










My SciELO
 Custom services
Custom servicesServices on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkPastos y Forrajes
Print version ISSN 0864-0394
Pastos y Forrajes vol.35 no.2 Matanzas Apr.-June 2012
ARTÍCULO DE INVESTIGACIÓN
Comportamiento de la simbiosis leguminosa-rizobio en Centrosema plumieri inoculada con Bradyrhizobium sp. Ensayo de campo
Performance of the legume-rhizobium symbiosis in Centrosema plumieri inoculated with Bradyrhizobium sp. Field essay
C. J. Bécquer, J. A. Nápoles, Orquidia Álvarez, Yamilka Ramos y L. A. Palmero
Instituto de Investigaciones de Pastos y Forrajes, Estación Experimental Sancti Spíritus, Apdo 2255, Sancti Spíritus, Cuba
E-mail: pastossp@yayabo.inf.cu
RESUMEN
Se efectuó un experimento de campo, sin riego, en la Estación Experimental de Pastos y Forrajes de Sancti Spíritus, Cuba, durante el periodo de noviembre de 2001 a abril de 2002. El objetivo fue evaluar el comportamiento simbiótico de Centrosema plumieri, inoculada con cepas de Bradyrhizobium sp., previamente aisladas de leguminosas forrajeras procedentes de ecosistemas ganaderos de Cuba. El diseño experimental fue de bloques al azar; se utilizaron 11 tratamientos inoculados con cepas nativas, un testigo inoculado con la cepa de referencia 5030 (Bradyrhizobium sp.) y un control no inoculado, con tres réplicas. Se aplicó un análisis de varianza (ANOVA), así como de correlación y regresión. Los valores de los tratamientos inoculados fueron estadísticamente iguales a los del control absoluto, en cuanto al peso seco aéreo (PSA) y el rendimiento de N (RN), excepto el inoculado con la cepa HA2 en RN. En general, se demostró una débil efectividad de la inoculación. No obstante, en el índice de nodulación (IN) sobresalieron JH2 y HA2, con valores estadísticamente superiores (p<0,05) al resto. El grado de dependencia del RN y el PSA fue débil en los tratamientos inoculados con respecto al IN, aunque fuerte en el RN en relación con el PSA. Se concluye que las cepas nativas inoculadas en C. plumieri, con excepción de HA2 en el RN, así como JH2 y HA3 en el IN, no ejercieron un efecto altamente positivo en la planta. Estos resultados deben valorarse para la continuación de estudios relacionados con la simbiosis leguminosa-rizobio.
Palabras clave: Bradyrhizobium sp., Centrosema plumieri, inoculación.
ABSTRACT
A field trial, without irrigation, was conducted at the Sancti Spiritus Experimental Station of Pastures and Forages, Cuba, from November, 2001, to April, 2002. The objective was to evaluate the symbiotic performance of Centrosema plumieri, inoculated with Bradyrhizobium sp. strains, previously isolated from forage legumes of Cuban livestock production ecosystems. The experimental design consisted in randomized blocks; 11 treatments inoculated with native strains, a control inoculated with the reference strain 5030 (Bradyrhizobium sp.) and a non-inoculated control were used, with three replications. A variance analysis (ANOVA), as well as correlation and regression analyses, were applied. The values of the inoculated treatments were statistically equal to those of the absolute control, regarding aerial dry weight (ADW) and N yield (NY), except the one inoculated with the strain HA2 in NY. In general, a weak activity of inoculation was shown. Nevertheless, in the nodulation rate (NR) JH2 and HA2 stood out, with values statistically higher (p<0,05) than the others. The degree of dependence of NY and ADW was weak in the inoculated treatments with regards to NR, although it was strong in NY with regards to ADW. It is concluded that the native strains inoculated in C. plumieri, with the exception of HA2 in NY, as well as JH2 and HA3 in NR, did not exert a highly positive effect on this plant. These results should be evaluated for the continuation of studies related to the legume-rhyzobium symbiosis.
Key words: Bradyrhizobium sp., Centrosema plumieri, inoculation.
INTRODUCCIÓN
La simbiosis leguminosa-rizobio relación recíproca entre las plantas y la bacteria fue reconocida por primera vez hace más de 100 años, y su valor agrícola se ha demostrado a través de las investigaciones científicas realizadas en el mundo entero.
La importancia de la fijación biológica del nitrógeno para la seguridad de los alimentos en el mundo es realmente incuestionable, y el uso de cultivos capaces de efectuar una fijación simbiótica del N es el componente primario en una agricultura sostenible (Vance et al., 2000). Por otra parte, en Cuba existen trabajos que describen la eficiencia simbiótica de cepas nativas cubanas en leguminosas tropicales (Tang et al., 1994; Tang, 1996; López et al., 2002)
La provincia Sancti Spíritus se caracteriza por una amplia gama de géneros y especies de leguminosas nativas o naturalizadas, diseminadas en cuatro zonas edafoclimáticas (Hernández, 1989), y se pueden localizar en ecosistemas con características ambientales estresantes (acidez, alcalinidad o altas temperaturas, entre otras) tanto para las plantas como para los microorganismos.
Centrosema es un género neotropical que contiene especies muy importantes desde el punto de vista pratense y forrajero. Es nativo del sur de América tropical, abunda en los trópicos (Skerman et al., 1991) y se afirma que puede fijar hasta 280 kg de N/ha/año en asociación con gramíneas (Sylvester-Bradley et al., 1983). Es moderadamente específico en la nodulación, y el suelo puede carecer de cepas que lo infecten efectivamente, por lo que se recomienda su inoculación. En estudios realizados por Hernández et al. (1999), en leguminosas nativas o naturalizadas de la provincia Sancti Spíritus, se encontró que Centrosema fue uno de los géneros más adaptados a las condiciones edafoclimáticas de la provincia. Olivera et al. (2008) también hallaron este género ampliamente diseminado en tres provincias de Cuba. Dichos estudios posibilitan un trabajo más profundo sobre la utilización, en esas condiciones, de las especies naturales; se consideran como tales aquellas plantas que son originarias de un país determinado, o introducidas desde hace varios años, y que se encuentran adaptadas totalmente al ambiente (Funes, 2007).
El objetivo de este trabajo fue evaluar el comportamiento simbiótico de Centrosema plumieri, inoculada con cepas de Bradyrhizobium sp. en condiciones de campo, en Sancti Spíritus, Cuba.
MATERIALES Y MÉTODOS
El experimento se realizó en áreas de la Estación Experimental de Pastos y Forrajes Sancti Spíritus, Cuba (21o 53' 00'' latitud Norte, 79o 21' 25'' longitud Oeste, y a 40 msnm) y tuvo una duración de seis meses (noviembre de 2001-abril de 2002).
El suelo del área experimental corresponde al tipo Pardo con Carbonatos típico (Anon, 1979), deficitario en P2O5 y K2O, y con niveles medios de materia orgánica (tabla 1), lo cual es característico de este tipo de suelo (Hernández, A. et al., 1999).
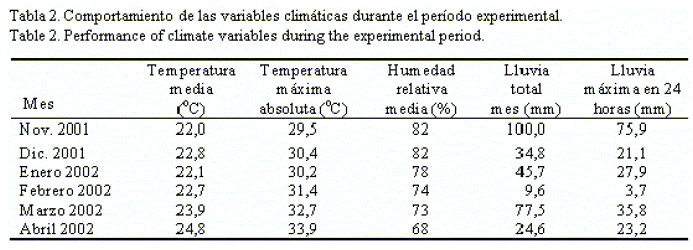
No se aplicó riego a pesar de que en el periodo evaluativo predominó la escasa precipitación, excepto en el mes de noviembre, fecha de siembra del experimento (tabla 2), lo cual propició la adecuada germinación de la semilla. Las semillas procedían de la Estación Experimental de Pastos y Forrajes de Sancti Spíritus. Se escarificaron en agua a 90°C, durante tres minutos, previamente a la siembra.
Las cepas nativas utilizadas en el experimento pertenecen a Bradyrhizobium sp. y fueron aisladas de C. plumieri y Centrosema virginianum, leguminosas procedentes de la zona centro de Sancti Spíritus, las que fueron caracterizadas taxonómicamente y evaluadas agronómicamente en condiciones de invernadero (Bécquer, 2002). Se utilizó también la cepa de referencia 5030 (Bradyrhizobium sp.), donada por el Instituto de Investigaciones de Pastos y Forrajes de Cuba (tabla 3).

Las cepas se conservaron en cuñas agarizadas de medio sólido levadura-manitol, a 4ºC (Vincent, 1970). Los inóculos se prepararon mediante el crecimiento de las cepas en erlenmeyers de 250 mL, que contenían 100 mL de medio líquido levadura-manitol, y estas fueron incubadas a 29-30oC, de cinco a ocho días, en un agitador rotatorio (160 rpm) hasta alcanzar una UFC de 106-108 cel/mL. Se confeccionaron los inóculos correspondientes en sustrato sólido (turba) y se inocularon las semillas de forma convencional, según la metodología de Somasegaran y Hoben (1994).
El tamaño de las parcelas fue de 3 m x 15 m. Se utilizó una dosis de siembra de 15 kg/ha y se sembró a chorrillo espaciado, con una distancia entre surcos de 0,75 m. El peso seco se determinó con un marco de 1 m², en cuatro muestras dentro de la parcela.
Las mediciones se realizaron en el segundo corte, durante la fase de floración e inicio de la aparición de las vainas, para garantizar la máxima expresión agronómica del cultivo después del establecimiento.
El diseño experimental fue de bloques completamente aleatorizados. Se utilizaron 13 tratamientos: 11 de ellos inoculados con cepas aisladas de Centrosema spp., un testigo inoculado con la cepa de referencia 5030 y un control absoluto (tratamiento no inoculado ni fertilizado), con cuatro réplicas. Se aplicó un análisis de varianza (ANOVA) para experimentos de inoculación. Las variables evaluadas fueron: peso seco aéreo, PSA (g/parcela), índice de nodulación (IN) y rendimiento de nitrógeno, RN (mg/parcela). Se calculó el índice de efectividad de la inoculación, IEI (Davies et al., 2005), a partir del valor del PSA, para lo cual se utilizó la fórmula:
IEI: [(tratamiento inoculado control no inoculado) / control no inoculado] x 100
Se consideró un único factor (cepas) y su efecto sobre la leguminosa en comparación con el control no inoculado, sin la utilización del control fertilizado (Danielle Prévost, comunicación personal). Las diferencias entre medias se determinaron por la dócima de comparación de Duncan (1955). Se utilizó el paquete estadístico SPSS 8.0 para Windows. Para determinar el carácter de dependencia de la variable RN con respecto a IN, así como de RN con IN y de PSA con RN, se usó un coeficiente de regresión lineal simple (Ostle, 1984).
El IN se calculó de acuerdo con los parámetros que se muestran en la tabla 4 (Danielle Prévost, comunicación personal) y los datos fueron transformados por Öx (Ostle, 1984).
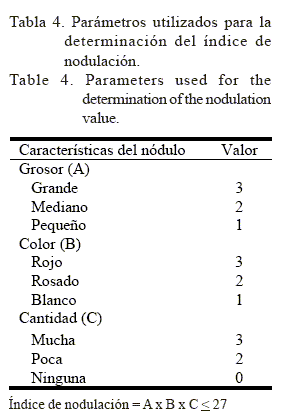
El nitrógeno total se calculó a partir de su digestión en ácido sulfúrico y la aplicación del método microanalítico de Kjeldahl. Con esos datos se calculó el rendimiento de nitrógeno (RN), mediante la multiplicación del porcentaje de N por el PSA de la planta.
RESULTADOS Y DISCUSIÓN
El marcado déficit de nutrientes en el suelo propició una mayor confiabilidad de los resultados experimentales, ya que no era de esperar una interferencia significativa por alta disponibilidad de macroelementos que pudiera enmascarar el efecto positivo de las cepas, sobre la base de sus propiedades como organismos fijadores del nitrógeno atmosférico.
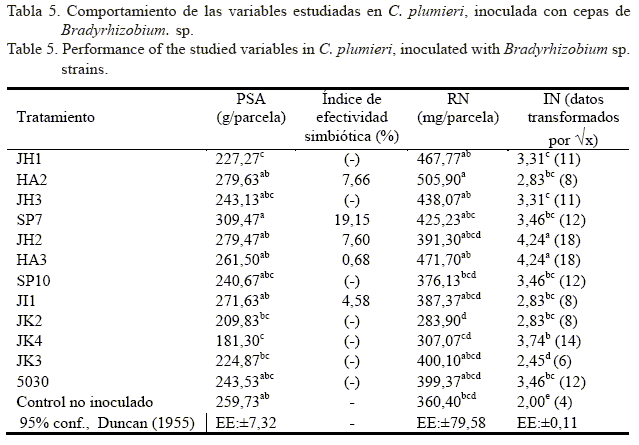
En la variable PSA (tabla 5) no se observó una respuesta efectiva de la inoculación en la planta. Ninguno de los valores de los tratamientos inoculados superó estadísticamente (p<0,05) el control no inoculado (259,73 g/parcela). El inoculado con la cepa nativa JK4 (181,30 g/parcela) fue estadísticamente inferior con respecto al control no inoculado. No hubo diferencias entre los tratamientos inoculados con las cepas nativas y el inoculado con la cepa comercial 5030 (243,53 g/parcela). Similares resultados encontró Date (1991) al inocular Macroptilium atropurpureum cv Siratro con cepas de Bradyrhizobium spp., seleccionadas previamente. Relacionado con esta variable, los índices de efectividad simbiótica (tabla 5) tuvieron valores muy bajos; el tratamiento inoculado con la cepa SP7 mostró el mayor valor (19,15%), y JI1, el menor (4,58%); así mismo, hubo tratamientos con valores negativos (JH1, JH3, SP10, JK2, JK4, JK3, 5030). Esta fórmula corroboró la débil efectividad simbiótica de las cepas inoculadas con respecto al control no inoculado, explicada anteriormente en los valores del PSA.
Por otra parte, en la variable RN (tabla 5) solo el tratamiento inoculado con la cepa nativa HA2 (505,90 mg/parcela) resultó estadísticamente superior al control no inoculado (360 mg/parcela), así como a los inoculados con las cepas nativas SP10 (376,13 mg/parcela), JK2 (283,90 mg/parcela) y JK4 (307,07 mg/parcela). Al igual que en la variable PSA, de estos resultados se podría inferir que la población natural de rizobios en la rizosfera del control no inoculado ejerció un efecto superior en la planta en cuanto a la fijación del dinitrógeno atmosférico, aunque ello no implicara, necesariamente, una mayor capacidad de infección radicular de dichas cepas. Tampoco se descarta que la planta haya utilizado de forma más eficiente el N mineral en el control no inoculado por la ausencia de rizobios autóctonos efectivos; como se observó en la variable IN, dicho control mostró los peores índices de infección radical.
En este sentido, Collins et al. (2009) y Arresé-Igor et al. (2009) aseguraron que el estrés climático posterior a la siembra afecta el proceso de fijación biológica del nitrógeno, con la inactivación de la nitrogenasa como primera respuesta al déficit hídrico creciente y, por lo tanto, se afecta la actividad nodular de forma previa a la fotosíntesis. Como se muestra en la tabla 2, en los meses posteriores a la siembra de la leguminosa hubo una depresión notable de las precipitaciones en el área experimental (diciembre: 34,8 mm; enero: 45,7 mm), por lo que no se descarta que este fuera el motivo por el cual las cepas que se inocularon no pudieran expresar al máximo su potencial de fijación del dinitrógeno atmosférico. Por otra parte, según Nápoles et al. (2008) la eficiencia simbiótica puede estar basada no solo en los factores edafoclimáticos, sino también en los factores de nodulación nod, los cuales pueden diferir entre las cepas.
No se observó, sin embargo, que el bajo índice de nodulación implicara obligatoriamente el bajo rendimiento de nitrógeno en el tratamiento inoculado con la cepa HA2 (tabla 5). Gómez et al. (2009) observaron, en plantas de Cicer arietinum y de Vigna unguiculata, en condiciones controladas, que la deficiencia de P disminuyó significativamente la nodulación; mientras que la actividad nodular específica y la producción de raíces se incrementó en algunos tratamientos.
La variable IN (tabla 5) mostró que todos los tratamientos inoculados fueron estadísticamente superiores (p<0,05) al control no inoculado (2,00), lo cual constituye una evidencia de que la capacidad de infección radicular de las cepas inoculadas fue mayor que la de la población natural de rizobios del suelo experimental. Los inoculados con las cepas nativas JH2 (4,24) y HA3 (4,24) resultaron estadísticamente superiores al resto. Estos resultados apuntan, indudablemente, hacia una formación de nódulos estadísticamente superior (p<0,05) en las plantas inoculadas con las cepas que se utilizaron en el experimento; ello pudo deberse a que el suelo presentó tenores ligeros de acidez, y la mayor parte de las cepas aplicadas a C. plumieri combinan las características de ser tolerantes a pH ácido y poseer un alto potencial de fijación simbiótica (Bécquer, 2002). Sin embargo, en prospecciones de leguminosas realizadas en el Valle del Cauto, Cuba, Gómez et al. (2010) encontraron que tanto C. plumieri como el resto de las leguminosas colectadas presentaron altos índices de nodulación natural. Ello contradice, de cierta forma, lo expresado por Howieson et al. (2008) acerca de que la mayor causa del uso no óptimo de la fijación simbiótica del dinitrógeno en la agricultura es la competencia de las cepas introducidas con las autóctonas, ya que estas últimas tienden a tener muy alta capacidad infectiva, pero baja capacidad de fijación. Por otra parte, los altos índices de nodulación presentados por las cepas de la zona central no siempre implicaron mayores valores en PSA y RN, debido posiblemente a un desbalance del intercambio de nutrientes entre los simbiontes. Al respecto, Martínez-Romero y Palacios (1990) consideraron que la capacidad de infección radicular de los rizobios, expresada por los genes comunes nod, no siempre implica una alta capacidad simbiótica, que a su vez constituye la expresión de otros genes, como los nif y los fix.
Estos resultados se corroboraron con los análisis de correlación y de regresión. En los tratamientos inoculados, el grado de dependencia y la correlación de la variable RN con respecto al IN (tabla 6) fueron débiles (R2 = 0,04; r = 0,06), así como del PSA con respecto al IN (R2 = 0,26; r = 0,16). Es obvio que no existió una relación estrecha entre estas variables en las plantas hospederas, por lo que se asume que la relación bioquímico-fisiológica entre la capacidad de infección radicular y la de fijación del nitrógeno atmosférico pudo variar en las cepas estudiadas, debido quizás a factores externos como los edafoclimáticos o internos como los genéticos (Martínez-Romero et al., 1990). Fesenko et al. (1995) obtuvieron resultados similares en la correlación de estas variables al inocular Pisum sativum con Rhizobium leguminosarum.

Sin embargo, se halló un mayor grado de regresión y de correlación (R2 = 0,34; r = 0,59) entre el RN y el PSA (tabla 7). Estos resultados, sobre todo los de la regresión, no se deben considerar altos desde el punto de vista estadístico (Antonio Sigarroa, comunicación personal), pero según la opinión de los autores del presente trabajo tienden a corroborar lo observado por Bordeleau et al. (1977) en Medicago sativa; en esta especie existió una alta correlación entre el rendimiento de nitrógeno y el peso seco de la planta, por lo que ellos consideran, como mejor criterio de selección de las cepas, los resultados en el peso seco. Otros autores, como Hume et al. (1985) y Prévost (1987), informaron que el rendimiento de nitrógeno en Onobrychis viciifolia estuvo altamente correlacionado con el crecimiento de la planta. Este fenómeno fue valorado, además, por Provorov y Tikhonovich (2003), quienes afirmaron que la intensidad de la fijación del dinitrógeno está correlacionada con el proceso energético de la planta, como lo es la actividad fotosintética, la cual provee no solo la energía necesaria para la reacción de la nitrogenasa, sino también las estructuras de carbono para la asimilación de sus productos.

A partir de los resultados, se concluye que las cepas nativas inoculadas en C. plumieri no presentaron, en general, una alta eficiencia simbiótica en comparación con el control no inoculado. Estas cepas, con la excepción de HA2 en el RN, así como JH2 y HA3 en el IN, no ejercieron un efecto altamente positivo en la planta, debido quizás a la intervención de factores climáticos adversos, como el estrés hídrico postsiembra. No obstante, dichas cepas poseen un buen potencial infectivo, lo cual es altamente valorado como factor de competitividad en la rizosfera. Se observó una interrelación débil entre las variables RN y PSA con respecto a IN, así como una interrelación más alta entre la variable RN con respecto a PSA.
Estos resultados se deben tener en cuenta para la continuación de estudios relacionados con la simbiosis leguminosa-rizobio.
REFERENCIAS BIBLIOGRÁFICAS
1. Anon. Clasificación genética de los suelos de Cuba. Academia de Ciencias de Cuba. Instituto de Suelos. La Habana, Cuba. 1979
2. Arresé-Igor, C. et al. Aspectos fisiológicos de la respuesta a sequía en leguminosas. Resúmenes XXIV RELAR. La Habana, Cuba. p. 38. 2009
3. Bécquer, C.J. Caracterización y selección de rizobios, aislados de leguminosas nativas de Sancti Spiritus, Cuba. Tesis en opción al grado científico de Doctor en Ciencias Biológicas. Universidad de La Habana, Cuba. 140 p. 2002
4. Bordeleau, L.M. et al. Effects des souches de Rhizobium meliloti et des coupes succesives de la Luzerne (Medicago sativa) sur la fixation symbiotique d'azote. Can. J. Plant Sci. 57:433. 1977
5. Collins, D. et al. Importancia de la FBN en cultivos y principales factores ambientales que la condicionan. Resúmenes XXIV RELAR. La Habana, Cuba. p. 36. 2009
6. Date, R.A. Nodulation success and persistence of recommended inoculum strains for subtropical and tropical forage legumes in northern Australia. Soil Biol. Biochem. 23:533. 1991
7. Davies, F.T. et al. Influence of arbuscular mycorrhizae indigenous to Peru and a flavonoid on growth, yield and leaf elemental of «Yungay» potatoes. Hort. Science. 40 (2):381. 2005
8. Fesenko, A.N. et al. Selection of Rhizobium leguminosarum bv. viceae strains for inculation of Pisum sativum L. cultivars: Analysis of symbiotic efficiency and nodulation competitiveness. Plant and Soil. 172:189. 1995
9. Funes, F. Los recursos fitogenéticos y la agroecología en Cuba. Memorias VII Taller Internacional sobre Recursos Fitogenéticos-FITOGEN. Sancti Spíritus, Cuba. p. 13. 2007
10. Gómez, I. et al. Leguminosas naturalizadas en el Valle del Cauto, Cuba. Pastos y Forrajes. 33:393. 2010
11. Gómez, L.A. et al. Variabilidad genotípica en la fijación simbiótica del nitrógeno en garbanzo y caupi: tolerancia a la deficiencia de fósforo y la salinidad. Resúmenes XXIV RELAR. La Habana, Cuba. p. 63. 2009
12. Hernández, A. et al. Nueva revisión de la clasificación genética de los suelos de Cuba. Instituto de Suelos, La Habana. 23 p. 1999
13. Hernández, Neice. Contribución al estudio de la regionalización de gramíneas en la provincia Sancti Spíritus. Tesis presentada en opción al grado científico de Dr. en Ciencias Agrícolas. Estación Experimental de Pastos y Forrajes Sancti Spíritus, Cuba. 134 p. 1989
14. Hernández, Neice et al. Leguminosas naturalizadas en las regiones ganaderas de Sancti-Spíritus. Pastos y Forrajes. 22:205. 1999
15. Howieson, J.G. et al. Prospects for the future use of legumes. En: Nitrogen-fixing leguminous symbioses. (Eds. M.J. Dilworth et al.). Springer, The Netherlands. p. 363. 2008
16. Hume, L.J. et al. Nitrogen fixation in sainfoin (Onobrychis viciifolia). Effectiveness of the nitrogen-fixing system. N. Z. J. Agric. Res. 28: 337. 1985
17. López, R. et al. Efectividad de cepas nativas de Rhizobium de un suelo Vertisol en la nodulación de leguminosas pratenses tropicales. Pastos y Forrajes. 25: 93. 2002
18. Martínez-Romero, Esperanza & Palacios, R. The Rhizobium genome. Cri. Rev. Plant Sci. 9:59. 1990
19. Nápoles, María Caridad et al. Influencia de algunos factores edáficos sobre la eficiencia de inoculantes en soya. Resúmenes Biotecnología'2008. La Habana. p. 368. 2008
20. Olivera, Yuseika et al. Colecta de leguminosas forrajeras en tres provincias orientales de Cuba. Pastos y Forrajes. 31:25. 2008
21. Ostle, B. Estadística aplicada. Editorial Científico Técnica, La Habana., Cuba. 629 p. 1984
22. Prévost, Danielle. Caractérisation des Rhizobium isolés de trois espèces de légumineuses arctiques: Astragalus alpinus L., Oxytropis maydelliana Trautv., et Oxytropis arctobia Bunge. (en francés). Thèse prèsenteé à l`ecole des gradués de l`Université Laval pour l`obtention du grade de Philosophiae Doctor (Ph.D.). Université Laval, Québec, Canada. 125 p. 1987
23. Provorov, N.A. & Tikhonovich, I.A. Genetic resources for improving nitrogen fixation in legume-rhizobia symbiosis. Genetic Resources and Crop Evolution. 50:89. 2003
24. Skerman, P. et al. Leguminosas forrajeras tropicales. FAO. Roma. 326 p. 1991
25. Somasegaran, P. & Hoben, H.J. Handbook for Rhizobia. Springer-Verlag, New York, USA. 450 p. 1994
26. Sylvester-Bradley, R. et al. Use of undisturbed soil cores for evaluation of Rhizobium strains and methods for inoculation of tropical forage legumes in a Colombia Oxisol. Plant Soil. 2: 237. 1983
27. Tang, M. Efecto de la nodulación natural en ocho leguminosas. Pastos y Forrajes. 19: 131. 1996
28. Tang, M. et al. Efecto de las cepas nativas de rizobios sobre varias leguminosas tropicales. Pastos y Forrajes. 17:45. 1994
29. Vance, C.P. et al. Biological nitrogen fixation: phosphorus-a critical need? En: Nitrogen fixation: from molecules to crop productivity. Proceedings of the 12th International Congress on Nitrogen Fixation. Foz do Iguaçu, Paraná, Brazil. p. 509. 2000
30. Vincent, J.M. A manual for the practical study of root nodules bacteria. IBP Handbook No. 15. Blackwell Scientific Publications, Oxford and Edinburgh, 164 p. 1970
Recibido el 7 de noviembre del 2011
Aceptado el 30 de mayo del 2012

{kind=link}
{kind=link}
{kind=link}