










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPastos y Forrajes
versión impresa ISSN 0864-0394
Pastos y Forrajes vol.34 no.3 Matanzas jul.-sep. 2011
ARTÍCULO DE INVESTIGACIÓN
Riqueza y abundancia de la macrofauna edáfica en cuatro usos de la tierra en las provincias de Artemisa y Mayabeque, Cuba
Richness and abundance of soil macrofauna in four land uses of the Artemisa and Mayabeque provinces, Cuba
Grisel Cabrera1, Nayla Robaina2 y D. Ponce de León3
1Instituto de Ecología y Sistemática, CITMA, Carretera de Varona km 3½, Capdevila, Boyeros, C.P. 10800 Ciudad de La Habana, Cuba
E-mail: grisel17@ecologia.cu
2 Estación Experimental de Plantas Medicinales, Güira de Melena, Artemisa, Cuba
3 Facultad de Agronomía, Universidad Agraria de La Habana, Mayabeque, Cuba
RESUMEN
Se evaluó el efecto de la intensidad de uso de la tierra sobre la riqueza y abundancia de las comunidades de la macrofauna del suelo, en un gradiente desde bosques secundarios y pastizales hasta cultivos varios destinados a la producción de papa y cañaverales. El estudio se realizó al final del período lluvioso del año 2009, en el mes de octubre, en las provincias Artemisa y Mayabeque, donde están localizados los diferentes sistemas de uso de la tierra. La macrofauna edáfica fue recolectada según la metodología propuesta por el TSBF. Se evaluó la riqueza taxonómica, la densidad y la biomasa de la macrofauna del suelo, y en el análisis de los datos se usaron pruebas no paramétricas. Los mayores valores de riqueza, densidad y biomasa de la macrofauna se obtuvieron en los bosques secundarios, y los menores valores en los pastizales, los cultivos varios y los cañaverales. Haplotaxida, Formicidae, Isoptera, Coleoptera y Diplopoda fueron las unidades taxonómicas de la macrofauna, dominantes en densidad y en biomasa, en la mayoría de los usos de la tierra estudiados. Los resultados sobre la riqueza taxonómica, la densidad y la biomasa de la macrofauna del suelo indicaron el nivel de degradación del medio edáfico, debido a la intensidad de uso de la tierra.
Palabras clave: Fauna del suelo, uso múltiple de la tierra.
ABSTRACT
The study assessed the effect of the intensity of different land uses (secondary forests, pasturelands, fields with varied crop cultivation dedicated to potato production and sugarcane plantations) on the richness and abundance of soil macrofauna communities. The research was conducted in October, at the end of the rainy season, 2009, in the Artemisa and Mayabeque provinces where the different land use systems are located. The soil macrofauna was collected using the methodology proposed by the TSBF program. The taxonomic richness, density and biomass of soil macrofauna were evaluated and the data processing included nonparametric tests. The highest values of richness, density and biomass were obtained in the secondary forests and the lowest values in the pasturelands, varied crop cultivation and sugarcane plantations. Haplotaxida, Formicidae, Isoptera, Coleoptera and Diplopoda were the prevailing taxonomic units of the macrofauna regarding density and biomass in most of the land uses. The results about taxonomic richness, density and biomass of the edaphic macrofauna indicated the degree of soil degradation due to land use intensity.
Key words: Multiple land use, soil fauna.
INTRODUCCIÓN
Los macro-invertebrados edáficos (mayores de 2 mm de diámetro) actúan como agentes determinantes en la fertilidad del suelo y, por ende, en el funcionamiento global del sistema edáfico. Esta fauna puede ser afectada por diferentes usos y manejos de la tierra. Debido a su susceptibilidad y rápida respuesta ante los cambios en la cobertura, la transformación de la vegetación, el comportamiento ante distintas variables ambientales y la actividad ecológica que desempeñan, muchos autores proponen su uso como indicadores de calidad o alteración ambiental (Lavelle et al., 2003).
En Cuba son pocas las investigaciones acerca de la macrofauna edáfica que involucran todos los grupos que la componen. Entre los primeros estudios en el país se pueden señalar los realizados sobre la macrofauna de la hojarasca en ecosistemas boscosos, especialmente de la Sierra del Rosario (González y López, 1987; Prieto y Rodríguez, 1996). Con posterioridad, la mayoría de los estudios ecológicos han abordado grupos específicos de la macrofauna, principalmente las lombrices de tierra y los diplópodos en ecosistemas naturales y/o perturbados, y el efecto que tales perturbaciones provocan en estas comunidades (Rodríguez, 2000; Prieto et al., 2003).
Otros trabajos en el país han caracterizado la macrofauna del suelo en diferentes sistemas de uso de la tierra, con el objetivo de evaluar, a partir de las características de estas comunidades edáficas, el tipo de uso o formación vegetal, y el manejo de los suelos y de los cultivos (Rodríguez et al., 2002; Sánchez y Reyes, 2003; Cabrera-Dávila et al., 2007; Serrano, 2010). No obstante, se reconoce la necesidad de profundizar e incrementar las investigaciones en tal sentido. Los futuros estudios deben identificar índices, o específicamente taxones, que puedan ser usados como indicadores de la alteración ambiental.
En el trabajo se evaluó el efecto de la intensidad de uso de la tierra sobre la riqueza y abundancia de las comunidades de la macrofauna del suelo, en un gradiente desde bosques secundarios y pastizales hasta cultivos varios destinados a la producción de papa y cañaverales, ubicados en las provincias Artemisa y Mayabeque.
MATERIALES Y MÉTODOS
La investigación se realizó en el mes de octubre, en el período lluvioso del año 2009, en 11 sistemas de referencia a todo lo largo de la Llanura Roja de la antigua provincia La Habana, actualmente localizada entre las provincias Artemisa y Mayabeque. Se trabajaron cuatro usos de la tierra: bosques secundarios, pastizales, cultivos varios destinados a la producción de papa y cañaverales. El tipo de suelo en todos los usos fue Ferralítico Rojo, según la clasificación de Hernández et al. (1999).
Descripción de los sistemas de uso de la tierra
Bosques secundarios. Se estudiaron tres áreas de bosque en las localidades de Managua (22°56'44.80" N y 82°16'11.07" W), Nazareno (22°58'05.40" N y 82°14'02.72" W) y Aguacate (22°59'17.90" N y 81°50'01.03" W), situadas en la provincia Mayabeque. Son sistemas boscosos de regeneración natural o con diferente nivel de antropización, con edades de 40 a 120 años y extensiones entre dos y cinco hectáreas. Presentan elementos florísticos de vegetación semidecidua, así como especies arbóreas frutales y maderables, con estratos herbáceos y arbustivos que dan diferentes grados de cobertura al suelo. Entre las especies arbóreas se destacan: mamey colorado (Pouteria sapota H.E. Moore & Stearn), mango (Mangifera indica L.), caimito (Chrysophyllum cainito L.), framboyán rojo (Delonix regia (Boj. ex Hooker) Raf.), palma real(Roystonea regia (Kunth) O. F. Cook), baría (Cordia gerascanthus L.), ocuje (Calophyllum inophyllum L.), yamagua (Guarea guidonia (L.) Sleumer), teca (Tectona grandis L. fil.), caoba de Cuba (Swietenia mahagoni (L.) Jacq.),cedro (Cedrela odorata L.), pino (Pinus sp.) y majagua (Taliparitielatum (L.) Fryxell).
Pastizales. Se seleccionaron dos sistemas de pastizales en el municipio San José de las Lajas de la provincia Mayabeque, con una extensión máxima de 12 ha, ubicados en las siguientes coordenadas (pastizal 1: 23°00'01.50" N y 82°09'49.10" W; pastizal 2: 22°53'52.10" N y 82°02'08.12" W). Se trata de pastizales naturalizados con más de 25 años de explotación. Las principales especies de pasto eran: hierba de guinea (Panicum maximum Jacq.) y pasto estrella (Cynodon nlemfuensis Vanderyst), que presentaban una cobertura entre 80 y 90%. En estos pastizales pastoreaba ganado vacuno de las razas Holstein y Siboney, y el manejo era principalmente rotacional racional intensivo, con una carga animal media de 2,8 UGM ha-1.
Cultivos varios. Se seleccionaron tres sistemas localizados en los municipios Güira de Melena (22°45'40.50" N y 82°29'21.71" W), Batabanó (22°46'42.40" N y 82°15'08.27" W) y Güines (22°47'43.60" N y 82°02'31.46" W), pertenecientes a Artemisa y Mayabeque. El cultivo principal era la papa (Solanum tuberosum L.), en constante rotación con boniato (Ipomoea batatas (L.) Lam.), malanga (Colocasia esculenta (L.) Schott), frijol (Phaseolus vulgaris L.), maíz (Zea mays L.), calabaza (Cucurbita moschata (Duch. ex Lam.) Duch. ex Poir) y yuca (Manihot esculenta (L.) Crantz). Entre la vegetación indeseable en las áreas se destacaban: don Carlos (Sorghum halepense (L.) Pers.), cebolleta (Cyperus rotundus L.), bledo (Amaranthus hybridus L.) y romerillo (Bidens pilosa L.). Se trata de áreas dedicadas a la agricultura por más de 10 años, fundamentalmente con labranza tradicional y riego eléctrico por aspersión. La aplicación de NPK en estos sistemas fue de 1 490 kg ha-1 año-1 y la de urea fluctuó entre 224 y 298 kg ha-1 año-1.
Cañaverales. Las tres áreas en explotación con caña de azúcar (Saccharum officinarum L.) se ubicaron en Artemisa y Mayabeque, en los municipios Güira de Melena (22°50'24.80" N y 82°26'50.56" W), San Nicolás de Bari (22°46'32.60" N y 81°55'05.90" W) y Madruga (22°58'47.00" N y 81°50'49.24" W). Ocupaban estas áreas las variedades CP 52-43 y C 86-12, C 323-68 y C 86-56, respectivamente. Las plantas arvenses fundamentales en los sistemas eran: don Carlos, cebolleta, sancaraña (Rottboellia cochinchinensis L.F.) y pata de gallina(Eleusine indica (L.) Gaertn). Se empleaban sistemas con riego por gravedad y sistema eléctrico de enrrolladores; la fertilización química fue de: 50-60 kg ha-1 año-1 de urea, 20-83 kg ha-1 año-1 de K2O y 25 kg ha-1 año-1 de P2O5.
Muestreo y procesamiento de la macrofauna edáfica
La recolección de la macrofauna se realizó según la Metodología del Programa Internacional «Biología y Fertilidad del Suelo Tropical» o TSBF (Anderson e Ingram, 1993; Lavelle et al., 2003). Por área o réplica de uso de la tierra se tomaron ocho monolitos de suelo (25 x 25 x 30 cm), distanciados a 20 m en un diseño completamente aleatorizado, para un total de 24 monolitos procesados en cada uno de los usos bosques secundarios, cultivos varios y cañaverales, y 16 monolitos en el uso de pastizales. La macrofauna se recolectó manualmente in situ, y se preservó principalmente en alcohol a 75%.
La macrofauna se separó hasta el nivel taxonómico de orden y familia (Borror et al., 1976; Sims, 1980; Brusca y Brusca, 2003). Se realizó un acercamiento a la riqueza taxonómica de estas comunidades en cada uso de la tierra, estimada a partir del número de familias que pudieron ser identificadas. Los órdenes de la macrofauna con menor representación en este estudio, como Blattodea, Diptera, Hemiptera, Orthoptera y Lepidoptera, y también Araneae, no se incluyeron en el análisis de la riqueza taxonómica.
Se calcularon los valores promedios de densidad (ind.m-2) y biomasa (gm-2) para las comunidades edáficas y para las diferentes unidades taxonómicas de la macrofauna, en cada uso de la tierra. La densidad se determinó a partir del número de individuos y la biomasa, sobre la base del peso húmedo en la solución preservante.
Con el propósito de determinar las variaciones de la densidad y la biomasa de las comunidades de la macrofauna del suelo entre usos, se empleó el análisis no paramétrico de Kruskal-Wallis, y como prueba a posteriori para la comparación de medias se utilizó la U de Mann-Whitney con ajuste de Bonferroni. El procesamiento estadístico se realizó a través del programa PAST-1.75, 2001.
RESULTADOS
Composición y riqueza taxonómica
Las comunidades de la macrofauna del suelo en los cuatro usos de la tierra estudiados en las provincias Artemisa y Mayabeque, comprenden tres phyla, siete clases, 22 órdenes y 39 familias determinadas (tabla 1).
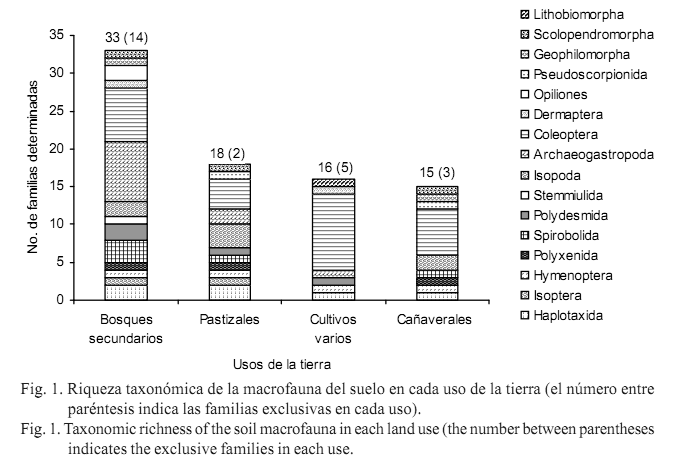
El análisis a nivel de orden mostró 19 órdenes en los bosques secundarios, 14 en los pastizales y los cañaverales, y solo 12 órdenes en el uso de cultivos varios. La riqueza taxonómica, valorada a nivel de familias identificadas, mostró también un mayor número en los bosques secundarios (33), seguido de los pastizales (18) y de forma muy cercana los cultivos varios (16) y los cañaverales (15). Los órdenes mejor representados en número de familias en los bosques secundarios, fueron Archaeogastropoda y Coleoptera con ocho y siete familias, respectivamente. En los restantes usos el orden con mayor número de familias fue Coleoptera, con su máxima representación en el uso de cultivos varios (10 familias) que tuvo alguna incidencia como plaga agrícola. En el uso de pastizales, este orden tuvo cuatro familias y en los cañaverales seis, y los otros órdenes de la macrofauna en todos los usos solo presentaron entre una y tres familias. De igual manera, se obtuvieron 14 familias exclusivas de los bosques secundarios, dos de los pastizales, cinco de cultivos varios y tres de los cañaverales (fig. 1).
Densidad y biomasa
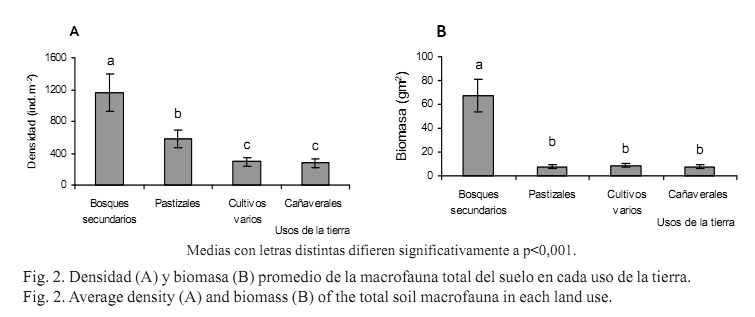
La densidad y la biomasa promedio de la macrofauna total del suelo en los bosques secundarios fueron de 1 166,60 ind.m-2 y 67,45 gm-2, respectivamente, valores considerablemente superiores a los obtenidos en los pastizales (581 ind.m-2 y 7,71 gm-2), en los cultivos varios (294 ind.m-2 y 8,79 gm-2) y en los cañaverales (280 ind.m-2 y 7,85 gm-2). La prueba de Kruskal-Wallis confirmó esta marcada variación, ya que mostró diferencias altamente significativas entre los usos en densidad (p<0,001) y en biomasa (p<0,001) (figs.2 A y B).
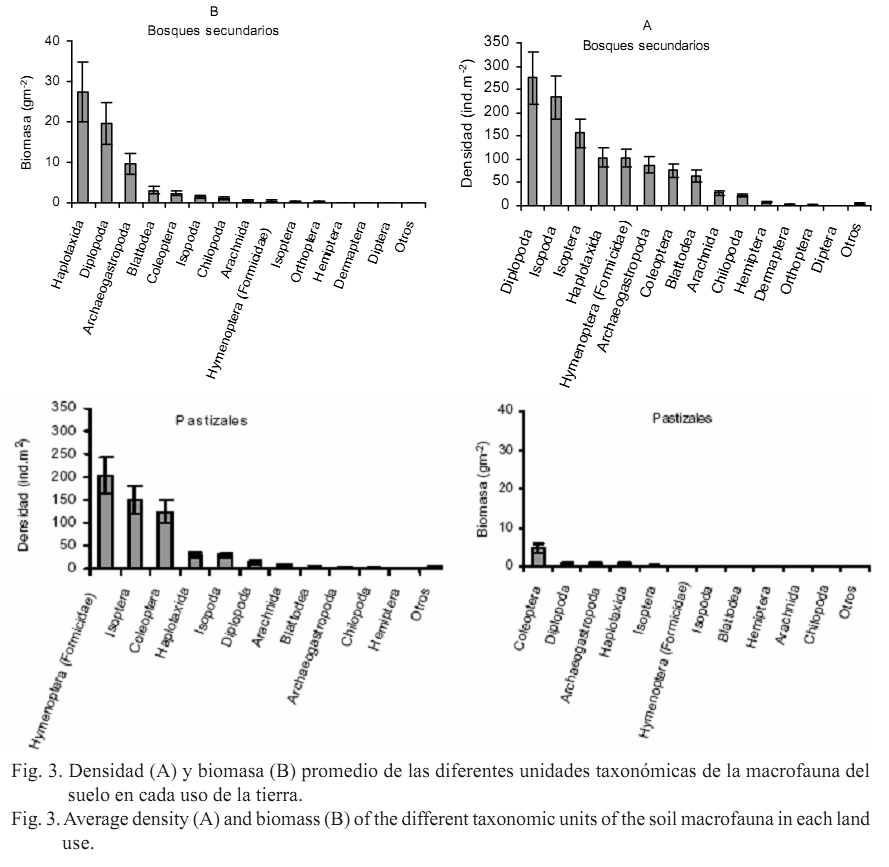
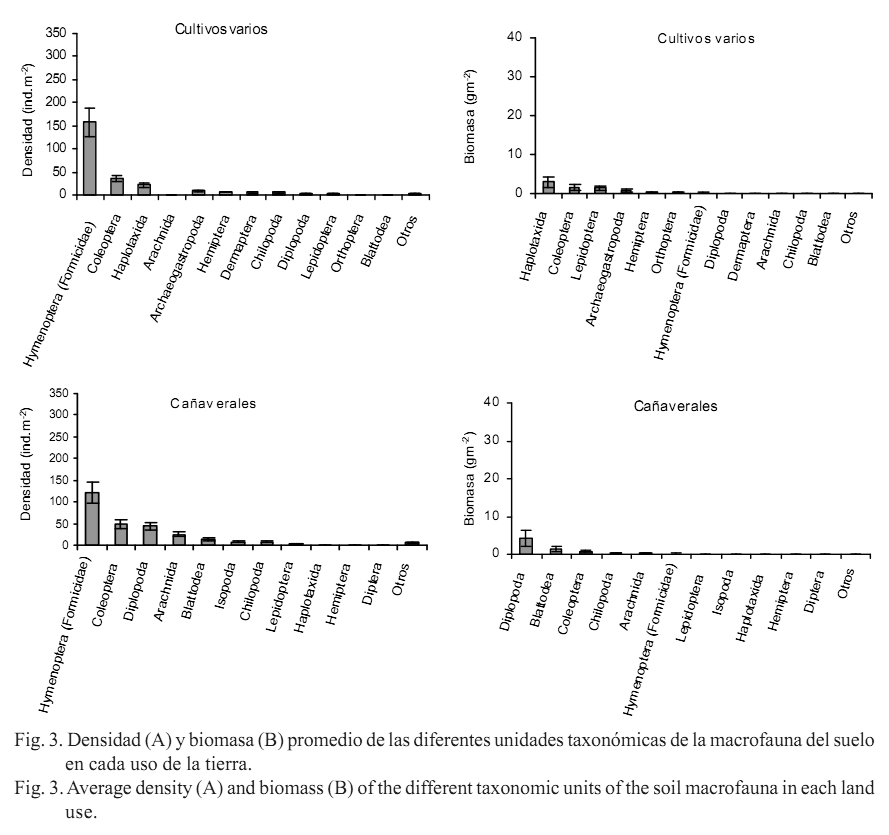
De acuerdo con la dominancia en densidad y biomasa de los diferentes grupos taxonómicos de la macrofauna (figs. 3 A y B), se observó que en los bosques secundarios los taxones más importantes en densidad incluyeron a Diplopoda, Isopoda e Isoptera, y los mayores componentes en biomasa fueron Haplotaxida y Diplopoda.
En los pastizales se destacaron Hymenoptera (Formicidae), Isoptera y Coleoptera con valores altos y cercanos en densidad; mientras en biomasa dominó esencialmente Coleoptera. En los cultivos varios se destacó Hymenoptera (Formicidae) en densidad, y Haplotaxida y Coleoptera fueron los principales representantes en biomasa. En los cañaverales prevalecieron, en densidad, Hymenoptera (Formicidae), Coleoptera y Diplopoda, y en biomasa solo Diplopoda.
DISCUSIÓN
Composición y riqueza taxonómica
La cantidad de familias exclusivas encontradas en cada uso, permitió inferir que todos los usos de la tierra compartieron un número alto de familias. Los sistemas en estudio poseen cierto grado de intervención o alteración por el hombre (bosques secundarios o intervenidos, pastizales con manejo ganadero, y cultivos con un intenso laboreo agrícola), donde pueden colonizar igualmente familias con características semejantes de tolerancia a un variado rango de condiciones edafoclimáticas y, por tanto, adaptables y resistentes a las perturbaciones inducidas.
Tanto el mayor valor de riqueza de familias como el de exclusividad alcanzados en los bosques secundarios, evidenciaron la importancia de la variabilidad florística y de la heterogeneidad de los recursos en el suministro de las fuentes de alimento y refugio necesarias para conservar la diversidad de las comunidades edáficas (Fragoso y Lavelle, 1992). Ello también se relacionó con la cobertura presente en estos bosques, que provee un aporte considerable de hojarasca y sombra para mantener estables los valores de temperatura y humedad en el suelo, todo lo cual favoreció el desarrollo de comunidades más diversas. En este sentido, Granados y Barrera (2007) encontraron mayor número de especies, géneros y familias en relictos de bosques en comparación con las áreas sin cobertura arbórea y, de modo general, concluyeron que las áreas con mayor diversidad y riqueza de plantas y un porcentaje más alto de cobertura pueden manifestar una mayor diversidad de la macrofauna del suelo.
En Cuba, González y López (1987) reconocieron en ecosistemas forestales un número alto de taxones sobresalientes de la macrofauna, entre ellos Hymenoptera (Formicidae), Coleoptera y Haplotaxida. Más recientemente, en un relicto de bosque con características de vegetación original semidecidua en la costa norte de La Habana, Serrano (2010) halló un total de 21 unidades taxonómicas y los órdenes dominantes resultaron ser Hymenoptera, Araneae e Isopoda. Este valor está próximo a la riqueza referida en el presente estudio para los sistemas boscosos (19 órdenes y 33 familias), a pesar de ser de formación secundaria.
Rodríguez et al. (2002) y Sánchez y Reyes (2003) obtuvieron en el país, en pastizales con diferente manejo ganadero, hasta nueve órdenes de la macrofauna edáfica. Por su parte, Cabrera-Dávila et al. (2007) encontraron en áreas de policultivos, en pastizales asociados con cobertura arbórea de leguminosas y en monocultivo de gramíneas, valores superiores de riqueza de órdenes (14 en los dos primeros sistemas y 12 en el último) y de familias (30, 26 y 22, respectivamente), con un predominio de formícidos, haplotaxidos, diplópodos e isópodos.
También a escala mundial en varios trabajos se hallaron resultados cercanos a los alcanzados en este estudio, a pesar de que en la mayoría el área de recolecta fue más grande (mayor cantidad de monolitos por sistemas de uso de la tierra). Pashanasi (2001) recolectó en diferentes sistemas de uso de la tierra en la Amazonía peruana, entre 20 y 30 unidades taxonómicas de la macrofauna, en bosques secundarios con 20 y cinco años de regeneración natural; en pastizales entre 18 y 23 taxones y en cultivos anuales entre 15 y 25 unidades taxonómicas. En los ecosistemas de Brasil hubo un decremento gradual de la diversidad de la macrofauna ante los cambios en la intensidad de uso de la tierra, donde se examinaron desde los bosques, los sistemas agroforestales y los pastizales, hasta los sistemas de cultivos anuales (Barros et al., 2002). Otros autores en Perú registraron una mayor cantidad de grupos taxonómicos en los bosques primarios y secundarios (13), en comparación con sistemas de cultivos (12) (Villavicencio et al., 2009).
Densidad y biomasa
Los resultados de la densidad y la biomasa demostraron la sensibilidad de las comunidades de la macrofauna ante el manejo del suelo, los cambios en la cobertura y la transformación de la vegetación, así como el efecto negativo de las perturbaciones impuestas por los sistemas de cultivo.
Múltiples elementos pueden enumerarse como causantes de los valores reducidos de densidad y biomasa en los usos pastizales, cultivos varios y cañaverales, con respecto a los alcanzados en los bosques secundarios (figs. 2 A y B). Numerosos autores han mencionado y comprobado que uno de los principales factores es la pérdida de la materia orgánica del suelo (Velásquez et al., 2009), por la erosión hídrica acelerada debido a la disminución de la cobertura vegetal, como uno de los posibles motivos que explican este suceso.
La densidad y la biomasa también se han relacionado con el nivel de macronutrientes, el pH, la textura del suelo, la capacidad de retención de agua y la alta calidad de la hojarasca (Barros et al., 2002). Lok (2005) planteó que los organismos de la macrofauna prefieren los restos vegetales con una relación C/N relativamente baja, por ser de más fácil acceso y descomposición, lo que explica su fuerte selectividad hacia la vegetación que existe sobre el suelo. Las plantas y los desechos orgánicos proveen los hábitats y los alimentos para la edafofauna, y el adecuado contenido de materia orgánica favorece no solo las propiedades físico-químicas, sino también proporciona una fuente energética que estimula la actividad de los invertebrados del suelo.
Otro aspecto que se debe resaltar es la estructura de la vegetación, principalmente la presencia de diferentes estratos. La cobertura arbórea y/o arbustiva en el ordenamiento de los ecosistemas es de suma importancia, pues garantiza la entrada continua y abundante de material orgánico vegetal y un microambiente más favorable para el establecimiento de los macroinvertebrados del suelo (Rodríguez et al., 2002).
Existen pocos estudios en Cuba sobre la macrofauna en los ecosistemas boscosos, con un nivel moderado de conservación. Los valores de densidad y biomasa de la macrofauna en el uso bosques secundarios son comparables con los obtenidos en el país, en diferentes sistemas agroforestales con manejo silvopastoril e influencia de las leguminosas arbóreas. Cabrera-Dávila et al. (2004) obtuvieron en este tipo de sistema una densidad superior en la época de mayor pluviosidad (2 346 ind.m-2) y una biomasa de 66,90 gm-2; otros investigadores cubanos hallaron estimados inferiores (Lok, 2005; Sánchez, 2007). En investigaciones recientes, en un bosque seminatural del litoral norte de La Habana, Serrano (2010) encontró una densidad de la macrofauna menor que la de este estudio (835 ind.m-2). Con referencia a otras recolectas recientes efectuadas en los bosques primarios siempreverdes de la Sierra del Rosario en Artemisa (Cabrera-Dávila, G. y Hernández, A. inédito) también se hallaron valores inferiores (706 ind.m-2 de densidad y 42,50 gm-2 de bioma-sa) que los observados en los ecosistemas de bosques secundarios en estudio (1 166,60 ind.m-2 y 67,45 gm-2). Se debe tener en cuenta que este tipo de ecosistema puede albergar una parte de la fauna de la vegetación original, y además ser colonizado en mayor proporción por especies oportunistas e invasoras, propias de terrenos con algún nivel de perturbación.
Las investigaciones en los bosques secundarios y en las plantaciones forestales del trópico húmedo reportaron estimados semejantes a los del presente trabajo. Pashanasi (2001), en algunos bosques secundarios de Perú (20 y cinco años de regeneración natural), registró una densidad entre 485 y 838 ind.m-2 y la biomasa entre 33,9 y 102,0 gm-2. En Brasil Barros et al. (2002), en los bosques y los sistemas agroforestales, encontraron entre 884 y 1 761 ind.m-2, y en biomasa obtuvieron valores bajos (entre 10,18 y 9,46 gm-2, respectivamente). Tsukamoto y Sabang (2005) en plantaciones arbóreas de Malasia, calcularon una densidad de más de 1 000 ind.m-2 y 60 gm-2 de biomasa de macroinvertebrados.
El uso de pastizales fue el segundo en densidad, pero en biomasa fue el de más bajos valores (figs.2 A y B), e igualmente ambas variables fueron menores que las de otros pastizales cubanos estudiados anteriormente. En pastizales de hierba de guinea Rodríguez et al. (2002) obtuvieron una biomasa de 34,61 ind.m-2 ; mientras que Sánchez (2007) informó en este tipo de pastizal una densidad de solo 88,87 ind.m-2 y una biomasa de 6,51 gm-2.
El régimen de pastoreo y, por ende, el pisoteo de los animales, pudo haber afectado la densidad y la biomasa de la macrofauna en el pastizal. Lok (2005) expresó que la compactación de los suelos de los pastizales por una determinada carga ganadera, puede reducir la población de invertebrados edáficos.
En otros pastizales del mundo, también la densidad y la biomasa de la macrofauna fueron superiores a las obtenidas en los del presente estudio. En pastizales de México, Perú, Brasil y Uruguay la densidad fue mayor que 600 ind.m-2 y la biomasa de 35 gm-2 (Brown et al., 2001; Barros et al., 2002; Huerta-Lwanga et al., 2008); sobre todo la biomasa en este tipo de ecosistemas ha estado influida por la contribución, en peso, de las lombrices de tierra.
En los agroecosistemas, las comunidades de la macrofauna del suelo son generalmente muy bajas debido a las alteraciones causadas por la perturbación, en correspondencia con lo planteado. Esta investigación siguió ese patrón, tanto en el uso de cultivos varios como en el de cañaverales (figs. 2 A y B). Los valores de biomasa ligeramente más altos en dichos usos con respecto a los pastizales, pueden justificarse a través del predominio de determinados grupos detritívoros en los cañaverales, y de las lombrices de tierra en los cultivos varios (figs. 3 A y B). En Cuba son escasos los resultados sobre la respuesta de la macrofauna ante la evaluación de diferentes procedimientos agrícolas en sistemas de cultivo. Solamente Cabrera-Dávila et al. (2004), en áreas policultivadas del país con la aplicación de métodos agroecológicos (fertilización orgánica, rotación de cultivos y asociación de cultivos de ciclo corto con ciclo largo), hallaron una densidad de 2 016 ind.m-2 y biomasa de 63 gm-2, similares a las que se pueden apreciar en los ecosistemas bien conservados; estos resultados son contrarios a los hallados en las áreas cultivadas de este estudio, que tuvieron prácticas más agresivas como la fertilización química, la labranza mecánica y la introducción de maquinaria.
A nivel internacional algunos autores refieren, para los cultivos anuales, valores de bajos a intermedios tanto en densidad (362-851 ind.m-2) como en biomasa (5,10-32,40 gm-2) (Pashanasi, 2001; Barros et al., 2002), los que son superiores a los mostrados para las áreas en estudio. En otros agroecosistemas tropicales también se observó una menor biomasa (< 20 gm-2) y densidad (< 250 ind.m-2) con relación a los sistemas naturales (Villalobos et al., 2000; Brown et al., 2001; Huerta-Lwanga et al., 2008).
El análisis de la dominancia de determinados grupos de la macrofauna evidenció que el orden Haplotaxida no desempeñó un papel preponderante en el uso de pastizales, como ha sido comúnmente observado para otros pastizales naturales o inducidos del mundo, debido a la influencia de las lombrices de tierra, de amplia distribución y de gran talla (Feijoo et al., 2007).
Por lo general, las lombrices predominan en biomasa en la mayoría de los ecosistemas, pero según Lavelle et al. (1994) tienen mayor contribución en los pastizales de regiones húmedas y decrecen en las áreas forestales y en las zonas secas.
Otros grupos, como Isoptera, adquieren importancia sobre todo en zonas de cultivos, y su invasión y agresividad en estos sitios están relacionadas con la pobre cantidad y calidad de material orgánico, el incremento de la temperatura y la reducida humedad en el suelo (Barros et al., 2002). No obstante, diversos autores comentan sobre la afectación de las comunidades de termitas, primariamente de las especies humívoras que habitan en el suelo, en función de la degradación de los hábitats (Bandeira et al., 2003). De hecho en el presente trabajo, las termitas (representadas principalmente por una especie humívora) se encontraron solo en los usos bosques secundarios y pastizales, y no en los de mayor afectación como consecuencia de las prácticas agrícolas. Cunha (2006) señaló que estos organismos son los primeros colonizadores en los ambientes deforestados con suficiente material leñoso remanente, y ayudan en su descomposición.
CONCLUSIONES
El uso de bosques secundarios presentó valores altos de riqueza taxonómica, densidad y biomasa de la macrofauna edáfica, en correspondencia con una mayor estabilidad y un menor grado de intervención, al no tener actividades de laboreo continuo ni pastoreo, con relación a los restantes usos. Los usos de pastizales, cultivos varios y cañaverales tuvieron valores más bajos, lo que evidenció un nivel de intermedio a superior de antropización, debido al manejo ganadero en los pastizales, y al laboreo agrícola constante en los sistemas de cultivos. Los resultados sobre la riqueza taxonómica, la densidad y la biomasa de la macrofauna del suelo indicaron el nivel de perturbación del medio edáfico debido a la intensidad de uso de la tierra.
AGRADECIMIENTOS
Los resultados se obtuvieron dentro del marco de la Maestría de la Ciencia del Suelo de la Universidad Agraria de La Habana, y fueron financiados por el Proyecto de Investigación 08-22 «Causas de la degradación de la estructura de los suelos Ferralíticos Rojos de la Llanura Roja de La Habana», del PRCT Protección del Medio Ambiente y el Desarrollo Sostenible Cubano. Agradecemos también a diferentes especialistas cubanos por la clasificación taxonómica de la macrofauna edáfica, principalmente hasta el nivel de familia (Dr. L. F. de Armas, MCs. A. Lozada, Lic. T. Tcherva, Lic. M. Hernández, Lic. A. Alegre y Lic. R. Barba).
REFERENCIAS BIBLIOGRÁFÍCAS
1. Anderson, J.M. &. Ingram, J.S.I. Tropical soil biology and fertility: a handbook of methods. CAB International. Wallingford, United Kingdom. 236 p. 1993
2. Bandeira, A.G. et al. Effects of habitat disturbance on the termite fauna in a highland humid forest in the Caatinga Domain, Brazil. Sociobiology. 42 (1):117. 2003
3. Barros, E. et al. Effects of land-use system on the soil macrofauna in western Brazilian Amazonia. Biology and Fertility of Soils. 35 (5):338. 2002
4. Borror, D. et al. An introduction to the study of insects. 4th edition. Holt, Rinehart and Winston. New York, USA. 852 p. 1976
5. Brown, G. et al. Diversidad y rol funcional de la macrofauna edáfica en los ecosistemas tropicales mexicanos. Acta Zoológica Mexicana. Número especial 1:79. 2001
6. Brusca, R. & Brusca, G. Invertebrates. Sinauer Associates. Sunderland, Massachusetts, USA. Segunda edición. 966 p. 2003
7. Cabrera Dávila, G. et al. Variación estacional de la macrofauna del suelo en áreas con manejo agrícola - ganadero. Poeyana. 491:19. 2004
8. Cabrera Dávila, G. et al. La macrofauna del suelo en sistemas agroecológicos en Cuba. Brenesia. 67:45. 2007
9. Cunha, H. da. Cupins (Isoptera) bioindicadores para Conservação do Cerrado em Goiás. Tesis presentada en opción al grado científico de Doctor en Ciencias Ambientales. Universidad Federal de Goiás. Brasil. 79 p. 2006
10. Feijoo, A. et al. Relaciones entre el uso de la tierra y las comunidades de lombrices en la cuenca del río La Vieja, Colombia. Pastos y Forrajes. 30 (2): 235. 2007
11. Fragoso, C. & Lavelle, P. Earthworms communities of tropical rain forest. Soil Biol. Biochem. 24 (12):1397. 1992
12. González, R. & López, R. La macrofauna de la hojarasca y del suelo de algunos ecosistemas forestales de Cuba. Reporte de Investigación, Instituto de Zoología. 46:1. 1987
13. Granados, A. & Barrera, J. I. Efecto de la aplicación de biosólidos sobre el repoblamiento de la macrofauna edáfica en la cantera Soratama, Bogotá, DC. Universitas Scientiarum, Revista de la Facultad de Ciencias. Edición especial II. 12:73. 2007
14. Hernández, A. et al. Nueva versión de la clasificación genética de los suelos de Cuba. Ministerio de la Agricultura. La Habana, Cuba. 26 p. 1999
15. Huerta-Lwanga, E. et al. Relación entre la fertilidad del suelo y su población de macroinvertebrados. Terra Latinoamericana. 26 (2):171. 2008
16. Lavelle, P. et al. The relationship between soil macrofauna and tropical soil fertility. In: The biological management of tropical soil fertility (Eds. P.L.Woomer and M. J. Swift). John Wiley-Sayce. New York. p. 137. 1994
17. Lavelle, P. et al. Soil macrofauna. In: Trees, crops and soil fertility. Concepts and research methods. (Eds. G. Schroth and F.L.Sinclair). CABI Publishing. UK. p. 303. 2003
18. Lok, S. Determinación y selección de indicadores del sistema suelo-pasto en pastizales dedicados a la producción de ganado vacuno. Tesis presentada en opción al grado científico de Doctor en Ciencias Agrícolas. Instituto de Ciencia Animal. La Habana, Cuba. 98 p. 2005
19. Pashanasi, B. Estudio cuantitativo de la macrofauna del suelo en diferentes sistemas de uso de la tierra en la Amazonia Peruana. Folia Amazónica. 12 (1-2):75. 2001
20. Prieto, D. & Rodríguez, C. Índices de agregación de los invertebrados de la hojarasca en un bosque siempreverde de la reserva de la biosfera de la Sierra del Rosario, Pinar del Río, Cuba. Análisis comparativo. Rev. Biología. 10:27. 1996
21. Prieto, D. et al. Caracterización de la biodiversidad de la fauna edáfica cubana. Informe final de Proyecto. Programa Ramal «Sistemática y colecciones biológicas». Facultad de Biología, Universidad de La Habana, Cuba. 2003.
22. Rodríguez, C. Comunidades de lombrices de tierra en ecosistemas con diferente grado de perturbación. Rev. Biología. 14:147. 2000
23. Rodríguez, I. et al. Comportamiento de la macrofauna del suelo en pastizales con gramíneas naturales puras o intercaladas con leucaena para la ceba de toros. Rev. cubana. Cienc. Agríc. 36 (2):181. 2002
24. Sánchez, S. Acumulación y descomposición de la hojarasca en un pastizal de Panicum maximum Jacq. y en un sistema silvopastoril de P. maximum y Leucaena leucocephala (Lam.) de Wit. Tesis presentada en opción al grado científico de Doctor en Ciencias Agrícolas. Instituto de Ciencia Animal. La Habana, Cuba.135 p. 2007
25. Sánchez, S. & Reyes, F. Estudio de la macrofauna edáfica en una asociación de Morus alba y leguminosas arbóreas. Pastos y Forrajes. 26:315. 2003
26. Serrano, A. Estructura y dinámica de la comunidad de macroinvertebrados edáficos en dos formaciones vegetales de Boca de Canasí, La Habana, Cuba. Tesis presentada en opción al título académico de Máster en Zoología y Ecología Animal. Universidad de La Habana, Cuba. 75 p. 2010
27. Sims, R.W. A classification and the distribution of earthworms suborder Lumbricina (Haplotaxida: Oligochaeta). Bull. Br. Mus. Nat. Hist. (Zool.). 39 (2): 103. 1980
28. Tsukamoto, J. & Sabang, J. Soil macro-fauna in an Acacia mangium plantation in comparison to that in a primary mixed dipterocarp forest in the lowlands of Sarawak, Malaysia. Pedobiología. 49: 69. 2005
29. Velásquez, E. et al. Cambios en las comunidades de plantas influenciados por la macroagregación del suelo a través de las actividades de la macrofauna del suelo en la Amazonía Brasileira. http://www.iamazonica.org.br/conteudo/eventos/biodiversidadedesolo/ pdf/Resumos/Painel3_Velasquez E.pdf. 2009
30. Villalobos, F.J. et al. Patrones de la macrofauna edáfica en un cultivo de Zea maiz durante la fase postcosecha en la Mancha, Veracruz, México. Acta Zoológica Mexicana. 80:167. 2000
31. Villavicencio, D. et al. Macrofauna del suelo en diferentes sistemas de uso de la tierra en el parque nacional Tingo María, Huánuco, Perú. http://www.iamazonica.org.br/conteudo/eventos/biodiversidadedesolo/pdf/resumos/Poster_DaliaL.pdf. 2009
Recibido el 25 de noviembre del 2010
Aceptado el 16 de junio del 2011

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}