










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPastos y Forrajes
versión impresa ISSN 0864-0394
Pastos y Forrajes v.32 n.4 Matanzas oct.-dic. 2009
ARTÍCULO DE INVESTIGACIÓN
Cinética ruminal y crecimiento de cabritos suplementados con un probiótico de bacterias ácido-lácticas
Ruminal kinetics and growth of kids supplemented with a lactic acid bacteria probiotic
M.A. Galina1, M. Delgado-Pertiñez2, M.A. Ortíz-Rubio1, L.J. Pineda 3y D.C. Puga4 1Facultad de Estudios Superiores Cuautitlán-Universidad Nacional Autónoma de México, México
E-mail: miguelgalina@correo.unam.mx
2 EUITA-Universidad de Sevilla, España
3 Facultad de Medicina Veterinaria y Zootecnia, Universidad de Colima, México.
4Instituto Nacional de Ciencias Médicas y Nutrición «Salvador Zubirán», México
RESUMEN
Con el objetivo de evaluar la cinética ruminal y el crecimiento de cabritos suplementados con un probiótico de bacterias ácido-lácticas, se asignaron 86 animales Alpinos en un experimento durante 120 días. En el tratamiento uno (T1) 43 cabritos recibieron una dieta constituida por 50% de heno de alfalfa, 40% de concentrado comercial y 10% de alimento nitrogenado de lento consumo (DI). Los animales del tratamiento dos (T2) recibieron DI rociada diariamente con 50 mL de probiótico (BAL) por kilogramo de MS. Se determinó la desaparición de MS in situ, el consumo voluntario de MS, la degradación de la fibra, la concentración de NH3 y AGV, la digestibilidad in vivo, el pH ruminal, los derivados de purinas y la ganancia de peso. La ganancia de peso diaria fue de 129 y 169 g para T1 y T2, respectivamente (P<0,05). El NH3 y la digestibilidad del N y de la fibra fueron mayores para T2 (P<0,05). El tiempo medio de desaparición de la hemicelulosa fue mayor (P<0,05) en T2. Los conteos totales BAL fueron 1,6 y 2,5 millones de ufc/mL en T1 los días 1 y 7. Los cabritos del T2 presentaron 2,4 y 12,5 millones de ufc/mL. Se concluye que la adición de BAL a la dieta para cabritos en crecimiento puede incrementar la ganancia de peso vivo, así como se producen cambios favorables en la digestibilidad, la proteína microbiana y la cinética ruminal.
Palabras clave: Bacterias acidolácticas, cabritos, probióticos
ABSTRACT
With the objective of evaluating the ruminal kinetics and growth of kids supplemented with a lactic acid bacteria (LAB) probiotic, 86 Alpine animals were assigned to a trial for 120 days. In treatment 1 (T1), 43 kids received a diet constituted by 50% alfalfa hay, 40% commercial concentrate and 10% slow intake nitrogen feed (ID). The animals in treatment two (T2) received ID daily sprayed with 50 mL probiotic (LAB) per kilogram of DM. In situ DM disappearance, voluntary DM intake, fiber degradation, NH3 and VFA concentration, in vivo digestibility, ruminal pH, purine derivatives and weight gain were determined. The daily weight gain was 129 and 169 g for T1 and T2, respectively (P<0,05). NH3 and N and fiber digestibility were higher for T2 (P<0,05). The half-time disappearance of hemicellulose was higher (P<0,05) for T2. The total LAB counts were 1,6 and 2,5 million fcu/mL in T1 on days 1 and 7. The kids in T2 showed 2,4 and 12,5 million fcu/mL. It is concluded that the addition of LAB to the diet for growing kids can increase live weight gain, as well as produce favorable changes in digestibility, microbial protein and ruminal kinetics.
Key words: Lactic acid bacteria, goat kids, probiotics
INTRODUCCIÓN
Los agricultores orgánicos, los medioambientalistas y la comunidad científica preocupada por el bienestar animal, enfrentan desafíos en la prevención y el control de enfermedades en los animales domésticos y en el fortalecimiento de la producción, debido a la prohibición del uso de medicamentos y aditivos químicos (Patra et al., 2009). En este sentido, los probióticos constituyen una alternativa (Ortiz et al., 2009).
Los probióticos contienen levaduras, bacterias ácido-lácticas, hongos, cultivo de Bacillus subtillis, estreptococos y/o la mezcla de los anteriores (Nocek et al., 2002; 2003). Las bacterias ácido-lácticas, que incluyen el género Lactobacilli, son las bacterias probióticas administradas con mayor frecuencia (Brashears et al., 2003); estas son residentes normales del tracto gastrointestinal y a menudo se consideran sustitutos naturales de los antibióticos alimentarios (Reid y Friendship, 2002).
Los animales suplementados con probióticos de bacterias lácticas (BAL) muestran un incremento en la eficiencia de la utilización del alimento y la resistencia a las enfermedades (Ortíz et al., 2009). Recientemente, los probióticos y los prebióticos han recibido atención, por su papel en el control de las enfermedades infecciosas y el mejoramiento de la productividad en bovinos lecheros (Galina et al., 2009). Generalmente las bacterias productoras de ácido láctico han probado su eficiencia como probióticos y se han utilizado como promotores del crecimiento, para prevenir infecciones intestinales, disminuir el estrés, estimular la respuesta inmune y aumentar la producción de leche (Couret et al., 2004; Galina et al., 2009).
Las bacterias benefactoras, como las bifido-bacterias, los lactobacilli y algunas especies de enteroccocci, proveen de nutrientes a las células intestinales, lo que estimula la absorción de nutrientes, crea un ambiente intestinal saludable y promueve un vigoroso sistema inmune (Czarnecki-Maulden, 2008).
Con mayor especificidad, se reportó que los simbióticos (combinaciones de prebióticos y probióticos), mezclados en el alimento, mejoran la producción de leche en vacas Holstein. Además inhiben el crecimiento de salmonella y previenen la diarrea, mientras aumentan la ganancia de peso en becerros (Yasuda et al., 2007).
Se ha discutido que el uso de probióticos lácticos en los rumiantes aumenta las concentraciones de amoniaco en el rumen, incrementa la digestibilidad del nitrógeno y de la fibra, además de favorecer mayores conteos de lactobacilos, hasta 12,5 millones de ufc/mL de contenido ruminal, en comparación con la cantidad normal de 1,5 millones/mL (Ortiz et al., 2009); también se registran cantidades superiores de purinas. El mejor comportamiento de los animales alimentados con probióticos puede explicarse por una disminución de la metanogénesis, lo que conserva la energía para el desarrollo animal (Ortiz et al., 2009). La producción de proteína microbiana a través de bacterias probióticas es una tecnología desarrollada para incrementar la cantidad y la calidad de la proteína y la energía enviadas al intestino delgado (Ortiz et al., 2009).
El objetivo del presente estudio fue evaluar la cinética ruminal y el crecimiento de cabritos suplementados con un probiótico de bacterias ácido-lácticas.
MATERIALES Y MÉTODOS
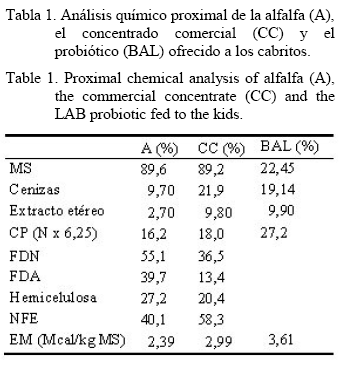
El presente trabajo se realizó en Querétaro, México. Durante 120 días se utilizaron 86 cabritos de raza Alpina, de 20,4 (± 0,5) kg de peso corporal (PC), sin distinción de sexo, distribuidos en un diseño completamente al azar, en dos tratamientos: T1): 43 cabritos (20,7 ± 0,7 kg PC) alimentados con una dieta integral (DI) de 50% de heno de alfalfa, 40% de concentrado y 10% de un suplemento nitrogenado de lento consumo (SNLC) (Galina et al., 2007) y T2): 43 cabritos (20,9 ± 0,4 kg PC) alimentados con la DI rociada diariamente con 50 mL/kg de MS de probiótico (BAL), formado por una mezcla de suero de quesería, melaza, urea, sulfato amónico y un cultivo de bacterias lácticas (Lactobacillus plantarum, L. helveticus, L. delbrueckii, Lactoccocus lactis, L. cremoris, Leuconostoc mesenteroides, Bifidus esscencis y Saccharomyces cerevisiae) diluidas en agua. Se realizaron los análisis químicos de los alimentos de acuerdo con la metodología del AOAC (1995), cuyos resultados se muestran en la tabla 1.
Los animales se alojaron en corrales separados y permanecieron en estabulación durante el período experimental (120 días), con agua a libre acceso; el alimento se ofreció dos veces por día, a las 8:00 y a las 14:00 h., ajustando la oferta para asegurar un 15% de rechazo. Los cabritos se pesaron mensualmente; el consumo de alimento se midió tres veces a la semana, pesando el alimento ofrecido con relación al rechazado.
Se utilizaron cuatro cabras fistuladas para el estudio de la cinética ruminal, bajo un diseño de cuadrado latino (4 x 2), a las cuales se les ofrecieron los dos tratamientos en evaluación. Cada animal tuvo 15 días de adaptación a la dieta y cinco días para la toma de muestras.
La degradabilidad in situ se realizó mediante la técnica de bolsa de nailon (Ørskov et al., 1980); se utilizaron bolsas de poliéster, de 7 x 15 cm, con 1 600 poros/cm2, de la marca Ankom, con un tamaño de poro de 3 mm. Dentro de las bolsas, se colocaron 3 g de muestra de rastrojo de maíz como forraje de referencia para los estudios de digestibilidad (Galina et al., 2004). Las bolsas fueron colocadas dentro del rumen a intervalos de 8, 16, 24, 48, 72 y 96 horas. Al final del tiempo de incubación, el material fue retirado del rumen para ser lavado durante períodos de 10 minutos en un agitador mecánico, hasta que el fluido fue transparente; posteriormente se secó a 65°C durante 48 horas. Al material residual se le determinó el contenido de MS y las fracciones de la fibra. Las bolsas correspondientes a la hora cero, únicamente fueron lavadas para determinar la cantidad de material soluble en la muestra. Para interpretar los datos se utilizó la ecuación sugerida por McDonald (1981) y Dhanoa (1988), la cual surge de la ecuación: Pt = a + b (1-ect) propuesta por Ørskov y McDonald (1979), donde:
Pt = igual que Yt
Yt =Wo + Bd (1-e-c (t-tL))
Yt =Pérdida en tiempo `t', horas
Wo=residuos del lavado a tiempo 0; el tiempo cero incluye las pérdidas por el lavado de material soluble y partículas pequeñas.
Bd =es el material insoluble pero potencialmente degradable
e = exponencial
c= es la tasa fraccionada constante del exponencial. El exponencial es una escala de b, por lo tanto b y c juntos definen la degradación del forraje
t = Tiempo
Digestibilidad in vivo. Se recolectaron las heces durante un período de cinco días, tomando un alícuota de 25% para su procesamiento. Las heces se secaron en una estufa de aire forzado a 70ºC por 36 h, y se almacenaron en botellas herméticas hasta su análisis. Se determinó la MS, el N, la MO, la FND, la celulosa y la hemicelulosa. Además se recolectó la excreción total de orina durante cinco días, agregando ácido sulfúrico (10%) para mantener el pH por debajo de 3; después se tomaron muestras de 100 mL y se congelaron a -20ºC hasta los análisis. Se midió la velocidad de pasaje con la fibra Cr-mordante, dada a través de la aplicación del Cr-mordante en el alimento y la recuperada en las heces, en un período de tiempo de 24 y 48 h.
Determinaciones de pH, NH3 y AGV. Se extrajeron muestras de fluido ruminal (50 mL) a las 0, 2, 4, 6, 8, 12, 16 y 22 horas, mediante una cánula en el rumen, las cuales se filtraron a través de dos capas de gasa y se mantuvieron en un termo previamente gaseado con CO2. En el laboratorio se transfirieron a botellas más pequeñas mientras se gasificaban con CO2. Estas fueron almacenadas en una incubadora a 39ºC; el fluido ruminal fue diluido (106-108) y se inocularon e incubaron 0,5 mL de cada una de las tres disoluciones; después se contaron las colonias según la técnica descrita por Hungate (1969). Se utilizó una submuestra de este fluido para medir el pH con un potenciómetro portátil; mientras que el NH3 se determinó en forma directa, ya que en solución acuosa el amoníaco se encuentra en su forma ionizada (NH4+), a través de un electrodo específico para este ión (marca Orion). La medición se llevó a cabo a diferentes intervalos: 0, 2, 4, 6, 8, 10 y progresivamente hasta las 96 horas del primer día del período experimental; se filtró a través de tres paños de gasa para obtener una muestra libre de partículas sólidas; para evitar la pérdida del compuesto se adicionó de tres a cuatro gotas de HCl a 0,2 N por cada 10 mL de líquido filtrado. Los AGV totales se determinaron mediante la cromatografía de gases, así como sus proporciones molares.
Análisis químico. La determinación de las fracciones de fibra se realizó de acuerdo con la metodología de Goering y VanSoest (1970), y la MS, la MO y el N, por la AOAC (1995). El valor de energía bruta (Mcal/kg) de las dietas y las muestras fecales se determinó con un calorímetro de bomba adiabática.
Análisis matemático. Los resultados de la cinética ruminal se analizaron mediante un análisis de varianza y las diferencias entre medias fueron evaluadas con la prueba de Tukey (P<0,05); mientras que se utilizó el test de Student para comparar las diferencias de las cabras canuladas de las dietas T1 y T2, con el programa SAS (1996).
RESULTADOS Y DISCUSIÓN
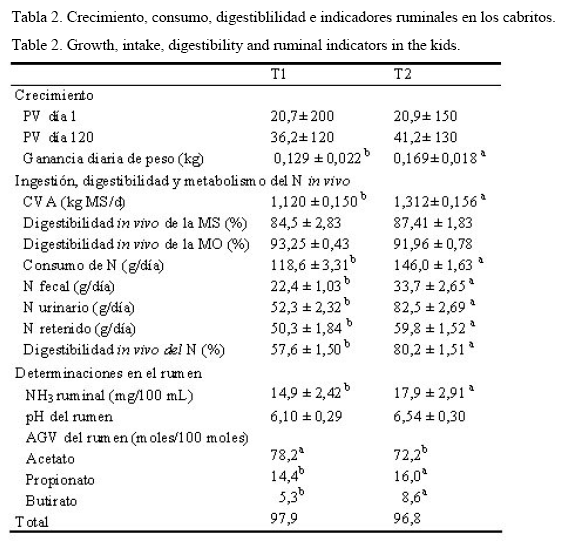
Los cabritos suplementados con probiótico (T2) mostraron un mejor crecimiento (P<0,05), manifestado en un mayor peso corporal (169 g en comparación con 129 g/d para los cabritos control en la dieta T1 (tabla 2). Estos resultados están en concordancia con la investigación realizada en cabras maltesas en crecimiento que recibieron Lactobacilli, las cuales tuvieron mayores ganancias de peso corporal que el control (Chiofalo et al., 2004). Los incrementos en la tasa de degradabilidad, en la proteína microbiana y en la digestibilidad de la fibra pueden explicar los resultados. Además, el consumo de MS y N fue mayor en T2 en comparación con T1 (P<0,05).
El pH ruminal (tabla 2) no mostró diferencias estadísticas entre los tratamientos. No obstante, la cantidad de concentrado (45%) en la dieta T1 pudiera explicar la disminución del pH, debido a la formación de AGV cuyos valores fueron de 6,2; mientras que la utilización efectiva del ácido láctico pudo influir en la moderación de la acidez en el T2.
La estabilización del pH se asocia generalmente con la disminución de los niveles de ácido láctico en el rumen. A pesar de que se aumentó la concentración de los Lactobacilli, los cuales incrementan la concentración de ácido láctico ruminal, al parecer este se utilizó como sustrato para el crecimiento de bacterias ruminales que transforman el ácido láctico en propiónico, con la correspondiente moderación del pH ruminal (Galina et al., 2009).
El tratamiento con probióticos BAL de la dieta T2 tarjo consigo mayores concentraciones de amoniaco ruminal (tabla 2) en comparación cn T1.
Satter y Slyter (1974) sugirieron una concentración de 3-5 mg de NH3/100 mL de líquido ruminal como óptima para aumentar el crecimiento de los microorganismos ruminales; los resultados del presente trabajo cubren fácilmente los requerimientos de concentración de amoniaco sugeridos (14 vs 17 mg/100 mL) en T1 y T2, respectivamente.
La digestibilidad in vivo de la MS y la MO no difirió entre las dietas (tabla 2); en sentido general la digestibilidad de la alfalfa y de los concentrados es alta, debido a los bajos contenidos de la pared celular.
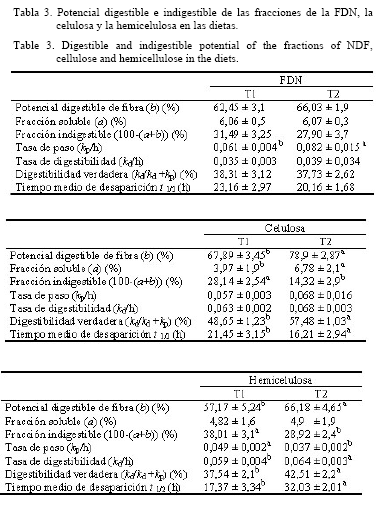
La velocidad de pasaje de la FND (Kp/h) fue mayor (P<0,05) en T2 (0,061/h y 0,082/h para el T1 y T2, respectivamente). La digestibilidad verdadera de la FND fue superior (P<0,05) en T2 (47,73%) con respecto al T1 (38,31%) (tabla 3). En el tratamiento con probióticos las mayores poblaciones microbianas de bacterias ácido-lácticas pudieron incrementar la utilización de la fibra (con el aumento de la velocidad de pasaje) y el consumo de MS, lo cual se manifiestó en un mayor crecimiento de los cabritos.
Nocek et al. (2002; 2003) demostraron que el tratamiento con probióticos (lactobacilos y cultivo en levadura) incrementa el número de bacterias celulolíticas en el rumen y, en algunos casos, aumenta la degradación de la celulosa. En este sentido, la tasa de digestión para la hemicelulosa fue mayor en T2 que en T1 (P<0,05), mientras que las diferencias en la velocidad de digestión para la celulosa no fueron significativas (tabla 3). Sin embargo, la digestibilidad verdadera de la celulosa y la hemicelulosa fue mayor en T2 con respecto a T1 (P<0,05). La desaparición en tiempo medio (t ½) para la hemicelulosa fue mayor (P<0,05) en T2 (32,03 h) con relación a T1 (17,37 h), pero no para la celulosa (tabla 3).
Newbold et al. (1995) sugirieron que A. oryzae y S. cerevisiae estimularon la velocidad de la digestión de la fibra por los microorganismos ruminales.
El incremento del uso de la celulosa y la hemicelulosa probablemente se debió a una mayor degradación de las paredes celulares, por la explosión de la población microbiana ruminal y la acción de las bacterias lácticas. Sin embargo, se necesitan estudios más profundos sobre el efecto específico de las bacterias lácticas en la digestión de la fibra en cabras.
La excreción de derivados de las purinas (DP) en la orina (µmol/W0,75) fue afectada (P<0,05) por la adición de los probióticos lácticos a la dieta T2 (tabla 4). Aunque no se ha encontrado información disponible sobre el efecto de las bacterias lácticas en la excreción de DP en cabritos, ello pudiera explicar el incremento de estas, lo cual se manifestó en una explosión de la proteína microbiana digerible en el intestino.
La proteína bacteriana puede ser una gran porción de la proteína total necesitada por un rumiante cuando mejora la fermentación microbiana (Galina et al., 2007; Galina et al., 2009), como se demostró anteriormente.
Por otra parte, Jouany et al. (1998) no reportaron diferencias en la excreción de DP en ovinos suplementados con S. cerevisiae y Aspergillus oryzae. Las diferencias en las especies de microorganismos utilizadas en las mezclas probióticas y prebióticas (sustancias que favorecieron la acción de los probióticos) y sus diversas actividades, pudieran explicar los resultados variables (Ortíz et al., 2009).
Las bacterias L. plantarum, L. helveticus, L. delbrueckii, L. lactis, L. cremoris y L. mesenteroides fueron aisladas del rumen de las cabras canuladas, después del tratamiento con probióticos y en el control. Los conteos totales medios fueron de 1,6 (T1) y 2,4 (T2) millones de ufc/mL en el primer día y aumentaron a 2,5 (T1) y 12,5 (T2) millones de ufc/mL en el séptimo día, con predominio de Lactobacilli (P<0,05). Estos resultados son similares a los de Dawson et al. (1990), quienes encontraron un aumento significativo en los conteos de bacterias ruminales en novillos que consumieron una dieta de forraje suplementada con un probiótico vivo.
CONCLUSIONES
La adición de BAL a la dieta para cabritos en crecimiento puede incrementar la ganancia de peso vivo, así como se producen cambios favorables en la digestibilidad, la proteína microbiana y la cinética ruminal.
AGRADECIMIENTOS
La investigación fue financiada por PAPPIT IN200809 y la cátedra CONS-207.
REFERENCIAS BIBLIOGRÁFICAS
1. AOAC. 1995. Official Methods of Analysis. 16th ed., Association of Official Analytical Chemists, Washington DC, USA. p. 600
2. Brashears, M.M. et al. 2003. Isolation, selection, and characterization of lactic acid bacteria for a competitive exclusion product to reduce shedding of Escherichia coli O157:H7 in cattle. J. Food Prot. 66:355
3. Chiofalo, V. et al. 2004. Effects of the administration of Lactobacilli on body growth and on the metabolic profile in growing Maltese goat kids. Reprod. Nutr. Dev. 44:449
4. Couret, V. et al. 2004. Number and strains of lactobacilli in some probiotic products. International Journal of Microbiology. 97:147
5. Czarnecki-Maulden, G.L. 2008. Effect of dietary modulation of the intestinal microbiota on reproduction and early growth. Theriogenology 70:286
6. Dawson, K.A. et al. 1990. Effects of microbial supplements containing yeast and lactobacilli on roughage fed ruminal microbial activities. J. Anim. Sci. 68:3392
7. Dhanoa, M.S. 1988. On the analysis of Dacron bag data for low degradability feeds. Grass and Forage Science. 43:441
8. Galina, M.A. et al. 2004. Effect of a slow intake urea supplementation on growing kids feed corn stubble or alfalfa with a balanced concentrate. Small Rum. Res. 53 (1-2):29
9. Galina, M.A. et al. 2007. Fattening Pelibuey lambs with sugar cane tops and corn complemented with or without slow intake urea supplement. Small Rum. Res.70:101
10. Galina, M. et al. 2009. Effect of Lactobacilli probiotic supplementation on blood glucose, insulin and NEFA performance of dairy cattle during late pregnancy and early lactation. In:Ruminant Physiology. Digestion, metabolism, and effects of nutrition on reproduction and welfare. Wageningen Academic Publisher. The Netherlands. p. 512
11. Goering, H.K. & Van Soest, P.J. 1970. Forage fiber analysis (apparatus, reagents, procedures, and some applications). United States Department of Agriculture. Agriculture Handbook. No. 379. Agricultural Research Service. Wachington, D.C.
12. Hungate, P.E. 1969. A roll tube method for cultivation of strict anaerobes. In: Microbiology. (Norris, J.R., Ribbons, D.W., Eds.). Academic Press, London. p. 117
13. Jouany, J.P. et al. 1998. Effect of Saccharomyces cerevisiae and Aspergillus oryzae on the digestion of nitrogen in the rumen of defaunated and refaunated sheep. Anim. Feed Sci. Technol. 75:1
14. McDonald, I. 1981. A revised model the estimation of protein degradability in the rumen. J. of Agric. Sci. Cambridge. 96:251
15. Newbold, C.J. et al. 1995. Different strains of Saccharomyces cerevisiae differ in their effects on ruminal bacterial numbers in vitro and in sheep. J. Anim. Sci. 73:1811
16. Nocek, J.E. et al. 2002. Ruminal supplementation of direct fed microbial on diurnal pH variation and in situ digestion in dairy cattle. J. Dairy Sci. 85:429
17. Nocek, J.E. et al. 2003. Direct-fed microbial supplementation on the performance of dairy cattle during the transition period. J. Dairy Sci. 86:331
18. Ørskov, E.R. et al. 1980. The use of the nylon bag technique for the evaluation of feedstuffs. Tropical Animal Production. 5:195
19. Ørskov, E.R. & McDonald, I. 1979. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J. Agric. Sci. 96:499
20. Ortíz, M.A. et al. 2009. Effect of a slow nitrogen intake supplementation with or without a lactic probiotic in Pelibuey lamb growth. Nutritional and Foraging Ecology of Sheep and Goats. Options Méditerranéennes, Serie A. 85:309
21. Patra, A.K. 2007. Nutritional management in organic livestock farming for improved ruminant health and production- An overview. Livest. Res. Rural Dev. 19 (3) Article#41. http://www.lrrd.org/lrrd19/3/part.19041.htm
22. Reid, G. & Friendship, R. 2002. Alternatives to antibiotic use: probiotics for the gut. Anim. Biotech. 13:97
23. SAS. 1996. User's Guide: Statistics. Version 6.1. SAS Institute, Inc, Cary, NC, USA.
24. Satter, L.D. & Slyter, L.L. 1974. Effect of ammonia concentration on rumen microbial protein production in vitro. Brit. J. Nutr. 32:194
25. Yasuda, K. et al. 2007. A new symbiotic consisting of Lactobacillus casei subsp. casei and dextran improves milk production in Holstein dairy cow. J. Vet Med Sci. 69:205
Recibido el 2 de octubre del 2009
Aceptado el 19 de octubre del 2009

{kind=link}
{kind=link}
{kind=link}
{kind=link}