










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPastos y Forrajes
versión impresa ISSN 0864-0394
Pastos y Forrajes vol.37 no.2 Matanzas mar.-jun. 2014
ARTÍCULO DE INVESTIGACIÓN
Caracterización de la macrofauna edáfica en sistemas de producción intensiva de césped
Characterization of the edaphic macrofauna in intensive turfgrass production systems
Wendy Ramírez, Y. García, Saray Sánchez, Mislaidys López y L. Hernández
Estación Experimental de Pastos y Forrajes Indio Hatuey, Universidad de Matanzas Camilo Cienfuegos, Ministerio de Educación Superior Central España Republicana, CP 44280, Matanzas, Cuba
Correo electrónico: wendy.ramirez@ihatuey.cu
RESUMEN
Se realizó una investigación con el objetivo de evaluar el comportamiento de la macrofauna edáfica en áreas de la finca de producción intensiva de césped, en la Estación Experimental de Pastos y Forrajes Indio Hatuey. Se evaluaron tres sistemas, con diferentes años de explotación y tres réplicas por sistema: S1: suelos con diez años o más de explotación intensiva; S2: suelos con cuatro años de explotación; y S3: suelos de condición estándar o de referencia, en los que se establecerán las parcelas para la producción intensiva de césped. En sentido general, la composición taxonómica de la macrofauna edáfica fue similar en los sistemas evaluados, y existieron diferencias en cuanto a la diversidad y equitatividad de los órdenes. Los sistemas más perturbados (S1 y S2) fueron los de menor diversidad taxonómica, con cinco unidades taxonómicas (ut), debido al monocultivo continuado de gramíneas cespitosas. Los órdenes de mayor representación en el S1 fueron Coleoptera (38,63 %) y Haplotaxida (31,81 %); en el S2, Coleoptera (52,63 %) y Haplotaxida (28,94 %); y en el S3, Haplotaxida (34,48 %) y Lepidoptera (12,93 %). Estos resultados evidencian el potencial de la macrofauna edáfica como indicador de la calidad del suelo, y su sensibilidad ante las perturbaciones causadas por la intensidad en el uso y manejo del suelo.
Palabras clave: biota, manejo del suelo, gramíneas para céspedes.
ABSTRACT
A study was conducted in order to evaluate the performance of the edaphic macrofauna in areas of the intensive turfgrass production farm, of the Pasture and Forage Research Station Indio Hatuey. Three systems were evaluated, with different exploitation years and three replications per system: S1: soils with ten years or more of intensive exploitation; S2: soils with four years of exploitation; and S3: standard condition or reference soils, on which the plots for intensive turfgrass production will be established. In general, the taxonomic composition of the soil macrofauna was similar in the evaluated systems, and there were differences regarding diversity and equitability of the orders. The most disturbed systems (S1 and S2) were the ones with lower taxonomic diversity, with five taxonomic units (tu), due to the continuous monocrop of turfgrasses. The orders of higher representation in S1 were Coleoptera (38,63 %) and Haplotaxida (31,81 %); in S2, Coleoptera (52,63 %) and Haplotaxida (28,94 %) and in S3 the most outstanding were Haplotaxida (34,48 %) and Lepidoptera (12,93 %). These results show the potential of the edaphic macrofauna as indicator of the soil quality, and its sensitivity to the disturbances caused by the intensity in the soil use and management.
Key words: biota, soil management, turfgrasses.
INTRODUCCIÓN
La degradación de los suelos es una de las crisis más graves que enfrenta el planeta, por lo que se necesita estudiar los cambios ocurridos en el suelo en las escalas global, regional, nacional y local. La causa de la degradación de este recurso se relaciona con la acción directa o indirecta del hombre, quien rompe el equilibrio natural de los agroecosistemas, al intervenir en ellos. Esto ocurre mediante varios procesos físicos, químicos y biológicos, tales como: la compactación, la erosión, la acidificación, la salinización, la lixiviación, la escorrentía, la reducción de la capacidad de intercambio catiónico y de la disponibilidad de nutrientes para las plantas y la disminución de la biodiversidad (Kumar y Kafle, 2009).
El análisis de la calidad del suelo permite detectar los cambios que ocurren en él, especialmente en la parte biológica; además, provee los aspectos básicos para evaluar la sostenibilidad del manejo del sistema y tiene relación directa con la producción sostenible. Por tales razones, la calidad del suelo es el indicador primario del manejo de este y se considera un componente crítico de la agricultura sostenible (Karlen et al., 1997). En este contexto surgieron los indicadores de la calidad del suelo, los que permiten evaluar su estado y monitorear la estabilidad de los agroecosistemas, a partir de la caracterización de la macrofauna edáfica.
La macrofauna incluye los invertebrados mayores de 2 mm de diámetro y tiene un papel imprescindible en la productividad del suelo, debido a su capacidad de alterar el ambiente superficial y edáfico en el que se desarrollan las plantas (Lavelle et al., 2006). Además, los macroinvertebrados intervienen en los procesos de infiltración, aireación e incorporación de la materia orgánica en el suelo (Huerta et al., 2008).
Por tales motivos, la macrofauna es considerada un indicador biológico sensible de los impactos del uso del suelo en la calidad edáfica (Rousseau et al., 2013). La composición de las comunidades de la macrofauna, así como su abundancia, son indicadores de la biodiversidad y la intensidad de las actividades biológicas (Velásquez et al., 2007).
Cuando las prácticas de manejo y cosecha del césped son inadecuadas, se reducen los servicios que este brinda a la sociedad. Además, su impacto en los recursos naturales y en el medioambiente puede ser perjudicial y costoso (Strandberg et al., 2012), por lo que es necesario evaluar la calidad de los suelos destinados a la producción de césped, para proponer estrategias encaminadas al uso y manejo sostenible de este recurso.
Teniendo en cuenta lo antes expuesto, el objetivo de esta investigación fue caracterizar las comunidades de la macrofauna edáfica en sistemas de producción intensiva de césped.
MATERIALES Y MÉTODOS
Ubicación geográfica. El estudio se realizó en áreas de la finca de césped de la Estación Experimental de Pastos y Forrajes Indio Hatuey, ubicada entre los 22°48'7" de latitud Norte y los 81°2' de longitud Oeste, a 19,01msnm, en el municipio de Perico, provincia de Matanzas, Cuba (Academia de Ciencias de Cuba, 1989).
Condiciones edafoclimáticas
Suelo. El suelo donde se realizó la fase experimental se clasifica como Ferralítico Rojo lixiviado (Hernández et al., 1999) y de topografía plana.
Clima. En los últimos cinco años, la temperatura promedio anual de la zona fue de 24,38 ºC. Los meses más cálidos fueron julio y agosto, con temperaturas de 26,6 ºC y 26,7 ºC, respectivamente; y enero, el más frío (20,3 ºC). La humedad relativa promedio anual fue de 80,6 %, con el mayor valor en octubre (86,4 %) y el menor en abril (72,4 %).
Escenarios de la investigación
Para la selección de las áreas se tuvo en cuenta la caracterización general de cada sistema en estudio, las cuales tienen en común su historial en la producción intensiva de césped. El criterio de selección fue el tiempo de explotación de los suelos para la producción intensiva de gramíneas cespitosas.
Descripción de las áreas en estudio
Sistema 1 (S1): suelos con diez años o más de explotación intensiva para la producción de césped (4 ha). Se estudiaron tres parcelas, destinadas a la producción de tepes de las variedades bermuda 328 y 419.
Sistema 2 (S2): suelos con cuatro años de explotación intensiva para la producción de césped (5 ha). Se evaluaron tres parcelas dedicadas a la producción de tepes de las especies Stenothaphrum secundatum, Zoysia matrella y el híbrido de Cynodon dactylon x C. transvaalensis (bermuda 419).
Sistema 3 (S3): suelos de referencia, donde se establecerán nuevas parcelas para la producción intensiva de césped. Esta área (9 ha) se dividió en tres parcelas (de 3 ha cada una) para realizar el estudio, y fue considerada de referencia ya que no se ha utilizado para tal fin. Antes de la presente investigación era un sistema silvopastoril.
Procedimiento experimental
Muestreo. Los muestreos de la macrofauna se realizaron entre las 7:00 y 9:00 a. m., en sitios representativos de cada parcela, a partir de transeptos cuyo punto de origen y dirección se determinó al azar. Para el estudio se excavaron diez monolitos de 25 x 25 x 20 cm, en cada área, los que se dividieron en dos estratos: 0-10 y 10-20 cm; cada monolito estuvo separado por un intervalo de 5 m.
Los muestreos se realizaron en el mes de octubre, al final de la época lluviosa del año 2011, según lo recomendado por Lavelle et al. (2003), y de acuerdo con la metodología del Programa de Investigación Internacional «Biología y Fertilidad del Suelo Tropical» (Anderson e Ingram, 1993; Lavelle et al., 2003). Las lombrices se conservaron en formaldehÍdo al 4 %, y los invertebrados restantes, en alcohol al 70 %, para su posterior identificación en el laboratorio.
La macrofauna fue separada y contada de forma manual, y se identificó hasta el nivel taxonómico de orden mediante el empleo de las claves de Ruiz et al. (2008). Se determinaron los valores promedio de densidad (ind/m2) y biomasa (g/m2) para la comunidad edáfica, para cada taxón y por estrato, en cada parcela en estudio. La densidad se determinó en función del número de individuos, y la biomasa, a partir del peso húmedo en la solución preservante, con la utilización de una balanza analítica.
Diseño y análisis estadístico
El diseño fue completamente aleatorizado, para un total de tres réplicas por tratamiento. Los datos de la densidad y la biomasa de la macrofauna se analizaron mediante el programa InfoStat, versión libre. Se comprobó que estos cumplieran con los supuestos de homogeneidad de varianza (prueba de Levene, 1960) y de distribución normal (Shapiro Wilks, 1965); como no cumplían con dichos requisitos, se realizaron análisis no paramétricos. Para determinar las variaciones de la densidad y la biomasa de la macrofauna edáfica entre los sistemas, se empleó la prueba de Kruskal-Wallis.
RESULTADOS Y DISCUSIÓN
Composición y riqueza taxonómica de la macrofauna del suelo
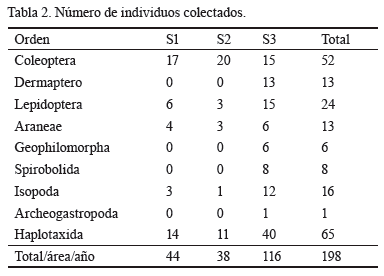
Las comunidades de la macrofauna del suelo en las áreas en estudio comprenden tres Phylum, siete clases y nueve órdenes. En la tabla 1 se muestra la composición taxonómica.
Se colectó un total de 198 individuos, en 90 monolitos realizados (tabla 2). Los órdenes más representados fueron Haplotaxida (32,8 %) y Coleoptera (26,2 %), y a continuación Lepidoptera (12,1 %); mientras que el resto de los órdenes no sobrepasó el 10 %.
El análisis de la riqueza taxonómica, que incluyó los grupos de la macrofauna con más alta frecuencia de aparición en este estudio, identificó el mayor número de órdenes (9) en el sistema de referencia (S3), mientras que en S1 y S2 se identificaron cinco para cada uno. Este mayor número de órdenes en el S3 se relaciona con la permanencia de cobertura en el suelo, motivado por el aporte de hojarasca de los árboles, lo que proporciona sombra y permite que se mantengan estables los valores de temperatura y humedad del suelo, y esto beneficia el desarrollo óptimo de las comunidades de la biota edáfica más diversas (Cabrera et al., 2011a). Por el contrario, en los sistemas en explotación el suelo se mantiene descubierto gran parte del tiempo, lo que incide en el aumento de su temperatura, así como en la disminución de la humedad. Estos factores atentan contra el desarrollo de las comunidades de la macrofauna (Zerbino et al., 2008).
En S1 y S2, la riqueza taxonómica (5) presente coincidió con el número de órdenes obtenidos por Sánchez y Milera (2002) en un pastizal con monocultivo de gramíneas. Sin embargo, fueron inferiores a los alcanzados por Pashanasi (2001) (23 taxones), y por Cabrera et al. (2011b), quienes encontraron 14 órdenes en pastizales y 12 en sistemas de cultivos varios. La diferencia en cuanto al número de órdenes informado por estos autores quizá se deba a la intensidad con que fueron explotados estos sistemas. Según Decaëns (2010), los bajos valores de riqueza se deben a que el número de especies decrece cuando el uso de la tierra se intensifica.
Asimismo, Sánchez y Reyes (2003) obtuvieron hasta nueve órdenes de la macrofauna en pastizales con diferente manejo ganadero, lo que coincide con la riqueza taxonómica encontrada en el área de referencia (S3), que contó con entrada de materia orgánica al igual que el pastizal a través de las bostas de los animales. Por ello, es posible que la coincidencia en la riqueza se relacione con la presencia de animales en el sistema.
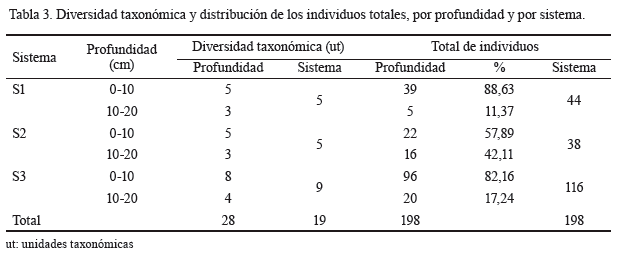
En la tabla 3 se muestran la diversidad taxonómica y el porcentaje de individuos totales, según la profundidad, en cada uno de los sistemas.
La mayor diversidad taxonómica correspondió al área de referencia, con nueve unidades taxonómicas (ut). En general, los valores de los tres sistemas son considerados bajos, lo que evidencia el grado de perturbación antropogénica presente en las áreas (Ramírez, 2013).
La existencia de solo 5 ut en los sistemas 1 y 2 indica que el monocultivo continuado de gramíneas cespitosas afectó la diversidad taxonómica de la macrofauna. La escasa diversidad de órdenes presentes en el área se debe a que el uso intensivo del suelo hizo que disminuyeran las comunidades de organismos, como consecuencia del efecto tóxico de los agroquímicos, la destrucción física de los hábitats y la reducción del alimento y de la materia orgánica del suelo (Ruiz et al., 2008).
Es evidente el efecto de la profundidad en la composición de la macrofauna y en la distribución de los individuos colectados. En la profundidad de 0-10 existió mayor diversidad taxonómica y los porcentajes de individuos colectados también fueron superiores. Estos resultados coinciden con los obtenidos por Crespo y Rodríguez (2000), quienes señalan que en la capa superficial del suelo se encuentra una significativa cantidad de individuos y se produce la mayor actividad de la macrofauna.
Densidad y biomasa
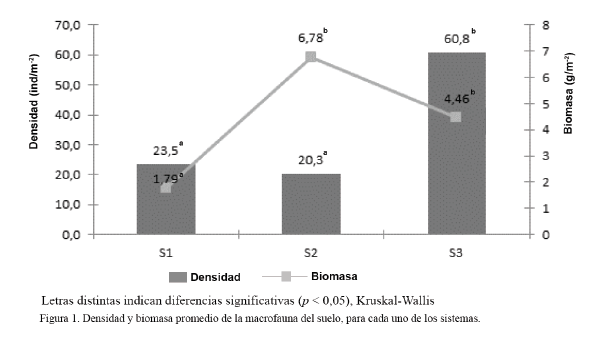
Según el análisis estadístico no paramétrico de Kruskal-Wallis (H = 26,57, p < 0,001), hubo diferencias altamente significativas entre la densidad del S3 y el resto de los sistemas; mientras que el S1 no difirió del S2 (fig. 1). Estos resultados demuestran el efecto negativo de la cosecha intensiva de tepes en las comunidades de la macrofauna edáfica.
La densidad de la macrofauna edáfica es una variable sensible ante el uso y manejo del suelo (Cabrera, 2012); los valores de densidad determinados en S1 y S2 fueron inferiores a los encontrados por Sánchez (2007), quien informó una densidad de 88,87 ind/m-2 en sistemas de pastoreo.Cuando se cosecha el tepe, el suelo se queda parte del tiempo desnudo, por lo que el aporte de la biomasa aérea es nulo y el de la fitomasa subterránea, escaso. Esto incide en que la disponibilidad de nutrientes sea insuficiente para el desarrollo de la macrofauna edáfica, y provoca el incremento de la temperatura y la pérdida de humedad en el suelo, las cuales constituyen condiciones inapropiadas para el desarrollo óptimo de los invertebrados (Szanser et al., 2011).
Por otra parte, la compactación del suelo en estos sistemas, debido a la entrada de equipos pesados y de maquinaria al campo (para las tareas de cosecha y transportación de los tepes), puede ser otro de los factores que afectan la densidad y la biomasa de la macrofauna. En este sentido, Lok (2005) señaló que la compactación de los suelos en los pastizales puede reducir la población de invertebrados edáficos.
Sin embargo, el S3 tuvo un menor grado de perturbación y presentó mejores condiciones para el funcionamiento de las comunidades de la macrofauna edáfica, ya que el estrato de una vegetación más densa contribuye a mantener estables los valores de temperatura y de humedad del suelo y, a la vez, aporta hojarasca. Estos resultados coinciden con los obtenidos por Zerbino (2008), quien plantea que la densidad es capaz de reflejar las diferencias de vegetación y manejo de los distintos usos del suelo.
En cuanto a la biomasa, según el análisis estadístico de Kruskal-Wallis (H = 11,79, p < 0,001) (fig. 1), existieron diferencias significativas entre el sistema con más de diez años de explotación en la producción intensiva de césped (S1) y el resto de los sistemas; mientras que el S2 no difirió del S3.
En el S2 se obtuvo 6,78 g/m2, valor similar al informado por Sánchez (2007) en pastizales: 6,51 g/m2; e inferior al reportado por Cabrera et al. (2011a): 7,71 g/m2.
A pesar de que el sistema con cuatro años de producción intensiva de césped (S2) tuvo una densidad similar al S1, presentó valores de biomasa muy superiores, ya que tenía un mayor tiempo de recuperación desde la última cosecha de tepes, lo cual incidió en que los individuos adquiriesen un mayor peso. Además, entre los órdenes de la macrofauna encontrados, las larvas de coleópteros y las lombrices fueron las de mayor peso en la biomasa, y también las de mayor número de individuos, por lo que tuvieron una gran influencia en los valores de la biomasa total. Estos resultados coinciden con los de Baretta et al. (2010), quienes encontraron individuos con mayor peso en los sistemas menos perturbados.
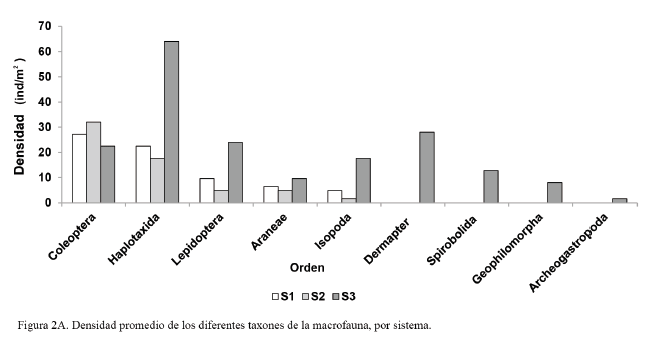
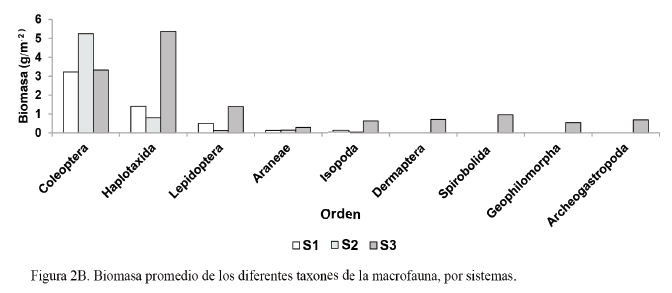
En la figura 2A y 2B se muestra el comportamiento de la densidad y la biomasa de los diferentes grupos taxonómicos de la macrofauna, en las áreas en estudio.
En cuanto a la densidad (fig. 2A), los órdenes de mayor representación fueron Coleoptera (27,2 ind/m2) y Haplotaxida (22,4 ind/m2) en el S1; en el S2 el comportamiento fue similar: Coleoptera (32 ind/m2) y Haplotaxida (17,6 ind/m2); mientras que en el S3, Haplotaxida (64 ind/m2) resultó ser el orden más representado, seguido por Lepidóptera (24 ind/m2).
Por otra parte, los órdenes que más aportaron a la biomasa total (fig. 2B) fueron Coleoptera (3,22 g/m2) y Haplotaxida (1,41 g/m2), en el S1; Coleoptera (5,25 g/m2) y Haplotaxida (0,79 g/m2), en el S2; y Haplotaxida (5,37 g/m2) y Coleoptera (3,32 g/m2), en el S3. A pesar de que la clasificación taxonómica no llegó hasta el nivel de familia y especie, los altos valores de biomasa del orden Coleoptera se deben, presumiblemente, a la abundancia de larvas Scarabaeidae, lo que coincide con lo informado por Cabrera et al. (2011a), quienes señalan que las larvas de Scarabaeidae predominan en lugares donde se desarrolla un buen sistema radicular, especialmente en los pastizales.
En los sistemas destinados a la producción intensiva de césped (S1 y S2) coincidieron los órdenes presentes, debido a la similitud en el manejo, por lo que colonizaron individuos de la macrofauna con la capacidad de adaptarse y resistir a las alteraciones a las que es sometido el medio edáfico.
En sentido general, en las áreas destinadas a la producción intensiva de césped, Coleoptera fue el taxón que más se destacó, con altos valores de densidad y de biomasa. Estos resultados coinciden con los de Cabrera et al. (2011b), quienes informaron que los individuos del orden Coleoptera sobresalen por sus altos valores en ambos indicadores.
Mientras, en el área de referencia (S3) se destacó el orden Haplotaxida en cuanto a la densidad y la biomasa. Feijoo et al. (2007) señalan la importancia de los individuos de este orden, debido a su amplia distribución y gran talla.
Los resultados de la densidad y la biomasa evidenciaron la sensibilidad de las comunidades de la macrofauna edáfica ante el uso y la intensidad con que se maneja el suelo, en los sistemas de producción intensiva de césped.
La densidad de la macrofauna edáfica no constituyó un indicador sensible en los sistemas con una composición florística similar, a pesar de que fueron manejados con distinta intensidad (S1 y S2); sin embargo, sí lo fue en el sistema donde hubo un contraste en cuanto a la diversidad de la vegetación (S3).
De forma general, el estudio de la macrofauna edáfica reflejó el grado de deterioro del suelo que se destina a la producción intensiva de gramíneas cespitosas, lo cual es ocasionado, en gran medida, por las prácticas de manejo inadecuadas y la intensidad en el uso y manejo de este, por lo que se revela su potencialidad para ser usada como un indicador que permite detectar las alteraciones que ocurren en el suelo.
REFERENCIAS BIBLIOGRÁFICAS
1. Academia de Ciencias de Cuba. Nuevo Atlas Nacional de Cuba. La Habana: Instituto Cubano de Geodesia y Cartografía. p. 41, 1989.
2. Anderson, J. M. & Ingram, J. S. I. (Eds.). Tropical soil biology and fertility: a handbook of methods. 2nd ed. Wallingford, United Kingdom: CAB International, 1993.
3. Baretta, D.; Gardner, G. B. & Nogueira, E. J. B. Potencial da macrofauna e outras variáveis edáficas como indicadores da qualidade do solo em áreas com Araucaria angustifolia. Acta Zool. Mex. 26 (spe2):135-150, 2010.
4. Cabrera, Grisel. La macrofauna edáfica como indicador biológico del estado de conservación/perturbación del suelo. Resultados obtenidos en Cuba. Pastos y Forrajes. 35 (4): 349-364. 2012.
5. Cabrera, Grisel; Robaina, Nayla & Ponce de León, D. Riqueza y abundancia de la macrofauna edáfica en cuatro usos de la tierra en las provincias de Artemisa y Mayabeque, Cuba. Pastos y Forrajes. 34 (3):313-330. 2011a.
6. Cabrera, Grisel; Robaina, Nayla & Ponce de León, D. Composición funcional de la macrofauna edáfica en cuatro usos de la tierra en las provincias de Artemisa y Mayabeque, Cuba. Pastos y Forrajes. 34 (3):331-346. 2011b.
7. Crespo, G. & Rodríguez, Idalmis. Contribución al conocimiento del reciclaje de los nutrientes en el sistema suelo-pasto-animal en Cuba. La Habana: EDICA, 2000.
8. Decaëns, T. Macroecological patterns in soil communities. Global Ecol. Biogeogr. 19 (3):287-302, 2010.
9. Feijoo, A.; Zúñiga, M. C.; Quintero, H. & Lavelle, P. Relaciones entre el uso de la tierra y las comunidades de lombrices en la cuenca del río La Vieja, Colombia. Pastos y Forrajes. 30:235. 2007.
10. Hernández, A.; Pérez, J. M.; Bosch, D. & Rivero, L. Nueva versión de la clasificación genética de los suelos de Cuba. La Habana: AGRINFOR, 1999.
11. Huerta, Esperanza; Rodríguez, Jannet; Castillo, Isabel E.; Montejo, E.; Cruz, Marcela de la & García, R. Relación entre la fertilidad del suelo y su población de macroinvertebrados. Terra Latinoamericana. 26 (2):171-181, 2008.
12. Karlen, D. L.; Mausbach, M. J.; Doran, J. W.; Cline, R. G.; Harris, R. F. & Schuman, G. E. Soil quality: A concept, definition, and framework for evaluation. Soil Sci. Soc. Amer. J. 61:4-10. 1997.
13. Kumar, A. & Kafle, N. Land degradation issues in Nepal and its management through agroforestery. J. Agr. Environ. 10:133-143. 2009.
14. Lavelle, P.; Decaëns, T.; Aubert, M.; Barot, S.; Blouin, M.; Bureau; et al. Soil invertebrates and ecosystem services. Eur. J. Soil Biol. 42:3-15, 2006
15. Lavelle, P.; Senapati, B. K. & Barros, E. Soil macrofauna. In: G. Schroth and F. L. Sinclair, eds. Trees, crops and soil fertility. Concepts and research methods. Bangor, United Kingdom: School of Agricultural and Forest Sciences, University of Wales. p. 303-317, 2003.
16. Levene, H. Robust test for equality of variance. In: I. Olkin, S. G. Ghurye, W. Hoeffing, W. G. Madow and H. B. Mann, eds. Contributions to probability and statistics: essays in honor of Harold Hotelling, California: Stanford University Press, p. 278-292, 1960.
17. Lok, Sandra. Determinación y selección de indicadores del sistema suelo-pasto en pastizales dedicados a la producción de ganado vacuno. Tesis presentada en opción al grado científico de Doctor en Ciencias Agrícolas. San José de las Lajas, Cuba: Instituto de Ciencia Animal, 2005.
18. Pashanasi, B. Estudio cuantitativo de la macrofauna del suelo en diferentes sistemas de uso de la tierra en la Amazonia Peruana. Folia Amazónica. 12 (1-2):75-93, 2001.
19. Ramírez, Wendy. Estudio de indicadores de la calidad del suelo en áreas destinadas a la producción intensiva de gramíneas cespitosas. Tesis en opción al título de Máster en Pastos y Forrajes. Matanzas, Cuba: EEPF Indio Hatuey, 2013.
20. Rousseau, L.; Fonte, S.; Téllez, O.; Hock, R.; & Lavelle, P. Soil macrofauna as indicators of soil quality and land use impacts in smallholder agroecosystems of western Nicaragua. Ecol Indic. 27:71-82, 2013.
21. Ruiz, Nuria; Lavelle, P. & Jiménez, J. Soil macrofauna field manual. Technical level. Rome: IRD, FAO, 2008.
22. Sánchez, Saray. Acumulación y descomposición de la hojarasca en un pastizal de Panicum maximum Jacq. y en un sistema silvopastoril de P. maximum y Leucaena leucocephala (Lam.) de Wit. Tesis presentada en opción al grado científico de Doctor en Ciencias Agrícolas. La Habana: Instituto de Ciencia Animal, 2007.
23. Sánchez, Saray & Milera, Milagros. Dinámica de la macrofauna edáfica en la sucesión de un sistema de manejo de gramíneas a un sistema con árboles intercalados en el pasto. Pastos y Forrajes. 25 (3):189-194, 2002.
24. Sánchez, Saray & Reyes, F. Estudio de la macrofauna edáfica en una asociación de Morus alba y leguminosas arbóreas. Pastos y Forrajes. 26 (4):315-320. 2003.
25. Shapiro S. S. & Wilk, M. B. An analysis of variance test for normality. Biometrika 52 (3/4):591-611. 1965.
26. Strandberg, M.; Blombäck, K.; Dahl, A. M. & Knox, W. Priorities for sustainable turfgrass management: a research and industry perspective. Acta Agr. Scand. B-S P. 62 (1):3-9, 2012.
27. Szanser, M.; Ilieva-Makulec, K.; Kajak, A.; Górska, E.; Kusiñska, A.; Kisielc, M. et al. Impact of litter species diversity on decomposition processes and communities. Soil Biol. Biochem. 43:9-19, 2011.
28. Velásquez, E.; Lavelle, P. & Andrade, M. GISQ, a multifuncional indicator of quality soil. Soil Biol. Biochem. 39:3066-3080, 2007.
29. Zerbino, S.; Altier, N.; Morón, A. & Rodríguez, C. Evaluación de la macrofauna del suelo en sistemas de producción en siembra directa y con pastoreo. Agrociencia. 12 (1):44-55, 2008.
Recibido el 28 de junio de 2013
Aceptado el 15 de marzo de 2014

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}