizobium leguminosarum, al ser inoculada en maíz. Por otra parte, en este experimento los tratamientos inoculados con cepas comerciales presentaron superíndices comunes con respecto al control absoluto, en cuanto a PSA, PSR y NM. Estos resultados pudieran deberse a una mayor secreción total de sustancias promotoras del crecimiento vegetal por las cepas nativas; o a la secreción por éstas de determinadas fitohormonas mayormente dirigidas al desarrollo de partes aéreas de la planta, con un efecto superior en comparación al de las cepas comerciales. Según Biswas et al. (2000) los microorganismos diazotrofos pueden promover el crecimiento vegetal mediante la transferencia del N2 fijado, o mediante el mejoramiento de la absorción de nutrientes a través de la modulación de actividades hormonales en las plantas inoculadas. Otra posibilidad es en cuanto al mecanismo de entrada de los rizobios en las raíces de los cereales, ya que se conoce que dicho mecanismo es diferente al realizado en la simbiosis leguminosa rizobio (Reddy, Ladha, So,Hernández, Ramos, Ángeles, Dazzo y de Bruijn, 1997), el cual depende principalmente de la capacidad de la bacteria (cepas) para romper las paredes de la célula vegetal, y no de reacciones bioquímicas basadas en dependencias genéticas específicas.
Aunque Lupwayi y Clayton (2004) plantearon que los rizobios pueden actuar como promotores del crecimiento vegetal y cambian la fisiología y morfología de las raíces inoculadas, en este experimento en la variable peso seco raíz ningún tratamiento presentó diferencias significativas con el control absoluto y todos fueron estadísticamente inferiores al control fertilizado (3,80 g/maceta). Algunos tratamientos, como los inoculados con JI2 (3,15 g/maceta), TD1 (3,04 g/ maceta), SP7 (3,11 g/maceta) y SP12 (3,00 g/maceta), mostraron una tendencia estadística al incremento de sus valores, por presentar superíndices superiores con respecto al control absoluto. También Bécquer et al. (2006), al inocular trigo en condiciones de invernadero, hallaron 27 tratamientos, inoculados indistintamente con cepas nativas y comerciales, cuyos valores fueron estadísticamente iguales a los del control absoluto en el peso de la raíz. Estos resultados apuntan necesariamente a una escasa secreción, por parte de las cepas, de fitohormonas específicas al desarrollo radicular, aunque no se descartan otras variantes. En la longitud del tallo, excepto los tratamientos inoculados con TE1 (53,93 cm), TE4 (55,63 cm), HA3 (61,63 cm), HG2 (61,88 cm), JH1 (61,88 cm), JJ4 (60,13 cm), JK3 (61,50 cm), SP1 (60,88 cm), SP8 (60,88 cm), SP16 (61,13 cm), SP18 (61,00 cm), SP23 (60,25 cm) y ATCC9930 (61,50 cm), los demás resultaron estadísticamente superiores (P<0,05) al control absoluto y dos de éstos, sin diferencias significativas con el control fertilizado (TD1: 74,68 cm y JK1: 73,38 cm).
Esto indica una fuerte dependencia del crecimiento de la planta con relación al desarrollo radicular, quizás basada en el mecanismo de emisión de fitohormonas específicas por parte de estas cepas. Sin embargo, al inocular trigo con cepas cubanas, Bécquer et al. (2006) encontraron que los tratamientos inoculados con las cepas mencionadas anteriormente (excepto HG2, JJ4 y JK3) presentaron superíndices comunes con el control absoluto. Resultados similares se encontraron en maíz, al ser inoculado con cepas canadienses (Bécquer, Salas, Archambault, Slaski y Anyia, 2008) en condiciones de invernadero.
Al parecer, estos mecanismos pudieran variar de acuerdo con el tipo de cultivo que se inocula, aunque deben efectuarse otras evaluaciones para determinar las verdaderas causas. En la variable diámetro del tallo sólo cinco tratamientos inoculados con cepas nativas fueron estadísticamente superiores (P<0,05) al control absoluto (TD1: 10,60 cm, TD4: 10,70 cm, JI2: 11,05 cm, SP7: 10,55 cm y SP18: 10,45 cm) y ninguno superior al control fertilizado. Según Yanni (2001), los beneficios de la asociación rizobio cereal conllevan una mayor producción de la biomasa vegetativa y reproductiva, y están mayormente relacionados con la modulación de la arquitectura radicular de la planta, provocada por el rizobio para una mejor adquisición de algunos nutrientes del suelo, más que por la fijación biológica del dinitrógeno. En cuanto a la presencia de mazorcas a los 60 días de la siembra, todos los tratamientos presentaron letras comunes con el control fertilizado y el control absoluto, por lo que esta variable no fue lo suficientemente discriminante como para seleccionar las cepas por su efecto fisiológico en la planta.
En el dendrograma (fig. 1) se muestran dos grandes conglomerados: el 1, compuesto por el control fertilizado y 16 tratamientos, que constituyen aquellas cepas que mostraron valores estadísticos superiores en una o más variables (TD1, JK6, SP12, SP7, TE4, JK1, SP21, USDA 110, JI2, JJ6, USDA 76, SP20, 25B6, SP15, NITRAGIN, ATCC 10317) (tabla 2). Por otra parte, el con glomerado 2 incluyó un total de 26 tratamientos y el control absoluto, agrupados de forma independiente, donde el 2.1.1 lo integraron el control absoluto y el tratamiento TE.

Este tratamiento presentó los valores estadísticamente inferiores con respecto al control fertilizado en las distintas variables, excepto en la presencia de mazorcas, y no difirió del control absoluto en todas las variables (tabla 2). Los tratamientos seleccionados positivamente en el dendrograma constituyeron el 69% para las cepas nativas y el 31% para las comerciales.
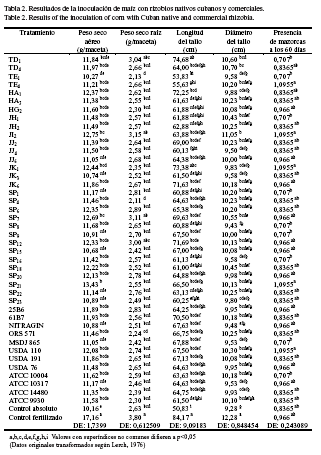
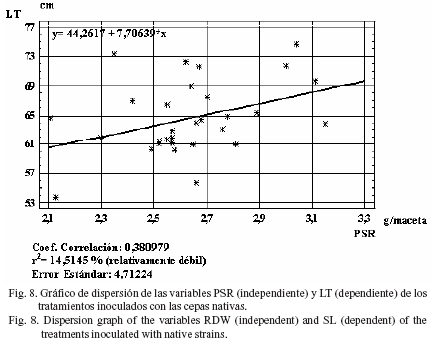
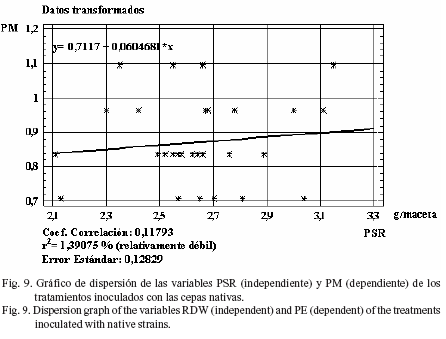
Al evaluar todos los tratamientos inoculados con cepas nativas o comerciales, indistintamente, en cuanto al grado de dependencia entre variables, se determinó que el índice de regresión fue relativamente débil en todos los casos analizados: r2 = 21,7% para la variable dependiente PSA y la independiente PSR (fig. 2), así como r2= 9,6% para LT y PSR (fig. 3), r2 = 21,3% para DT y PSR (fig. 4) y r2 = 4,3% para PM y PSR (fig. 5).
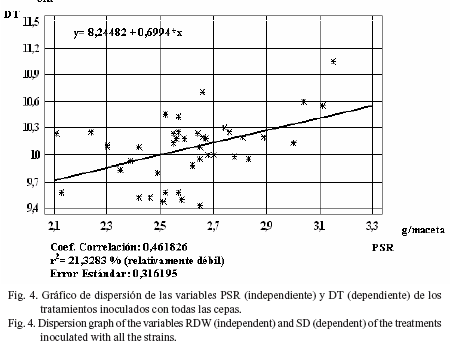
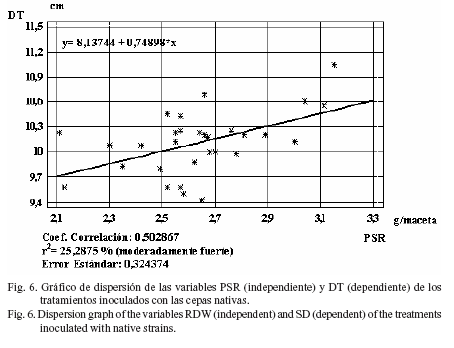
Sin embargo, al separar los análisis por el origen de las cepas, se observó que entre las cepas nativas estos índices variaron, al presentar el DT con respecto al PSR una regresión moderadamente fuerte (r2 = 25,29%) (fig. 6), aunque el índice de regresión fue relativamente débil (r2= 20,00%) en el PSA con respecto al PSR (fig. 7).
Estos resultados coinciden con la regresión encontrada en estas mismas variables, pero en plantas de trigo, al ser inoculadas con las cepas anteriormente señaladas en condiciones de invernadero (Bécquer et al., 2006). El índice de regresión en LT con respecto a PSR fue relativamente débil (r2 = 14,5%), así como el correspondiente a PM con relación a PSR (fig. 8 y fig. 9, respectivamente).
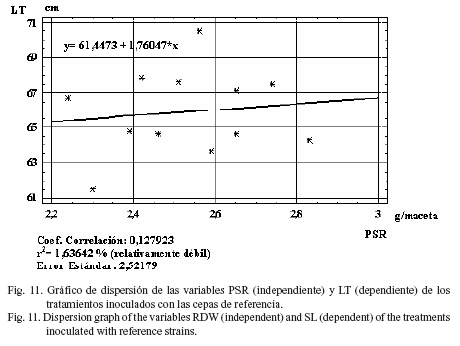
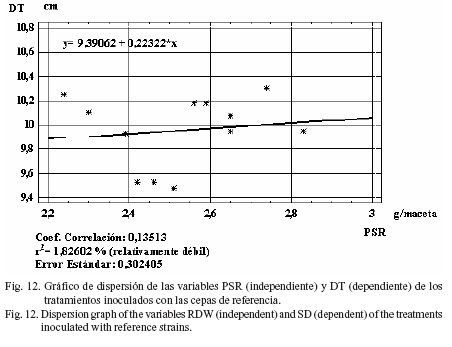
También se halló un índice moderadamente fuerte (r2= 30,5%) entre la variable PSA con respecto a PSR (fig. 10) en el análisis de regresión para las cepas comerciales, y débil (r2= 1,6%) para LT (fig. 11), DT (r2= 1,8%) (fig. 12) y PM (r2= 22,0%) (fig. 13).
Estos resultados indican una débil dependencia de estas variables con respecto al desarrollo radicular, excepto en el diámetro del tallo (cepas nativas) y en el peso seco aéreo (cepas comerciales), donde se corroboró lo planteado por Lupwayi y Clayton (2004) acerca de que los rizobios pueden actuar como promotores del crecimiento vegetal y cambiar la fisiología y morfología de las raíces inoculadas, lo que resulta en una mayor absorción de N y otros nutrientes.
Es posible que al no evaluar algunos de los indicadores agroproductivos de la planta, no se haya podido determinar una relación o dependencia más definitoria entre las variables; aunque este aspecto, en opinión de los autores, no le resta calidad a la selección de las cepas. Se concluye que en las variables PSA, LT y DT existieron tratamientos que fueron estadísticamente superiores al control absoluto, y en el caso de la LT dos tratamientos presentaron valores sin diferencias significativas a los del control fertilizado. En el PSR los tratamientos inoculados fueron inferiores al control fertilizado, excepto JI2, TD1, SP7 y SP12.
No se detectaron diferencias estadísticas entre tratamientos en la PM. Se obtuvo una agrupación, en conglomerados, de los tratamientos inoculados que mostraron valores estadísticos superiores o inferiores, de acuerdo con las distintas variables, por lo que se efectuó una selección de aquellas cepas (TD1, JK6, SP12, SP7, TE4, JK1, SP21, USDA 110, JI2, JJ6, USDA 76, SP20, 25B6, SP15, NITRAGIN, ATCC 10317) que se destacaron, para su posterior evaluación en condiciones de campo. Al evaluar todos los tratamientos en cuanto al grado de dependencia de las diferentes variables con respecto al PSR se determinó que, en general, el índice de regresión fue débil, pero al analizarlos por separado se observó una regresión fuerte en las variables DT (cepas nativas) y PSA (cepas comerciales).
Se recomienda la continuación de estas investigaciones en condiciones de campo. Agradecimientos Al igual que en experimentos anteriores realizados con trigo, el financiamiento, así como las instalaciones, equipos y materiales utilizados para este trabajo, fueron provistos por Alberta Research Council (Vegreville, Alberta, Canadá), a través de un proyecto de colaboración con la Estación Experimental de Pastos y Forrajes Sancti Spíritus, Cuba. Los autores agradecen profundamente a todos los funcionarios de esta Institución canadiense, especialmente al Dr. Daniel Archambault y al Dr. Jan Slaski, por su gran esfuerzo en hacer realidad este proyecto. REFERENCIAS BIBLIOGRÁFICAS 1. Antoun, H.; Beauchamp, C.J.; Goussard, N.; Chabot, R. & Lalande, R. 1998. Potential of Rhizobium and Bradyrhizobium species as plant growth promoting rhizobacteria on nonlegumes: Effect on radishes (Rhaphanus sativus L.). Plant and Soil. 204:57 2. Antoun, H. & Kloepper, J.W. 2001. Plant growth promoting rhizobacteria. Encyclopedia of Genetics. (Brenner, S. and Miller, J.F., Eds.). Academic Press, New York. p. 1477 3. Antoun, H. & Prévost, D. 2000. PGPR activity of Rhizobium with nonleguminous plants. Proceedings of the 5th International PGPR workshop. Villa Carlos Paz, Córdoba, Argentina. p .62 4. Bécquer, C.J. 2003. Caracterización y selección de rizobios adaptados a ecosistemas ganaderos de Sancti Spíritus, Cuba. Tesis en opción al grado científico de Doctor en Ciencias Biológicas. Universidad de La Habana. 140 p. 5. Bécquer, C.J.; Salas, Beatriz; Archambault, D.; Slaski, J. & Anyia, A. 2006. Inoculación de trigo (Triticum aestivum, L.) con rizobios adaptados a ecosistemas ganaderos de Sancti Spiritus, Cuba. Pastos y Forrajes. 29:255 6. Bécquer, C.J.; Salas, Beatriz; Archambault, D.; Slaski, J. & Anyia, A. 2008. Selección de rizobios adapta dos a ecosistemas ganaderos de Alberta, Canadá; inoculados en maíz (Zea mays, L.). Fase I: invernadero. Logro Científico del Instituto de Investigaciones de Pastos y Forrajes. La Habana, Cuba 7. Biswas, J.C.; Ladha, J.K. & Dazzo, F.B. 2000. Rhizobia inoculation improves nutrient uptake and growth of lowland rice. Soil Sc. Soc. of America J. 64:1644
8. Cocking, E.C. 2004. Endophytic colonization of plant roots by nitrogen fixing bacteria. Plant and Soil. 252:169 9. Lerch, G. 1976. La experimentación en las ciencias biológicas y agrícolas. Ed. Ciencia y Técnica. La Habana, Cuba. 452 p. 10. Lupwayi, N.Z. & Clayton, G.W. 2004. Endophytic rhizobia and other diazotrophic bacteria in nonlegume crops. Recent Res. Devel. Crop Sci. 1:227 11. Prévost, D.; Saddiki, S. & Antoun, H. 2000. Growth and mineral nutrition of corn inoculated with effective strains of Bradyrhizobium japonicum. Proceedings of the 5th International PGPR workshop. Villa Carlos Paz, Córdoba, Argentina. 7 p. 12. Reddy, P.M.; Ladha, J.K.; So, R.B.; Hernández, R.J.;Ramos, M.C.; Ángeles, O.R.; Dazzo, F.B. & de Bruijn, F.J. 1997. Rhizobial communication with rice roots: Induction of phenotypic changes, mode of invasion and extent of colonization. Plant and Soil. 194:81
13. Sabry, S.R.S.; Saleh, S.A.; Batchelor, Caroline; Jones, J.; Jotham, J.; Webster, G.; Kothari, S.L.; Davey, M.R. & Cocking, E.C. 1997. Endophytic establishment of Azorhizobium caulinodans in wheat. Proc. R. Soc. Lond. B. 264:341 Somasegaran, P. & Hoben, H.J. 1994. Handbook for rhizobia. Springer-Verlag, New York. 450 p.
14. Yanni, Y.G. et al. 2001. The beneficial plant growth promoting association of Rhizobium leguminosarum bv. trifolii with rice roots. Australian Journal of Plant Physiology. 28 (9):845 15. Bécquer, C.J.; Salas, Beatriz; Archambault, D.; Slaski, J. & Anyia, A. 2008. Selección de rizobios adapta dos a ecosistemas ganaderos de Alberta, Canadá; inoculados en maíz (Zea mays, L.). Fase I: invernadero. Logro Científico del Instituto de Investigaciones de Pastos y Forrajes. La Habana, Cuba
Recibido el 28 de junio del 2007
Aceptado el 4 de marzo del 2008

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}