










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPastos y Forrajes
versión impresa ISSN 0864-0394
Pastos y Forrajes vol.37 no.3 Matanzas jul.-set. 2014
ARTÍCULO DE INVESTIGACIÓN
Efecto de diferentes usos de la tierra en la composición y la abundancia de la macrofauna edáfica, en la provincia de Matanzas
Effect of different land uses on the composition and abundance of the edaphic macrofauna, in Matanzas province
Y. García, Wendy Ramírez y Saray Sánchez
Estación Experimental de Pastos y Forrajes Indio Hatuey,
Universidad de Matanzas Camilo Cienfuegos, Ministerio de Educación Superior
Central España Republicana, CP 44280, Matanzas, Cuba
Correo electrónico: yoansy.garcia@ihatuey.cu
RESUMEN
Se evaluó el efecto de cuatro sistemas de uso de la tierra en la riqueza y la abundancia de la macrofauna edáfica, durante el periodo lluvioso (junio-octubre) del año 2011, en los municipios de Colón, Calimete y Perico (provincia de Matanzas, Cuba). El muestreo de la macrofauna se realizó según la Metodología del Programa de Investigación Internacional Biología y Fertilidad del Suelo Tropical (TSBF). Los sistemas evaluados fueron: cultivos varios (suelos con 10 años âo másâ de explotación intensiva), fincas agropecuarias (suelos con más de cinco años de explotación, con laboreo mínimo y con fertilización orgánica e inorgánica), silvopastoril (suelos con más de 10 años de explotación y con una carga global de ganado vacuno entre 1,7 y 1,8 UGM/ha) y pastizal (suelos con más de 10 años de explotación y con una carga global de ganado vacuno entre 1,3 y 1,5 UGM/ha). Los mayores valores de densidad y biomasa se obtuvieron en los sistemas menos perturbados, siguiendo el gradiente: silvopastoril-pastizal-fincas agropecuarias-cultivos varios. La densidad solo fue sensible en los sistemas donde hubo un contraste en cuanto a la diversidad y los estratos de vegetación. La biomasa reflejó los cambios en los diferentes usos y en el manejo de los suelos. Se demostró que los distintos usos de la tierra, así como el grado de perturbación e intensidad del manejo, influyeron en la composición y la abundancia de la macrofauna edáfica.
Palabras clave: Cultivos, pastizales, sistemas silvopascícolas.
ABSTRACT
The effect of four land use systems on the richness and abundance of the edaphic macrofauna, during the rainy season (June-October), 2011, was evaluated in the Colón, Calimete and Perico municipalities (Matanzas province, Cuba). The macrofauna sampling was performed according to the Methodology of the International Research Program «Tropical Soil Biology and Fertility» (TSBF). The evaluated systems were: staple crops (soils with 10 years âor moreâ of intensive exploitation), agricultural farms (soils with more than five years of exploitation, with minimum tillage and both organic and inorganic fertilization), silvopastoral (soils with more than 10 years of exploitation and a overall stocking rate of cattle between 1,7 and 1,8 LAU/ha) and pastureland (soils with more than 10 years of exploitation and with an overall stocking rate of cattle between1,3 and 1,5 LAU/ha). The highest density and biomass values were obtained in the least-disturbed systems, following the gradient: silvopastoral-pastureland-agricultural farms-staple crops. The density was sensitive only in the systems where there was a contrast regarding vegetation diversity and strata. The biomass reflected the changes in the different land uses and in soil management. It was proven that the different land uses, as well as the degree of disturbance and management intensity, influenced the composition and abundance of the edaphic macrofauna.
Key words: Crops, pastures, silvopastoral systems.
INTRODUCCIÓN
Varios estudios han sugerido que la mesofauna edáfica puede ser considerada como un indicador temprano y sensible a los efectos del manejo del suelo (Govaerts et al., 2006). En el trópico, la macrofauna es una de las más representativas dentro del suelo; esta contiene invertebrados con un diámetro mayor de 2 mm y fácilmente visibles en la superficie o en el interior, los cuales desempeñan un papel imprescindible en la productividad del suelo debido a su capacidad de alterar el ambiente superficial y edáfico en que se desarrollan las plantas (Lavelle et al., 2006).
La biodiversidad de dichos individuos declina como función inversa a la intensidad con que son cultivadas las plantas mediante métodos mecanizados y agroquímicos. El uso intensivo del suelo en la explotación agrícola hace que disminuyan las comunidades de los organismos como consecuencia del efecto tóxico de los agroquímicos, la destrucción física de los hábitats y la reducción del alimento y de la materia orgánica (Ruiz et al., 2008).
Según Coral y Bonilla (1998), el cambio de uso de la tierra provoca variación en las poblaciones edáficas como respuesta a las modificaciones en la cobertura vegetal, la radiación solar, la lluvia y las propiedades físicas y químicas del suelo. La pérdida de la materia orgánica constituye uno de los principales factores que influyen en los bajos valores de densidad y biomasa de la macrofauna (Velásquez et al., 2009); además esta se relaciona estrechamente con las propiedades químicas del suelo, tales como: el contenido de cationes intercambiables, el pH, la textura y la capacidad de retención de agua (Barros et al., 2002).
Esta investigación tuvo como objetivo evaluar el efecto de cuatro usos de la tierra en la composición y la abundancia de la macrofauna edáfica.
MATERIALES Y MÉTODOS
El estudio se realizó en el periodo lluvioso del año 2011 (junio-octubre), en los municipios de Colón, Calimete y Perico, pertenecientes a la provincia de Matanzas (Cuba). Se trabajaron cuatro usos de la tierra: cultivos varios, fincas agropecuarias, silvopastoril y pastizal. El tipo de suelo en todos los usos fue Ferralítico Rojo (Hernández et al., 1999), de topografía llana, con lo cual se garantizaron condiciones edáficas lo más homogéneas posible.
Descripción de los sitios evaluados
Cultivos varios
Se evaluaron tres sitios ubicados en los municipios de Calimete (Fregat San Pablo I: 22° 31ʹ 41,87ʺ N y 80° 53ʹ 43,17ʺ O; extensión máxima: 35,05 ha), Colón (Fregat 3: 22° 41ʹ 45,95ʺ N y 80° 53ʹ 15,65ʺ O; extensión máxima: 20,60 ha) y Perico (Fregat 7 Roque: 22° 45ʹ 55,03ʺ N y 81° 03ʹ 08,01ʺ O; extensión máxima: 23,46 ha). En estos predominaba la papa (Solanum tuberosum) como cultivo principal, en rotación con maíz (Zea mays), boniato (Hypomoea batata) y calabaza (Cucurbita maxima); y en la vegetación indeseable se halló: don Carlos (Sorghum halepense), zancaraña (Rotboellia cochinchinensis), lechosa (Euphorbia heterophyllia), guisaso de caballo (Xanthium strumarium), bledo (Amaranthus dubius Mart.) y cebolleta (Cyperus rotundus L.). Las áreas tenían más de 10 años de explotación con este uso de la tierra. El método de preparación del suelo fue el tradicional; al inicio se utilizó un sistema de riego por gravedad y aspersión (máquinas Fregat), y al final, por aspersión con sistema eléctrico. Además, se aplicó fórmula completa 9-13-17 (NPK), a razón de 670 kg/ha/año, así como fertilización nitrogenada con portador simple (urea): entre 223 y 298 kg/ha/año. La carga tóxica osciló desde 18,03 hasta 20,36 g (ingrediente activo) por hectárea en los últimos tres años, basado fundamentalmente en el uso de herbicidas (premergentes y posemergentes) y fungicidas.
Fincas agropecuarias
Se estudiaron tres sitios: uno ubicado en el municipio de Calimete (finca El Estabulado: 22° 27ʹ 41,34ʺ N y 80° 56ʹ 00,08ʺ O; extensión máxima: 40,76 ha) y dos en Perico (finca Santa Rosa: 22° 47ʹ 32,26ʺ N y 81° 02ʹ 23,95ʺ O; extensión máxima: 30 ha; finca El Campero: 22° 45´ 42,76ʺ N y 81° 01´ 27,80ʺ W; extensión máxima: 9,4 ha). Estos no presentaban un cultivo principal definido y estaban en constante rotación con maíz, frijol (Phaseolus vulgaris), pimiento-ají (Capsicum annum), tomate (Solanum lycopersicum), col (Brassica oleraceae), yuca (Manihot sculentum) y boniato. Las principales plantas indeseables eran: don Carlos, zancaraña, cebolleta, lechosa y metebravo (Echinocloa colona). Las áreas tenían más de cinco años de explotación con este uso de la tierra. La preparación del suelo se realizó mediante el método de laboreo mínimo, y la fertilización se basó principalmente en enmiendas orgánicas (compost y humus de lombriz), entre 10-20 t/ha, y pequeñas aplicaciones de fórmula completa y urea. El riego se realizó por gravedad y las aplicaciones de agroquímicos fueron escasas, por lo que la carga tóxica fue muy baja.
Sistema agroforestal
Se seleccionaron tres cuartones en silvopastoreo (cada uno con una hectárea de extensión) de la Estación Experimental de Pastos y Forrajes Indio Hatuey (EEPF-IH), en el municipio de Perico (22° 50´ 12,26ʺ N y 81° 02´ 25,99ʺ O). La composición florística era la siguiente: cuartón 5: Leucaena leucocephala (Lam.) cv. CNIA-250 y pastos naturales; cuartón 4: L. leucocephala (Lam.) cv. Perú y Megathyrsus maximus Jacq. (guinea); y cuartón 2: L. leucocephala cv. Cunningham y M. maximus. La carga global estuvo entre 1,7 y 1,8 UGM/ha. Las áreas tenían más de 10 años de explotación.
Pastizal
Se evaluaron tres sistemas del municipio de Perico: la finca El Campero (22° 45´ 42,76ʺ N y 81° 01´ 27,80ʺ O; con una extensión de 4,02 ha de pastos naturales y una carga global de 1,40 UGM/ha), la finca Palo Lindo (22° 53´ 37,53ʺ N y 81° 01´ 53,55ʺ O; con una extensión total de 87,23 ha de pastos naturales y 1,30 UGM/ha) y los cuartones 1 y 2 de Campo 12 de la EEPF-IH (22° 50´ 12,26ʺ N y 81° 02´ 25,99ʺ O; con una extensión de 3,2 ha y 1,5 UGM/ha). Este último era un pastizal naturalizado, compuesto por M. maximus Jacq., que llevaba seis meses de reposo. Las especies encontradas en los dos primeros sistemas fueron: alpargata (Paspalum notatum Flüegeé), pitilla villareña (Dichanthium annulatum (Forssk.) Stapf.), jiribilla, yerba de línea (Dichanthium caricosum (L.) A. Camus) y camagüeyana (Bothriochloa pertusa (L.) A. Camus). La cobertura de pasto en todos los sistemas estaba por encima de 90 % y el manejo de la masa vacuna se realizaba por el método extensivo. Todas las áreas tenían más de 10 años de explotación.
Características del muestreo y técnicas empleadas
El muestreo de la macrofauna se realizó según la Metodología del Programa de Investigación Internacional Biología y Fertilidad del Suelo Tropical (TSBF), que consiste en la extracción de monolitos de 25 x 25 x 30 cm en un transepto cuyo punto de origen se determina al azar y en dirección lineal. La recolecta se realizó manualmente in situ, hasta los 20 cm de profundidad, explorando los estratos de suelo 0-10 y 10-20 cm. Se tomaron 10 monolitos de suelo, separados en 5 m por sitio de muestreo. Se utilizaron tres réplicas por tratamiento (uso de suelo).
La determinación taxonómica del material se realizó hasta el nivel de orden, basado en los trabajos relacionados con la taxonomía de los grupos (Brusca y Brusca, 2003; Peck, 2005) y con la ayuda de especialistas de la Universidad Central de las Villas Martha Abreu (Villa Clara, Cuba).
Los órdenes Isoptera e Hymenoptera no se cuantificaron; solo se evaluó su presencia y ausencia, según los criterios de Ruiz-Cobo et al. (2010).
Las lombrices se preservaron en solución de formalina (4 %) y alcohol (70 %); el resto de la fauna se conservó en alcohol (75 %).
Se calcularon los valores promedios de densidad (ind m-2) y biomasa (g m-2) para las comunidades edáficas, en cada taxón. La densidad se determinó a partir del número de individuos, y la biomasa, sobre la base del peso húmedo en la solución preservante.
Para determinar la diferencia significativa entre los usos del suelo se realizó el test no paramétrico de Kruskal-Wallis y a posteriori el test U de Mann-Whitney, ya que no se cumplieron los supuestos de normalidad y homogeneidad de la varianza. El paquete estadístico utilizado fue SPSS® versión 17 para Windows® (2011) e InfoStat versión L (2007). Para todos los análisis se utilizó un nivel de significación de 0,05.
RESULTADOS Y DISCUSIÓN
Composición y riqueza taxonómica
Durante la investigación se realizaron 120 monolitos y se recolectaron 1 025 individuos: 73 en el uso de cultivos varios, 114 en las fincas agropecuarias, 653 en el sistema silvopastoril y 186 en el pastizal. Estos resultados son inferiores a los obtenidos por Velázquez et al. (2007), quienes encontraron 169 individuos en cultivos de maíz y 196 en pastizales; sin embargo, son superiores a los reportados por estos autores en bosques secundarios (366) de Colombia, en un suelo de origen volcánico y con un clima bimodal de alta precipitación (1 900 mm/año).
En el sistema silvopastoril se colectó una cantidad de individuos superior respecto a los otros usos del suelo. Los factores que intervinieron en esta marcada diferencia se explican con posterioridad en el acápite sobre densidad.
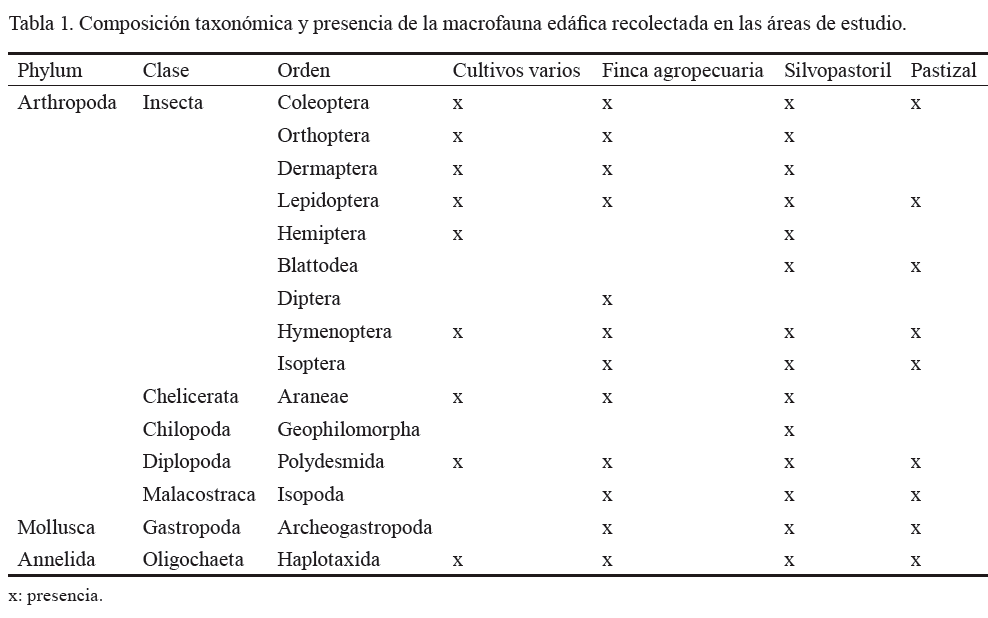
La composición taxonómica del experimento hasta el taxón orden se muestra en la tabla 1. Se encontraron individuos de tres Phylum, siete clases y 15 órdenes: 9 en el uso de cultivos varios, 12 en las fincas agropecuarias, 14 en el sistema silvopastoril y 9 en el pastizal.
Brown et al. (2001) obtuvieron un total de 16 grupos taxonómicos en los bosques y 14 en los pastizales, en ecosistemas mexicanos, lo cual supera los resultados alcanzados en la presente investigación. En estudios más recientes realizados en Cuba, Cabrera et al. (2011) hallaron 19 grupos taxonómicos en los bosques secundarios y 14 en los pastizales.
La riqueza tuvo una tendencia a disminuir, según el grado de disturbio físico y las diferencias en la vegetación (Zerbino, 2010); el sistema silvopastoril brindó las mejores condiciones de hábitat y de recursos para ser colonizado (Decäens et al., 2001).
El sistema de cultivos varios tuvo un efecto negativo en la mayoría de los grupos taxonómicos (con excepción de Coleoptera), debido al laboreo intensivo y a la baja disponibilidad de materia orgánica del suelo (Brown et al., 2001). También pudo influir el grado de descomposición de los restos vegetales, ya que en este uso del medio edáfico suele haber una reducción de la cantidad y la diversidad bioquímica de los exudados radiculares, así como de la hojarasca que ingresa al sistema (Joshi et al., 2004). Por otra parte, al comparar las fincas agropecuarias con los cultivos varios se observó que estas presentaron una mayor riqueza. Dicha diferencia quizás se debió al manejo, ya que en las fincas el suelo se preparó por laboreo mínimo, y además se aplicó materia orgánica y una cantidad baja de insecticida; contrariamente, en el otro sistema se realizó una preparación tradicional y un uso indiscriminado de agroquímicos, lo que elevó la carga tóxica.
Densidad
En cuanto a la densidad (fig. 1), según la prueba de Kruskal-Wallis (H = 54,42; p < 0,05; gl = 3), hubo diferencias significativas entre el sistema silvopastoril y el resto de los usos del suelo. El pastizal no difirió de las fincas agropecuarias y este último sistema, a su vez, no difirió de cultivos varios.
La densidad promedio fue de 60,8 ind m-2 en las fincas agropecuarias; 38,93 ind m-2, en cultivos varios; 347,73 ind m-2, en el sistema silvopastoril y 101,86 ind m-2, en el pastizal. Estos resultados son inferiores a los obtenidos por Zerbino (2007) y Cabrera et al. (2011) en usos intensivos del suelo: 294 y 137 ind m-2, respectivamente. Sin embargo, Ruiz et al. (2011), al estudiar varios usos de la tierra en cuatro sistemas de producción intensivos, encontraron valores desde 36,3 hasta 174,3 ind m-2.
Por otra parte, Cabrera et al. (2011) reportaron 1 166,60 ind m-2 en bosques secundarios y 581 ind m-2 en pastizales, los cuales son superiores a los obtenidos en el presente estudio. Asimismo, Zerbino (2010) encontró valores de 150 y 119 ind m-2 en pastizales, los que se asemejan a los hallados en esta investigación. Otros resultados similares fueron informados por Sánchez (2007) en pastizales de Cuba, así como por Velasquez et al. (2007) al comparar varios usos de la tierra: 577 ind m-2 en pastizales y 366 ind m-2 en bosques secundarios.
Los resultados mostraron la sensibilidad de la macrofauna edáfica ante diversos factores, como la cobertura vegetal, ya que en los cultivos varios y las fincas agropecuarias el muestreo se realizó en condiciones de cultivo limpio, solo con la planta que se estaba desarrollando (maíz), por lo que el aporte de biomasa aérea y radical de las plantas arvenses fue casi nulo. Tampoco hubo hojarasca y por ello la disponibilidad de elementos nutritivos para el desarrollo de la macrofauna edáfica fue muy escasa (Jean-Pierre y Blanchart, 2005).
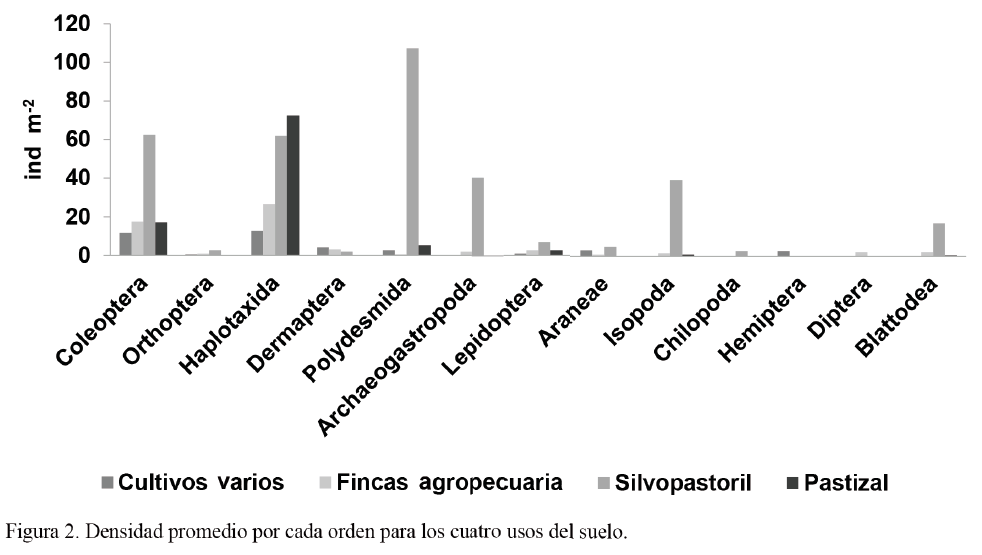
La densidad más elevada correspondió al orden Polydesmida, con 107 ind m-2 en el sistema silvopastoril (fig. 2). Los órdenes Haplotaxida y Coleoptera fueron los más abundantes para el resto de los sistemas evaluados (26,66 y 17,60 ind m-2 en las fincas agropecuarias; 12,80 y 11,73 ind m-2 en los cultivos varios; y 61,86 y 62,40 ind m-2 en el silvopastoril). Resultados similares fueron reportados por Cabrera et al. (2011) en las condiciones de Cuba. Por su parte, Zerbino (2007) obtuvo mayores valores de densidad para Oligochaeta (87,20 ind m-2), que fue la unidad taxonómica más abundante.
Asimismo Sevilla et al. (s. f.) reportaron, para lombrices de tierra, valores de 61 ind m-2 en bosques secundarios y 131 ind m-2 en pastos naturales, lo cual indica que estos últimos proveen un ambiente adecuado para el desarrollo de dichos individuos. En bosques secundarios, Cabrera et al. (2011) encontraron que Diplopoda, Isopoda e Isoptera fueron los órdenes más representativos en cuanto a densidad; mientras que en los pastizales se destacaron Hymenoptera (Formicidae), Isoptera y Coleoptera. Tales resultados son similares a los alcanzados en este trabajo, con la diferencia de que en la investigación anterior, realizada en las condiciones de Cuba y en el mismo tipo de suelo, los autores hallaron muy pocos individuos de lombrices.
El orden Polydesmida mostró los valores de densidad más elevados en el sistema silvopastoril respecto a los restantes usos del suelo, y ello coincide con lo informado por Sevilla et al. (s. f.). De manera similar, en bosques tropicales de Colombia, Cerón et al. (2008) hallaron Polydesmida, lombrices de tierra, Gastropoda y Coleoptera; mientras que en una pradera la mayor parte de la densidad estuvo representada por Oligochaeta y Coleoptera. Este resultado demuestra que Oligochaeta es un componente clave en la macrofauna de los sistemas en los que se desarrollan los pastos.
En este estudio se evidenció que Haplotaxida fue capaz de resistir en condiciones más adversas que el resto de los individuos, ya que aunque fue un componente importante de la densidad total del sistema silvopastoril, estuvo acompañada por otras unidades taxonómicas que no estuvieron representadas en los otros usos del suelo; mientras que las lombrices de tierra constituyeron casi la totalidad del número de individuos del resto de los sistemas.
Los órdenes Archeogastropoda e Isopoda manifestaron cierta influencia en la densidad de los sistemas silvopastoriles; sin embargo, Cerón et al. (2008) reportaron bajos valores de Isopoda en bosques. Estos individuos prefieren, al igual que Diplopoda, hábitats que favorezcan el mantenimiento de la humedad, y en los que no incida la acción directa de los rayos del sol y haya hojarasca de calidad (Adis et al., 1996). En este sentido, Xiaodong y Chen (2009), al evaluar tres tipos de bosques (secundarios, de hoja ancha y selvas tropicales), encontraron diferencias significativas en el número de individuos, debido a la calidad de la hojarasca.
Biomasa
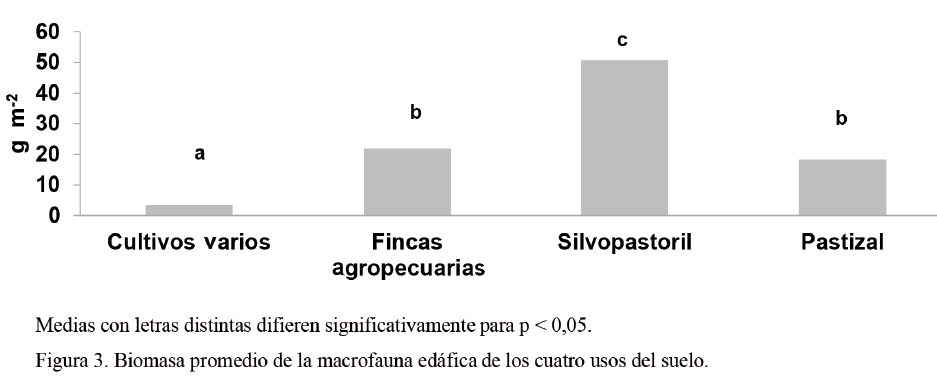
Respecto a la biomasa (fig. 3), según la prueba no paramétrica de Kruskal Wallis (H = 42,71; p < 0,05; gl = 3), se encontraron diferencias significativas entre el sistema silvopastoril (50,86 g m-2) y el resto de los usos del suelo; el pastizal (18,29 g m-2) no difirió de las fincas agropecuarias (21,84 g m-2), y los cultivos varios (3,46 g m-2) presentaron los valores más bajos. En este sentido, Cabrera et al. (2011) reportaron valores similares a los encontrados en los cultivos varios.
Los resultados obtenidos en el pastizal fueron superiores a los informados por Sánchez (2007) y por Cabrera et al. (2011), así como inferiores a los hallados por Rodríguez et al. (2002) en sistemas donde predominaba la yerba de guinea. Los valores de biomasa en el sistema silvopastoril se aproximan a los encontrados por Cabrera et al. (2011) en bosques secundarios (67,45 g m-2).
Las fincas agropecuarias, a pesar de que tuvieron una densidad muy similar a la de los cultivos varios, mostraron una biomasa superior. Esto se relaciona con que una sola o pocas unidades taxonómicas pueden tener una gran influencia en el contenido de biomasa total (Morales y Sarmiento, 2002). El hecho de que el suelo estuviese menos perturbado debido a la acción del hombre, así como menos contaminado por agroquímicos, al parecer incidió en que los individuos desarrollaran un mayor peso; sin embargo, diversos autores como Zerbino (2007) plantean que la biomasa no constituye un indicador fiable para detectar distintos usos del suelo.
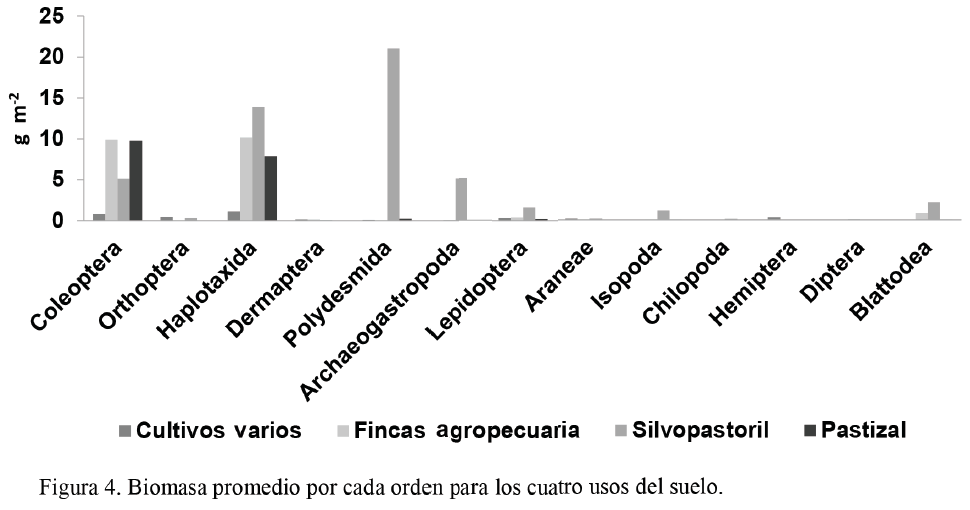
En la figura 4 se muestra la biomasa por cada orden encontrado. El valor de Haplotaxida (10,18 g m-2) en las fincas agropecuarias fue superior al de los cultivos varios (1,13 g m-2); algo similar sucedió con Coleoptera, ya que alcanzó 9,91 g m-2 en las fincas agropecuarias y 0,79 g m-2 en los cultivos varios. Esta tendencia coincide con los resultados obtenidos por Cerón et al. (2008).
En este sentido Birkas et al. (2010), al comparar varias formas de laboreo del suelo, encontraron bajos valores de lombrices de tierra en el sistema tradicional; mientras que en la siembra directa estos fueron más elevados, lo que demuestra que la intensidad de laboreo tiene una acción directa en la biomasa de la biota edáfica.
Además, la biomasa fue superior en el sistema silvopastoril, cuyo suelo fue el más conservado ya que es el que más se asemeja a los sistemas naturales âcomo las selvas o bosques tropicalesâ, y cuenta con una entrada constante y frecuente de materia orgánica de origen vegetal y animal (Huerta et al., 2008).
Teniendo en cuenta lo anterior, la biomasa es un factor importante para detectar cambios en el uso de la tierra y en el manejo, pero el análisis quizás no deba basarse en el número global ni en la media de la biomasa, sino en cada taxón evaluado; ya que en el sistema más idóneo los individuos se deben desarrollar mejor y, por consiguiente, deben alcanzar un mayor peso.
El orden Polydesmida mostró los mejores valores: 21,02 g m-2 en el sistema silvopastoril, en contraste con 0,21 g m-2 en el pastizal. Por otra parte, en el sistema silvopastoril el valor de Haplotaxida fue de 13,89 g m-2, superior al encontrado en el pastizal; sin embargo, el valor de densidad de este último fue ligeramente superior.
Así, en un sistema se encontró mayor número de lombrices mientras que en el otro estas estaban más pesadas, lo cual evidencia una mayor biomasa. Este resultado es similar al informado por Sevilla et al. (s. f.), quienes obtuvieron densidad de 50 ind m-2 y biomasa de 14,7 g m-2 en plantaciones en barbecho, mientras que en pastos naturales los valores fueron de 131 ind m-2 y 9,5 g m-2. El comportamiento de este orden se debió a las características especiales de sus individuos, los que pueden alcanzar diferentes tallas y pesos; de ello se infiere que una mayor densidad de lombrices no significa que exista una mayor biomasa ni mejores condiciones para su desarrollo. Por tanto, las lombrices se pueden considerar como un detector temprano de cualquier variación en las propiedades biológicas, químicas y físicas de los suelos, y, por ende, pueden ser un indicador de la calidad de este recurso (Paoletti, 1999; Nunes et al., 2007).
Al evaluar varios sistemas, Brown et al. (2001) encontraron que los que tenían una intensidad de laboreo y una escasa cantidad de materia orgánica presentaron valores más bajos (alrededor de 10 g m-2); así como que la mayor biomasa fue aportada por las lombrices de tierra, con un 81 al 96 % del total. Estos autores plantearon que con la intensificación del uso del suelo ocurre una pérdida parcial o total de la capa de hojarasca, así como cambios microclimáticos y transformaciones en las características físicas y químicas, debido a una mayor fotoxidación, lixiviación de los nutrientes y erosión, entre otras modificaciones. Estos cambios provocan la desaparición de los organismos que dependen de la hojarasca para sobrevivir y el desarrollo de organismos oportunistas invasores (algunas lombrices de tierra exóticas) o plagas invasoras (termitas y hormigas).
CONCLUSIONES
Los distintos usos del suelo repercutieron en la macrofauna edáfica, en dependencia del manejo y el grado de perturbación, ya que los sistemas menos perturbados alcanzaron los valores más altos de densidad y biomasa, siguiendo el gradiente: silvopastoril-pastizal-fincas agropecuarias-cultivos varios.
La densidad no es un indicador sensible en sistemas que tienen vegetación similar, aunque estén en diferentes usos del suelo; sin embargo, sí lo es en usos de la tierra donde haya un contraste en cuanto a la diversidad y los estratos de la vegetación.
La biomasa mostró las variaciones en los cuatro usos del suelo, así como en el manejo.
REFERENCIAS BIBLIOGRÁFICAS
1. Adis, J.; Golovatch, S.I. & Hamann, S. Survival strategy of the Terricolous millipede Cutervodesmus adisi Golovatch (Fuhrmannodesmidae, Polydesmida) in a blackwater inundation forest of Central Amazonia (Brazil) in response to the flood pulse. Acta Myriapodologica, Mém. Mus. Hist. Nat. 169:523, 1996.
2. Barros, E.; Pashanasi, B.; Constantino, R. & Lavelle, P. Effects of land-use systems on soil macrofauna in Western Amazon basin. Biol. Fertil. Soils. 35: 338, 2002.
3. Birkas, Marta; Bottlik, L.; Stingli, A.; Gyuricza, C. & Jolánkai, M. Effect of soil physical state on the earthworms in Hungary. Applied and Environmental Soil Science. [Disponible en:] http://www.hindawi.com/ journals/aess/2010/830853/, 2010. [20/5/2010]
4. Brown, G.; Fragoso, C.; Barois, I.; Rojas, P.; Patrón J.; Bueno, J.; Moreno, A.; Lavelle, P.; Ordaz, V. & Rodríguez, C. Diversidad y rol funcional de la macrofauna edáfica en los ecosistemas tropicales mexicanos. Acta Zoológica Mexicana (n.s.). Número especial. 1:79, 2001.
5. Brusca, R. & Brusca, G. Invertebrates. Sinauer Associates. Sunderland, Massachusetts, USA. Segunda edición. 966 p., 2003.
6. Cabrera, Grisel.; Robaina, Nayla & Ponce de León, D. Riqueza y abundancia de la macrofauna edáfica en cuatro usos de la tierra en las provincias de Artemisa y Mayabeque, Cuba. Pastos y Forrajes. 34:313, 2011.
7. Cerón, P.; Montenegro, S. & Noguera, E. Macrofauna en suelos de Bosque y Pajonal de la reserva natural Pueblo Viejo, Nariño, Colombia. Rev. Acad. Colomb. Cienc. 32 (125):447, 2008.
8. Coral, D.M. & Bonilla, C.R. Impacto de las prácticas agrícolas sobre la macrofauna del suelo en la cuenca alta del lago Guamues, Pasto, Colombia. Acta agronómica. 18 (3-4):55, 1998.
9. Decäens, T.; Lavelle P.; Jiménez J. J.; Escobar, G.; Rippstein, G.; Schneidmadl, J. et al. Impact of land management on soil macrofauna in the eastern plains of Colombia. Pp. 19-41. In: J. J. Jiménez and R. J. Thomas (Eds.). Natureâs plow: Soil macroinvertebrate communities in the Neotropical savannas of Colombia. CIAT Publication, Cali. No. 324., 2001.
10. Govaerts, B.; Sayre, K. D. & Deckers, J. A minimum data set for soil quality assessment of wheat and maize cropping in the highlands of Mexico. Soil & Tillage Research. 87: 163, 2006.
11. Hernández, A.; Ascanio, M. O.; Cabrera, A.; Morales, M.; Medina, N. &. Ribero, L. B.. Nueva versión de la clasificación genética de los suelos de Cuba. Ministerio de la Agricultura. La Habana, Cuba. 64 p., 1999.
12. Huerta, Esperanza; Rodríguez, Jannet; Evia, Isabel; Montejo, E.; Mondragón, Marcela & García, R.. Relación entre la fertilidad del suelo y su población de macroinvertebrados. Terra Latinoamericana. 26 (2):171, 2008.
13. Jean-Pierre, R. & Blanchart, E. Seasonal and land-use induced variations of soil macrofauna composition in the Western Ghats, southern India. Soil Biology & Biochemistry. 37:1093, 2005.
14. Joshi, J.; Otway, S.J.; Koricheva, J.; Pfisterer, A.B.; Alphei, J.; Roy, B.A.; et al. Bottom-up effects and feedbacks in simple and diverse experimental grassland communities. Ecological Studies. 173:115, 2004.
15. Lavelle, P.; Decaëns, T.; Aubert, M.; Barot, S.; Blouin, M.; Bureau; et al. Soil invertebrates and ecosystem services. European Journal of Soil Biology. 42:3, 2006.
16. Morales, J. & Sarmiento, Lina. Dinámica de los macroinvertebrados edáficos y su relación con la vegetación en una sucesión secundaria en el páramo venezolano. Ecotropicos. 15 (1):99, 2002.
17. Nunes, D.H.; Pasini, A.; Benito, N.P. & Brown, G.G. Minhocas como indicadoras da qualidade ambiental. Um estudo de caso na região de Jaguapitã, PR, Brasil. Em: Minhocas na América Latina: biodiversidade ecologia. (Eds. G.G. Brown and C. Fragoso). Embrapa Soja. Londrina, Brasil. p. 467, 2007.
18. Paoletti, M.G. The role of earthworms for assessment of sustainability and as bioindicators. Agriculture, Ecosystems and Environment. 74:137, 1999.
19. Peck, S.B. A checklist of beetles of Cuba with data on distributions and bionomics (Insecta:Coleoptera). Arthropods of Florida and neighboring land areas. Volume 18. Florida Department of Agriculture and Consumer Services. Carleton University. Ottawa, Canadá. 241 p, 2005.
20. Rodríguez, I.; Crespo, G.; Rodríguez, C.; Castillo, E. & Fraga, S. Comportamiento de la macrofauna del suelo en pastizales con gramíneas naturales puras o intercaladas con leucaena para la ceba de toros. Rev. cubana. Cienc. Agríc. 36 (2):181, 2002.
21. Ruiz, Nuria; Lavelle, P. & Jiménez, J. Soil macrofauna field manual. Technical level. FAO. Rome. 101 p, 2008.
22. Ruiz-Cobo, D.H.; Feijoo, A. & Rodríguez, C. Macroinvertebrados edáficos en la cuenca del río Otún, Colombia. Acta Zoológica Mexicana (n.s.). Número Especial. 2:165, 2010.
23. Ruiz, Nuria., Mathieu, J., Célini, L., Rollard, Christine., Hommay, G., Lorio, E.; et al. IBQS: A synthetic index of soil quality based on soil macro-invertebrate communities. Soil Biology & Biochemistry. 43: 2032, 2011.
24. Sánchez, Saray. Acumulación y descomposición de la hojarasca en un pastizal de Megathyrsus maximus Jacq. y en un sistema silvopastoril de P. maximum y Leucaena leucocephala (Lam.) de Wit. Tesis presentada en opción al grado de Doctor en Ciencias Agrícolas. Instituto de Ciencia Animal. La Habana, Cuba. 135 p. 2007.
25. Sevilla, F.; Oberthür, T.; Barrios, E.; Escobar, G.; Madrid, O. Distribución y abundancia de la macrofauna asociada con unidades locales de clasificación de suelos en la microcuenca de Potrerillo, Cauca, Colombia. Tesis de Pregrado Ingeniería Agronómica. Universidad Nacional de Colombia, Centro Internacional de Agricultura Tropical (CIAT). Colombia. 129 p., s.f.
26. Velásquez, E.; Lavelle, P.; Rendeiro, C.; Martins, M.; Barot, S. & Grimaldi, M. Cambios en las comunidades de plantas influenciados por la macroagregación del suelo a través de las actividades de la macrofauna del suelo en la Amazonía Brasileira. [Disponible en:] http://www.iamazonica.org.br/conteudo/eventos/biodiversidadedesolo/pdf/Resumos/Painel3_Velasquez E.pdf, 2009.
27. Velásquez, Elena; Lavelle, P. & Andrade, Mercedes. GISQ, a multifunctional indicator of soil quality. Soil Biology & Biochemistry. 39:3066, 2007.
28. Xiaodong, Y. & Chen, J. Plant litter quality inï¬uences the contribution of soil fauna to litter decomposition in humid tropical forests, southwestern China. Soil Biology & Biochemistry. 41:910, 2009.
29. Zerbino, M. S. Evaluación de la macrofauna del suelo en rotaciones cultivos-pasturas con laboreo convencional. Acta Zoológica Mexicana (n.s.). Número Especial. 2:189, 2010.
30. Zerbino, María S. Evaluación de la densidad, biomasa y diversidad de la macrofauna del suelo en diferentes sistemas de producción. Tesis presentada para obtener el grado de Magister en Ciencias Ambientales. Universidad de La República. Montevideo, Uruguay. 92 p., 2007.
Recibido el 16 de enero de 2014
Aceptado el 28 de mayo de 2014

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}