










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkPastos y Forrajes
versión On-line ISSN 2078-8452
Pastos y Forrajes vol.40 no.4 Matanzas oct.-dic. 2017
ARTÍCULO DE INVESTIGACIÓN
Identificación y degradación de mimosina, un compuesto tóxico en Leucaena leucocephala (Lam.) de Wit
Identification and degradation of mimosine, a toxic compound in Leucaena leucocephala (Lam.) de Wit
Luis Alejandro Ospina-Daza, María Eugenia Buitrago-Guillen y Julio Ernesto Vargas-Sánchez
Universidad de Caldas, Calle 65 No. 26-10. apdo. aéreo 275, Manizales, Colombia
Correo electrónico: alejo.mvz7@gmail.com
RESUMEN
Leucaena leucocephala es conocida por ser una arbustiva que genera beneficios nutricionales para la producción ganadera en el mundo. No obstante, su consumo se ha limitado debido a factores antinutricionales como la mimosina, tóxico propio de la planta. Esta revisión recopila investigaciones de diferentes autores que identificaron y caracterizaron bacterias halladas en el rumen y en la rizosfera de las plantas, capaces de degradar la mimosina y sus productos de degradación, 3,4 y 2,3-dihidroxipiridona (3,4 y 2,3 DHP). La degradación de la mimosina (aminoácido no proteico que se encuentra en las hojas y las semillas de L. leucocephala) por las bacterias es un factor fundamental en la protección de los animales cuando consumen leucaena. La mimosina y sus productos de degradación han sido usados en la rizosfera por la cepa TAL1145 de Rhizobium, como fuente de carbono y nitrógeno; mientras que a nivel ruminal bacterias como Synergistes jonesii pueden degradarla. Recientes investigaciones han evidenciado que otras bacterias ruminales, como Streptococcus lutetiensis poseen la capacidad de degradar este compuesto. Tales avances llevarán a la especie L. leucocephala a otro nivel de conocimiento, en aras de buscar la mayor eficiencia y seguridad en su uso para la alimentación animal.
Palabras clave: alimentación de los animales, Streptococcus lutetiensis, Synergistes jonesii.
ABSTRACT
Leucaena leucocephala is known for being a shrub that generates nutritional benefits for animal husbandry in the world. Nevertheless, its consumption has been limited due to antinutritional factors such as mimosine, a toxic that exists in the plant. This review compiles studies by different authors who identified and characterized bacteria found in the rumen and in the rhizosphere of the plants, capable of degrading mimosine and its degradation products, 3,4- and 2,3-dihydroxypyridone (3,4 and 2,3 DHP). Mimosine (non-protein aminoacid which is found in L. leucocephala leaves and seeds) degradation by bacteria is a fundamental factor in the protection of animals when consuming leucaena. Mimosine and its degradation products have been used in the rhizosphere by the strain TAL1145 of Rhizobium, as carbon and nitrogen source; while at rumen level bacteria such as Synergistes jonesii can degrade it. Recent studies have proven that other rumen bacteria, like Streptococcus lutetiensis have the capacity to degrade this compound. Such advances will take the species L. leucocephala to another knowledge level, in order to search for higher efficiency and safety in its use for animal feeding.
Keywords: animal feeding, Streptococcus lutetiensis, Synergistes jonesii.
INTRODUCCIÓN
En el trópico, la nutrición de los rumiantes se basa principalmente en el consumo de pastos, subproductos de cosechas y, en menor medida, alimentos conservados, tales como ensilajes y harinas. No obstante, las gramíneas utilizadas en la alimentación poseen altos contenidos de fibra y bajos porcentajes proteicos, lo que trae como resultado una reducción considerable en el desarrollo general del animal, particularmente en la época de seca (Villanueva et al., 2013). Un gran número de árboles forrajeros, fundamentalmente leguminosos, se utilizan como suplemento en las dietas de baja calidad nutricional; estos se caracterizan por presentar un alto contenido de proteína y buena digestibilidad, comparados con los pastos (Pereyra et al., 2015).
Además de su favorable composición química, los árboles y arbustos forrajeros tienen otras propiedades positivas, ya que ayudan a recuperan suelos degradados (Rangel et al., 2016), incrementan su contenido de nutrientes debido a la simbiosis que establecen con bacterias del género Rhizobium (Higashide, 2014), protegen contra la erosión, y también pueden usarse como combustible y materiales de construcción.
Todas las especies de leguminosas contienen metabolitos secundarios, algunos de los cuales se denominan factores antinutricionales (FAN), que pueden causar un efecto negativo en el valor nutricional del alimento, así como en la salud animal. Los FAN pueden definirse como aquellas sustancias generadas por el metabolismo natural de las especies vegetales, y que, por diferentes mecanismos, ejercen resultados contrarios en la nutrición óptima de los animales, por causar disminución de los efectos digestivos y/o metabólicos (Rodríguez y Ledesma, 2014).
Las leguminosas generan estas sustancias a través de su metabolismo secundario, como un mecanismo de defensa ante el ataque de mohos, bacterias, insectos y pájaros; o, en algunos casos, como producto del metabolismo cuando están sometidas a condiciones de estrés. Estas plantas, al estar contenidas en los ingredientes de la dieta, reducen el consumo e impiden la digestión, la absorción y la utilización de los nutrientes por el animal (Casso y Montero, 1995).
Entre los FAN más estudiados, presentes en la mayoría de las especies de leguminosas, se pueden citar: lecitinas, taninos, glucósidos cianógenos, vicina y convicina, oligosacárido galactosil sacarosa, gomas galactomananas, saponinas, aminoácidos no proteicos, neurolaterígenos, análogos de arginina, alcaloides, ácido fítico, proteínas antigénicas y aminoácidos aromáticos; y entre estos últimos destaca la mimosina [3-N-(3-hidroxi-4-piridona) ácido-ao-aminopropiónico], presente en Leucaena leucocephala (Lam.) de Wit. (Casso y Montero, 1995).
El objetivo de esta revisión fue recopilar investigaciones realizadas por diferentes autores que identificaron y caracterizaron bacterias capaces de degradar la mimosina y sus productos de degradación, las cuales fueron halladas en el rumen y en la rizosfera de las plantas.
Características de la mimosina
La mimosina es un aminoácido no proteico, presente en la leguminosa forrajera tropical L. leucocephala, la cual ha sido responsable de síntomas de toxicidad en algunas especies de animales domésticos; entre estos síntomas destacan: alopecia, anorexia, pérdida de peso, salivación profunda, lesiones a través del esófago, papilas necróticas en el rumen y el retículo, hiperplasia de la glándula tiroides y bajos niveles de hormona tiroxina circulante (Xuan et al., 2013).
La mimosina posee un anillo aromático de 3-hidroxi-4-(1H)-piridona (3,4 DHP) (Nguyen y Tawata, 2016). Este compuesto desempeña un papel importante en la resistencia de la planta a una gran variedad de agentes fitopatógenos; además, por ser estructuralmente análogo a la tirosina, le permite un comportamiento inhibidor o antagonista en muchos procesos en los que esta es intermediaria. En los animales intoxicados por consumo de mimosina se manifiesta un efecto sobre la producción, que incluye bajos índices reproductivos debido a mortalidad embrionaria precoz y muerte perinatal (Hammond, 1995).
El anillo aromático de 3,4DHP se encuentra en estado libre en el rumen y en la circulación de los animales intoxicados, lo que indica que la mimosina es fácilmente hidrolizada en el rumen y excretada a nivel renal (Barros-Rodríguez et al., 2012). También se han encontrado pequeñas cantidades de mimosina en los nódulos nitrificantes de L. leucocephala y en el exudado adyacente a sus raíces, que es degradado a través de cepas del género Rhizobium; por otra parte, cuando los animales no adaptados se alimentan con leucaena por encima del 30 % de materia seca (MS) en su dieta, ello puede inducir cuadros de toxicidad y causar la muerte (Soedarjo y Borthakur, 1998).
Estudios recientemente publicados demuestran los esfuerzos para disminuir los contenidos de mimosina de las hojas de L. leucocephala utilizando metanosulfonato de etilo (EMS), y de esta forma mejorar el valor nutricional de la planta. Con este avance los silvicultores pretenden reducir significativamente el precio de la alimentación animal en el futuro (Zayed et al., 2014).
Identificación molecular de la mimosina
A pesar que L. leucocephala se considera como una promisoria fuente alternativa de proteína, y que además puede ayudar a mitigar la emisión de metano ruminal (CH4) en los trópicos (Soltan et al., 2013), la presencia de mimosina en un 2-10 % de MS en la hoja y en un 2-5 % de MS en la semilla limita la cantidad de follaje que se puede suministrar al ganado, ya que la mimosina (agente depilatorio) y su producto de degradación 3-hidroxi-4 (DHP)3 (potente bociógeno) se consideran tóxicos para muchas especies. La resistencia parcial a la toxicidad por mimosina en rumiantes de ciertas áreas geográficas ha sido atribuida a la capacidad de sus microorganismos ruminales, los que, restrictivamente, metabolizan mimosina y DHP. Un ejemplo es el ocurrido en Hawái, en el que cabras resistentes, con microorganismos eficientes que contrarrestaban los efectos de la toxicidad por mimosina, transfirieron los microorganismos al rumen del ganado de Australia, que era susceptible a dicha toxicidad (Lalitha et al., 1993).
Para la identificación de la mimosina y el DHP, el método de reacción más comúnmente usado es el colorimétrico con FeCl3 (Ilham et al., 2015). Este tiene poca sensibilidad y especificidad, ya que su pobre solubilidad en solventes acuosos y orgánicos limita su aplicación, pues requiere una alta concentración para tener medidas precisas. La cromatografía de gas-líquida, la cromatografía líquida y la cromatografía líquida de alta resolución en fase inversa de pares de iones requieren de instrumentación sofisticada y no brindan una mejoría en la sensibilidad (Lalitha et al., 1993).
El método espectrofotométrico para la estimación de mimosina y/o DHP es más sensible que otros reportados, y puede ser el más adecuadamente acoplado a una cromatografía de intercambio de iones y cromatografía de papel para el ensayo específico de estos compuestos. Este se basa esencialmente en la formación de un color amarillo intenso azoico entre la mimosina y/o DHP y la sal diazonio de p-nitroanilina (Ilham et al., 2015).
Aplicando la espectrofotometría se estimó que la cantidad de mimosina en las hojas de L. leucocephala varía entre 3,75 y 5,5 % de MS, en dependencia del tipo de hojas, la estación, la calidad de la tierra, etc. Debido a la alta sensibilidad del procedimiento, la mimosina fue detectable en los extractos decolorizados con carbón activado incluso después de altas diluciones, y las concentraciones de otros compuestos interferentes se volvieron despreciables para estas diluciones. Una aplicación útil de esta técnica se extendió a la evaluación de la toxicidad de la mimosina en el material de alimentación experimental basado en hojas de L. leucocephala. Las observaciones tempranas indicaron que, durante la degradación anaeróbica de estas hojas usando inóculo específico mezclado, la mimosina era activamente metabolizada, como lo mostró el método con FeCl3 después de la separación de la cromatografía de papel. Al aplicar el método antes descrito se confirmó que únicamente permanecían rastros de mimosina y de DHP, lo que indicaba casi el 99 % de degradación en 48 h, y la biomasa sólida hacía una elección adecuada de material de alimentación con alto contenido en proteína. En la comparación del método reportado con otras técnicas de estimación de mimosina se comprobó que el colorimétrico con FeCl3 requiere una solución de 20-500 µl para una medición precisa (Lalitha et al., 1993).
Degradación de la mimosina a partir de cepas de Rhizobium sp.
La mimosina es una toxina que se encuentra en grandes cantidades en las semillas y en las hojas de los árboles y arbustos de leguminosas del género Leucaena. En su estructura es análoga de la dihidroxifenilalanina (L-Dopa) con un anillo de 3-hidroxi-4-piridona en lugar de un anillo de 3,4 dihidroxifenil (Nguyen y Tawata, 2016). Esta toxina está distribuida en toda la planta, y se encuentra del 4 al 5 % en las semillas (base seca); en diferentes partes de la planta, como los tallos y las hojas, puede variar del 1 al 12 %, y en la raíz del 1 al 1,5 % (Soedarjo et al., 1994). Se ha demostrado que algunas cepas del genero Rhizobium pueden degradar la sustancia mimosina.
Según el trabajo realizado en Hawái por Soedarjo et al. (1994), un total de 32 cepas han sido recolectadas y se han cultivado en medios óptimos, como TY (Beringer, citado por MartUínez et. al., 2015); medio completo para crecimiento de rizobios, YEM (Vincent, citado por Abrahamovich et al., 2014); extracto de levadura-manitol (usado también para cultivo de rizobios) y un medio de Rhizobium-mimosina (RM), con el objetivo de saber cuáles de las cepas colectadas tenían la capacidad de crecer en un medio con la toxina. Se determinó que las cepas TAL1145 y TAL1566 poseían capacidad para utilizar la mimosina como única fuente de carbono y nitrógeno. Las tasas de crecimiento de dichas cepas en medio RM con diferentes concentraciones de mimosina se determinaron mediante la inoculación de 50 mL de este medio en botellas con tapón de rosca, de 250 mL, que contenían 0,5 mL de cultivo de Rhizobium. Los cultivos se sometieron a 28 °C, con agitación, y el crecimiento se determinó cada 6 h mediante la medición de la densidad celular como la densidad óptica en un colorímetro.
Se determinó que solo un número limitado de cepas de Rhizobium podía utilizar la mimosina como sustancia de crecimiento selectivo; en contraste, las cepas que no tenían esta capacidad formaron nódulos fijadores de nitrógeno, como es el caso de la MS13. La utilización de la mimosina puede ser un mecanismo especializado que algunos rizobios que viven en la rizosfera de la leucaena han desarrollado para sobrevivir, lo que proporciona una ventaja competitiva a ciertas cepas de Rhizobium. No se logró establecer una relación directa entre la capacidad de catabolizar mimosina y la capacidad de fijar nitrógeno, debido a que dos cepas, TAL1145 y TAL1566, degradan mimosina y la usan como fuente de nitrógeno y carbono, pero solo TAL1145 es buena formadora de nódulos nitrificantes, a diferencia de TAL1566 que tiene deficiencias para formarlos (Soedarjo et al., 1994).
En un estudio posterior a este, en la isla de Guam, se empleó la cepa de mejores resultados en cuanto a nodulación y aprovechamiento de la mimosina para sí misma, en términos de nitrógeno y carbono (TAL1145). Para ello se aislaron Rhizobium indígenas de los nódulos de L. leucocephala, con el fin de verificar si estos tenían la capacidad de degradar la mimosina y examinar si contenían el gen midA. Como se mencionó anteriormente la leucaena nodula con varios tipos de Rhizobium, tales como Rhizobium cepa TAL1145, Rhizobium cepa NGR234, y con cepas de Rhizobium tropici, como CIAT899. Entre estos rizobios, solamente la cepa TAL1145 y algunas cepas relacionadas pueden degradar la mimosina y utilizarla como fuente de carbono y nitrógeno, mientras que cepas como CIAT899 y NGR234 no pueden degradarla (Soedarjo et al., 1994).
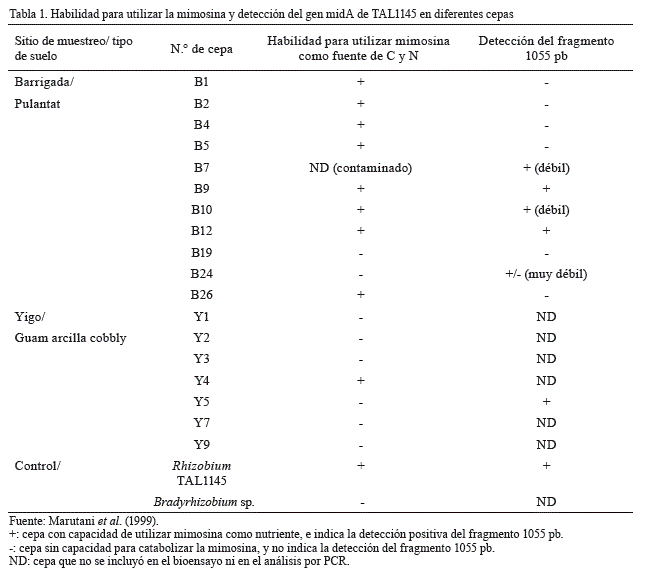
Después de aislar los rizobios, realizar el cultivo de las cepas halladas y someterlas a un medio líquido RM, y aislar y amplificar el ADN genómico de las cepas de Rhizobium que aprovecharon la mimosina se concluyó que ocho de las 11 cepas aisladas la utilizaban, ya que el medio RM cambió de color amarillo parduzco a incoloro. Esto fue corroborado por Soedarjo et al. (1994), quienes demostraron que la mimosina no se detectó en el medio por cromatografía líquida de alto rendimiento (HPLC) cuando el medio se volvió incoloro, con presencia de rizobios que la degradan. Como se muestra en la tabla 1, de los ocho degradantes de mimosina solo tres cepas generaron el fragmento de PCR 1055-pb, lo que sugiere que estas cepas contenían el gen midA de la cepa TAL1145 (Marutani et al., 1999).
Según lo descrito anteriormente, se evidenció la importancia de conocer más a fondo las cepas pertenecientes al género Rhizobium, que tienen la facultad de utilizar la mimosina y su producto de degradación 3,4 y 2,3 hidroxipiridona.
Soedarjo et al. (1994) aislaron en diferentes partes del mundo algunas cepas de Rhizobium de los nódulos de leucaena que cumplían con esta condición y que, además, podían utilizarla como fuente de carbono y nitrógeno. Aunque no se requiere la capacidad de catabolizar mimosina para la nodulación y la fijación de nitrógeno, esto proporciona una ventaja competitiva a los Rhizobium degradantes de la mimosina (mid+) en la rizosfera de la leucaena, al proveer una fuente selectiva de nutrientes y, al mismo tiempo, la inhibición del crecimiento de otros microorganismos y rizobios (Soedarjo y Borthakur, 1998). La cepa de Rhizobium TAL1145 es una de esas cepas mid+, conocida por ser competitiva para la ocupación del nódulo en la leucaena (Siddiqi y Athar, 2013), y se ha descrito como una formadora muy eficaz de nódulos fijadores de nitrógeno en Leucaena spp. La coincidencia en la capacidad de fijación eficiente de N2 y la capacidad competitiva de esta hacen que sea una opción ideal para su uso en preparaciones de inóculos.
Para identificar y caracterizar los genes mid presentes en el fragmento de 12 a 6 kb de TAL1145, los cuales son requeridos para la degradación de mimosina, se aisló el cósmido pUHR181 a partir de una biblioteca de clones de TAL1145 (Borthakur et al., 2003). Cuando este cósmido fue transferido a cepas no degradantes (mid-), como TAL182 y CIAT899, el producto de degradación 3-hidroxi-4-piridona (HP) se acumuló en el medio de cultivo. Esto sugirió que pUHR181 contenía genes para la degradación de la mimosina a HP. El plásmido pUHR191 es un derivado de pUHR181 que contiene un inserto de 12 a 6 kb, construido por la eliminación de un fragmento de aproximadamente 10 kb de pUHR181. Los transconjugantes de TAL182 y CIAT899 contienen pUHR191 que convierten mimosina en HP (Borthakur et al., 2003).
Inicialmente se establecieron 12 TAL1145 defectuosos en la degradación de la mimosina (mid-); estos se hicieron a través de Tn3Hogus (transposon), TnphoA (bacteriófago) o inserciones de casete de resistencia a la kanamicina. Un fragmento PstI (una enzima de restricción) de 5-0 kb de TAL1145, subclonado a partir de un clon cósmido que contiene genes mid para la degradación de mimosina, complementa la mayoría de los mutantes mid-. La secuenciación de este fragmento y el fragmento PstI de 0-9 kb que se encuentra adyacente identificó cinco genes: midA, midB, midC, midD y midR, de los cuales los tres primeros codifican proteínas transportadoras ABC implicadas en la absorción de mimosina; mientras midD codifica una aminotransferasa requerida para degradar la mimosina en HP, y midR es un gen regulador que codifica un activador transcripcional de tipo LysR. Por lo tanto, los genes mid son específicos para el Rhizobium de la leucaena y están ausentes en las cepas Rhizobium y en Bradyrhizobium spp. (Borthakur et al., 2003).
Tal como en la cepa TAL1145 se pudo determinar la presencia de genes responsables de la degradación y utilización de la mimosina, se debe estudiar más a profundidad cepas eficientes en la nodulación, que tengan potencial de degradar otros factores antinutricionales (Xu et al., 2013).
Degradación de la mimosina a partir de bacterias ruminales
La leguminosa arbórea L. leucocephala es un alimento de alta calidad que se emplea en los rumiantes, el cual es de vital importancia para la producción ganadera en los trópicos, a pesar de la presencia de mimosina en sus hojas. Este aminoácido tóxico no proteico limita la productividad y afecta adversamente la salud de los animales (Halliday et al., 2013). Dado que L. leucocephala tiene gran potencial en la producción, se han realizado estudios en aras de conocer más específicamente qué microorganismos ruminales podrían estar involucrados en la degradación de los productos derivados de la mimosina y así reducir la toxicidad en los rumiantes.
En Venezuela, Domínguez-Bello y Stewart (1991) aislaron una bacteria perteneciente al género Clostridium en ovejas alimentadas con leucaena que no desarrollaron síntomas tóxicos, la cual tenía la capacidad de degradar la 3-4 DHP. Para el aislamiento de la bacteria se utilizó contenido clarificado del rumen, que se sometió al crecimiento de bacterias en un medio de cultivo; este contenía micro- y macrominerales, resazurina, cisteína, fitona (peptona de soya) y azúcares, que en su conjunto promovían el crecimiento bacteriano. Adicionalmente, el medio se gaseó utilizando CO2 y quedó libre de oxígeno. Para la identificación de las bacterias anaerobias se usaron dos sistemas de pruebas comerciales (API-20A y 32A ATB), consistentes en tiras de cúpulas de ensayo que contenían sustratos deshidratados y propiciaban el crecimiento de bacterias del género Clostridium; a las bacterias que crecieron en los sistemas API se les adicionó mimosina, 3,4 DHP y 2,3 DHP. Como resultado, la cepa que degradaba los tóxicos mencionados correspondía a un Clostridium (denominado cepa 162), dada su baja proporción de guanina y citosina. La cepa 162 degradó mimosina, 3,4 DHP, 2,3 DHP y dihidroxifenilalanina (DOPA), pero no 3-hidroxipiridina o catecol cuando se añadieron estos compuestos al medio RPF (agar base Bird-Parker plasma de conejo y fibrinógeno) La degradación más alta fue hacia la mimosina (50-60 %) respecto a 2,3 DHP o 3,4 DHP (35-45 %).
Más tarde, Allison et al. (1992) identificaron y caracterizaron una bacteria aislada del rumen de una cabra (en Hawaii), capaz de degradar 3-4 DHP y su isómero 2-3 DHP, la cual fue nombrada Synergistes jonesii. Esta investigación se basó en la toxicidad que generaba el consumo de la leguminosa L. leucocephala en los rumiantes; sin embargo, en algunas partes del mundo se encontraron animales resistentes a la toxicidad de los componentes de esta planta, por su capacidad para degradar 3-4 DHP y su isómero 2-3 DHP.
S. jonesii se originó a partir de cuatro cepas (78-1, 100-6, 113-4, 147-1) aisladas de una cabra, en Hawaii (Jones y Megarrity, citados por Halliday et al., 2014); estas cepas se cultivaron en medios anaerobios similares a los descritos anteriormente para Clostridium, que fueron sometidos a kits de ensayo (API-ZIM, AN-IDENT, API-20A) para la identificación de bacterias Gram negativas anaerobias y de las enzimas que estas producen. Posteriormente, las bacterias identificadas se sometieron a una prueba de sensibilidad a antibióticos, basada en la inhibición del crecimiento de las cepas que ejerce la 3,4 DHP y visualizada por colorimetría. Este estudio permitió concluir que S. jonesii difería de cualquier organismo, después de haberla cotejado con aproximadamente 600 bacterias diversas, ya que la secuencia 16S de su ARN ribosomal es muy singular; además de usar como fuente principal de energía aminoácidos como la arginina y la histidina, factor que también lo separa de las otras bacterias con las cuales fue comparada.
Al realizar las comparaciones entre cepas de S. jonesii, los exámenes microscópicos indicaron que todas tenían morfología similar, y no se observaron diferencias entre las cepas 78-1,100-6 y 113-4; mientras que solo la 78-1 se sometió a la gama completa de pruebas fisiológicas. Se consideraron las pruebas más definitivas y únicas para estas bacterias con las cuatro cepas y no se detectaron diferencias fisiológicas entre estas últimas. Todas degradaron 2,3 y 3,4 DHP, y utilizaron arginina, histidina y DHP, además de otros sustratos energéticos para el crecimiento.
A partir de este trabajo, se describió que S. jonesii es una bacteria Gram-negativa, anaeróbica, no esporoforme, no motora; asimismo, tiene la capacidad de fermentar piridinedioles, su único hábitat conocido es el rumen y se aisló originalmente por su función de ayudar a las cabras de Hawaii que pastaban L. leucocephala (Holland-Moritz et al., 2014). El organismo no es omnipresente en las poblaciones de rumen, pero se han demostrado límites geográficos definidos para su distribución (Allison et al., 1992).
Cuando esta bacteria se inoculó en el rumen de animales no adaptados al consumo de leucaena, los efectos tóxicos se redujeron (Palmer et al., 2010). Sin embargo, a pesar de la inoculación, los animales de diversas latitudes (por ejemplo, Australia, Indonesia y Etiopía) solo pueden tolerar hasta un 30 % de leucaena en la dieta (Dalzell et al., 2012). En tales casos, se ha planteado la hipótesis de que las poblaciones de bacterias ruminales capaces de degradar la mimosina a DHP están saturadas por los altos niveles de ingestión de leucaena (mimosina).
Se conoce que la bacteria S. jonesii tiene la capacidad de degradar 3-4 y 2-3 DHP; sin embargo, no se ha confirmado su presencia o ausencia en los animales alimentados con leucaena en otros lugares en los que esta planta es nativa. Además, se desconoce si algunas bacterias tienen capacidad de degradar mimosina, ya que las mencionadas anteriormente solo tienen capacidad para degradar 3-4 y 2-3 DHP pero no mimosina (Jetana et al., 2012). Dalzell et al. (2012) sugirieron que puede haber otras bacterias con capacidad degradante para mimosina y sus metabolitos.
La degradación de mimosina se atribuye a un tipo de mimosinasa, enzima que se encuentra en la hoja de leucaena; a pesar de que el mecanismo de activación no se conoce, se cree que en el momento de ser masticada la hoja esta enzima se activa, al ser hidrolizada la mimosina y producir 3-4 DHP (Pereira et al., 2013). También se ha informado acerca de la actividad hidrolítica en el rumen de los animales alimentados con leucaena; por lo tanto, se presume que hay algunas bacterias que podrían degradar la mimosina (Kudo et al., 1984). Unido a lo anterior, se cree que la intoxicación por mimosina es escasa, ya que los animales son capaces de regular el consumo de L. leucocephala, con lo que evitan la posibilidad de intoxicación (Bacab et al., 2013).
Las aseveraciones de Kudo et al. (1984) y Dalzell et al. (2012) sobre la idea de encontrar más bacterias responsables de la degradación de los tóxicos más conocidos de L. leucocephala se concretaron recientemente. En este sentido, en el año 2015 un estudio reveló cuatro aislados bacterianos (Streptococcus lutetiensis, Clostridium butyricum, Lactobacillus vitulinus y Butyrivibrio fibrisolvens) que podían degradar completamente la mimosina dentro de los siete días de incubación. También se observó que C. butyricum y L. vitulinus fueron capaces de degradar parcialmente el 2,3-DHP dentro de los 12 días de incubación, mientras que S. lutetiensis fue capaz de degradar completamente tanto el 3,4 como el 2,3 DHP (Derakhshani et al., 2016).
CONCLUSIONES
La mimosina que se encuentra en la rizosferade L. leucocephala es beneficiosa al ser captada por ciertas cepas de Rizhobium, lo que favorece la degradación de la mimosina y la nodulación nitrificante. Por lo tanto, un aislamiento selectivo de estas cepas podría contribuir a la elaboración de inóculos específicos para cultivos intensivos de L. leucocephala.
La actividad de degradación de DHP por parte de las bacterias mencionadas en esta revisión confirma la importancia de los procesos catabólicos que tienen lugar en el rumen, para la tolerancia de los rumiantes a los metabolitos secundarios de las plantas.
Se necesita un rastreo exhaustivo del contenido ruminal del ganado en pastoreo de L. leucocephala, en aras de identificar otras bacterias transformadoras de mimosina y contribuir al desarrollo de inóculos más efectivos que puedan ser utilizados por los ganaderos contra la toxicidad de la mimosina.
REFERENCIAS BIBLIOGRÁFICAS
1. Abrahamovich, Eliana; López, Ana C. & Alippi, Adriana M. Diversidad de cepas de Agrobacterium rubi aisladas de arándanos. Rev. Argent. Microbiol. 46 (3):237-241, 2014.
2. Albert-Rodríguez, Anayansi & Rodríguez-Ledesma, Y. Estudio de los factores antinutricionales de las especies Morus alba Lin (morera), Trichanthera gigantea (H & B), nacedero y Erythrina poeppigiana (Walp. O. F), piñón para la alimentación animal. Tlatemoani. 17. http://www.eumed.net/rev/tlatemoani/17/pinon.html. [07/12/2016], 2014.
3. Allison, M. J.; Mayberry, W. R.; Mcsweeney, C. S. & Stahl, D. A. Synergistes jonesii, gen. nov.; sp.nov.: a rumen bacterium that degrades toxic pyridinediols. Syst. Appl. Microbiol. 15 (4):522-529, 1992.
4. Bacab, H. M.; Madera, N. B.; Solorio, F. J.; Vera, F. & Marrufo, D. F. Los sistemas silvopastoriles intensivos con Leucaena leucocephala: una opción para la ganadería tropical. AIA. 17 (3):67-81, 2013.
5. Barros-Rodríguez, M.; Solorio-Sánchez, J.; Ku-Vera, J.; Ayala-Burgos, A.; Sandoval-Castro, C. & Solís-Pérez, G. Productive performance and urinary excretion of mimosine metabolites by hair sheep grazing in a silvopastoral system with high densities of Leucaena leucocephala. Trop. Anim. Health Prod. 44 (8):1873-1878, 2012.
6. Belmar-Casso, R. F. & Nava-Montero, R. N. Factores antinutricionales en la alimentación de animales monogásticos. México: FMVZ-UADY; CRUPY-UACH. http://www.sian.info.ve/porcinos/publicaciones/ encuentros/viii_encuentro/roberto.htm. [07/12/2016], 2000.
7. Borthakur, D.; Soedarjo, M.; Fox, P. M. & Webb, D. T. The mid genes of Rhizobium sp. strain TAL1145 are required for degradation of mimosine into 3-hydroxy-4-pyridone and are inducible by mimosine. Microbiology. 149 (2):537-546, 2003.
8. Dalzell, S. A.; Burnett, D. J.; Dowsett, J. E.; Forbes, V. E. & Shelton, H. M. Prevalence of mimosine and DHP toxicity in cattle grazing Leucaena leucocephala pastures in Queensland, Australia. Anim. Prod. Sci. 52 (5):365-372, 2012.
9. Derakhshani, H.; Corley, S. W. & Al Jassim, R. Isolation and characterization of mimosine, 3, 4 DHP and 2, 3 DHP degrading bacteria from a commercial rumen inoculum. J. Basic Microbiol. 56 (5):580-585, 2016.
10. Dominguez-Bello, M. Gloria & Stewart, C. S. Characteristics of a rumen Clostridium capable of degrading mimosine, 3(OH)-4-(1H)-Pyridone and 2, 3 Dihydroxypyridine. Syst. Appl. Microbiol. 14 (1):67-71, 1991.
11. Halliday, M. J.; Giles, Hayley E.; Dalzell, S. A.; McSweeney, C. S. & Shelton, H. M. The efficacy of in vitro Synergistes jonesii inoculum in preventing DHP toxicity in steers fed leucaena-grass diets. Trop. grassl.-Forrajes trop. 2 (1):68-70, 2014.
12. Halliday, M. J.; Padmanabha, J.; McSweeney, C. S.; Kerven, G. & Shelton, H. M. Leucaena toxicity: a new perspective on the most widely used forage tree legume. Trop. grassl.-Forrajes trop. 1 (1):1-11, 2013.
13. Hammond, A. C. Leucaena toxicosis and its control in ruminants. J. Anim. Sci. 73 (5):1487-1492, 1995.
14. Higashide, Wendy. The isolation and characterization of a galactosly transferase mutant of Rhizobium sp. strain Tal1145 that nodulates Leucaena leucocephala. Honolulu, USA: University of Hawaii at Manoa. http:/hdl.handle.net/10125/32028. [07/12/2016], 2014.
15. Holland-Moritz, Hannah E.; Coil, D. A.; Badger, J. A.; Dmitrov, G. I.; Khouri, Hoda; Ward, Naomi L. et al. Draft genome sequence of the pyridinediol-fermenting bacterium Synergistes jonesii 78-1. Genome Announc. 2 (4):e00833-00814, 2014.
16. Ilham, Z.; Hamidon, H.; Rosji, N. A.; RamLi, N. & Osman, N. Extraction and quantification of toxic compound mimosine from Leucaena leucocephala leaves. Procedia Chem. 16:164-170, 2015.
17. Jetana, T.; Thongruay, S.; Uswang, S. & Hengtrakulsin, R. A comparative study on mimosine, 3,4-dihydroxy pyridone (3,4-DHP) and 2,3-dihydroxy pyridone (2,3-DHP), purine derivatives (PD) excretion in the urine, thyroid hormone and blood metabolites profiles of Thai swamp buffalo (Bubalus bubalis) and Murrah buffa (Bubalus bubalis). Trop. Anim. Health Prod. 44 (4):887-897, 2012.
18. Kudo, H.; Cheng, K.-J.; Majak, W.; Hall, J. W. & Costerton, J. W. Degradation of mimosine in rumen fluid from cattle and sheep in Canada. Can. J. Anim. Sci. 64:937-942, 1984.
19. Lalitha, K.; Vargheese, C. M. & Balasubramanian, N. Spectrophotometric determination of mimosine and 3-hydroxy-4-(1H)-pyridone- the toxic principles of Leucaena leucocephala. Analyt. Biochem. 213 (1):57-62, 1993.
20. Martínez, Rosario; Nebot, Elena; Porres, J. M.; Kapravelou, Garyfallia; Moral, Ana del; Talbi, Chouhra et al. Medicago sativa L: mejora y nuevos aspectos de su valor nutritivo y funcional por coinoculación bacteriana. Nutr. Hosp. 32 (6):2741-2748, 2015.
21. Marutani, M.; Richardson, J.; Edirveerasingam, V.; Taitano, D. & Borthakur, D. Indigenous Rhizobium strains from Guam contain a mimosine degrading gene. Micronesica. 31 (2):379-385, 1999.
22. Nguyen, B. C. Q. & Tawata, S. The chemistry and biological activities of mimosine: a review. Phytother. Res. 30 (8):1230-1242, 2016.
23. Palmer, B.; Jones, R. J.; Poathong, S. & Chobtang, J. The value of Leucaena leucocephala bark in leucaena-grass hay diets for Thai goats. Trop. Anim. Health Prod. 42 (8):1731-1735, 2010.
24. Pereira-Junior, G.; Pereira-Filho, M.; Roubach, R.; Barbosa, Paula de S. & Shimoda, E. Farinha de folha de leucena (Leucaena leucocephala Lam. de Wit) como fonte de proteína para juvenis de tambaqui (Colossoma macropomum CUVIER, 1818). Acta Amaz. 43 (2):227-234, 2013.
25. Pereyra, Gabriela; Hartmann, H.; Michalzik, Beate; Ziegler, W. & Trumbore, Susan. Influence of Rhizobia inoculation on biomass gain and tissue nitrogen content of Leucaena leucocephala seedlings under drought. Forests. 6 (10):3686-3703, 2015.
26. Rangel, W. M.; Thijs, S.; Janssen, J.; Oliveira-Longatti, S. M.; Bonaldi, D. S.; Ribeiro, P. R. et al. Native rhizobia from Zn mining soil promote the growth of Leucaena leucocephala on contaminated soil. Int. J. Phytoremediation. 19 (2):142-156, 2017.
27. Siddiqi, M. A. & Athar, M. Rhizobial competition and enhancing rhizobial colonization in the legume rhizosphere using a systemic fungicide. FUUAST J. Biol. 3 (2):45-52, 2013.
28. Soedarjo, M. & Borthakur, D. Mimosine, a toxin produced by the tree-legume Leucaena provides a nodulation competition advantage to mimosine-degrading Rhizobium strains. Soil Biol. Biochem. 30 (12):1605-1613, 1998.
29. Soedarjo, M.; Hemscheidt, T. K. & Borthakur, D. Mimosine, a toxin present in leguminous trees (Leucaena spp.), induces a mimosine-degrading enzyme activity in some Rhizobium strains. Appl. Environ. Microb. 60 (12):4268-4272, 1994.
30. Soltan, Y. A.; Morsy, A. S.; Sallam, S. M.; Lucas, R. C.; Louvandini, H.; Kreuzer, M. et al. Contribution of condensed tannins and mimosine to the methane mitigation caused by feeding Leucaena leucocephala. Arch. Anim. Nutr. 67 (3):169-184, 2013.
31. Villanueva, C.; Ibrahim, M.; Lombo, F. & Pérez, N. Potencial de las leñosas forrajeras en potreros para la alimentación del ganado en la época seca. En: D. Sánchez, C. Villanueva, G. M. Rusch, M. Ibrahim and F. DeClerck, eds. Estado del recurso arbóreo en fincas ganaderas y su contribución en la sostenibilidad de la producción en Rivas, Nicaragua. Turrialba, Costa Rica: CATIE. Boletín técnico No. 60. p. 42-48, 2013.
32. Xu, K. W.; Penttinen, P.; Chen, Y. X.; Chen, Q. & Zhang, X. Symbiotic efficiency and phylogeny of the rhizobia isolated from Leucaena leucocephala in arid-hot river valley area in Panxi, Sichuan, China. Appl. Microbiol. Biotechnol. 97 (2):783-793, 2013.
33. Xuan, T. D.; Tawata, S. & Khanh, T. D. Herbicidal activity of mimosine and its derivatives. In: A. J. Price and J. A. Kelton, eds. Herbicides-advances in research. Rijeka, Croatia: Intech. p. 299-312, 2013.
34. Zayed, M. Z.; Ahmad, F. B.; Zaki, M. A.; Hoand, W.-S. & Pang, S.-L. The reduction of mimosine content in Leucaena leucocephala (petai belalang) leaves using ethyl methanesulphonate (EMS). Arch. Appl. Sci. Res. 6 (4):124-128, 2014.
Recibido el 4 de abril del 2017
Aceptado el 29 de octubre del 2017


{kind=link}