










Meu SciELO
 Serviços customizados
Serviços customizadosServiços Personalizados
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkPastos y Forrajes
versão On-line ISSN 2078-8452
Pastos y Forrajes vol.40 no.4 Matanzas oct.-dez. 2017
ARTÍCULO DE INVESTIGACIÓN
Efecto de condiciones controladas en la germinación de cinco variedades de Morus alba L.*
Effect of controlled conditions on the germination of five Morus alba L. varieties*
Jorge Alberto Sánchez-Rendón1, Jorge Jesús Reino-Molina2, Mayté Pernús-Alvarez1, Dariel Morales-Querol2 y Giraldo Jesús Martín-Martín2
1Instituto de Ecología y Sistemática-Ministerio de Ciencia, Tecnología y Medio Ambiente Carretera de Varona #11835, Boyeros, CP 11900, La Habana, Cuba
2 Estación Experimental de Pastos y Forrajes Indio Hatuey, Matanzas, Cuba
Correo electrónico: jasanchez@ecologia.cu
RESUMEN
El objetivo de este estudio fue determinar el efecto de la temperatura del sustrato y de la alternancia de luz- oscuridad, sobre la respuesta germinativa de semillas frescas (obtenidas por vía sexual) de cinco variedades de Morus alba L. (cubana, tigreada, universidad, universidad mejorada y yu-62) cosechadas en Matanzas Cuba, procedentes de dos lotes diferentes. En los ensayos se empleó un diseño completamente al azar; los datos de porcentaje de germinación final y tiempo medio de germinación se procesaron mediante un análisis de varianza (ANOVA) de clasificación simple, con arreglo factorial de los tratamientos. La germinación se evaluó a temperatura fija (25 ºC) y en cuatro termoperíodos (25/30 ºC, 25/35 ºC, 25/40 ºC y 25/45 ºC), en dos condiciones (luz y oscuridad). Las semillas no presentaron dormancia seminal; los mayores porcentajes de germinación final para todas las variedades se alcanzaron a 25/35 ºC con iluminación, aunque también se obtuvieron altos porcentajes de germinación en oscuridad total cuando las semillas se colocaron a 25/40 ºC. Por otra parte, la temperatura fija de 25 ºC causó a todas las variedades una termoinhibición de la germinación. Se demostró la gran diversidad de respuestas germinativas en las variedades de M. alba estudiadas, en cada fecha de cosecha; lo que indica que el proceso de germinación puede ocurrir en diferentes condiciones ambientales, en particular donde exista una amplia fluctuación de la temperatura.
Palabras clave: luz, semillas, temperatura.
ABSTRACT
The objective of this study was to determine the effect of substrate temperature and light/darkness alternation, on the germination response of fresh seeds (sexually obtained) of five Morus alba L. varieties (cubana, tigreada, universidad, universidad mejorada and yu-62) harvested in Matanzas Cuba, from two different lots. In the essays a completely randomized design was used; the percentage data of final germination and mean germination time were processed through a simple-classification variance analysis (ANOVA), with factorial arrangement of the treatments. The germination was evaluated at fixed temperature (25 ºC) and in four thermoperiods (25/30 ºC, 25/35 ºC, 25/40 ºC and 25/45 ºC), under two conditions (light and darkness). The seeds did not show dormancy; the highest percentages of final germination for all the varieties were reached at 25/35 ºC with light, although high germination percentages were also obtained in total darkness when the seeds were placed at 25/40 ºC. On the other hand, the fixed temperature of 25 ºC caused in all the varieties a thermoinhibition of germination. The high diversity of germination responses in the studied M. alba varieties, in each harvest date, was shown; which indicates that the germination process can occur under different environmental conditions, particularly where there is a wide fluctuation of temperature.
Keywords: light, seeds, temperature.
INTRODUCCIÓN
Aún no se conoce lo suficiente acerca de las condiciones ambientales requeridas para la germinación de las plantas cultivadas, lo cual se debe, en parte, a que la acción del hombre ha favorecido la selección de genotipos de rápida germinación para diferentes ambientes (Smýkal et al., 2014; Dürr et al., 2015). Así, la domesticación de las plantas promueve rasgos en las semillas que difieren de las de sus progenitores o congéneres silvestres. Sin embargo, tener conocimiento acerca de los requerimientos germinativos de las plantas ayuda al desarrollo de los cultivos ya establecidos y a la creación de nuevas variedades/accesiones (Shalimu et al., 2015).
En Cuba, los estudios de germinación de las plantas de importancia económica para el hombre en los sistemas agrícolas se refieren en gran medida a cultivos como el arroz (Oryza sativa L.) y el tomate (Solanum lycopersicum L.), en los que se ha determinado, para algunas variedades, el efecto de la salinidad y de la temperatura del sustrato sobre el proceso germinativo y el desarrollo de las plántulas (Sánchez et al., 2001; Lamz y González, 2015); este aspecto también ha sido investigado, tanto en países en vías de desarrollo como en las naciones del llamado Primer Mundo (Górski et al., 2013; Tribouillois et al., 2016).
La germinación de las semillas es una etapa crítica del ciclo de vida de las plantas, que influye en el crecimiento y la producción de los cultivos, y puede ser afectada por diversos componentes abióticos y bióticos (Finch-Savage y Bassel, 2015). Entre estos se destacan, por solo citar algunos, las condiciones de cosecha y el método de obtención de las semillas; las diferencias entre poblaciones, accesiones y lotes (efectos maternos); y los efectos de la luz, la temperatura y la humedad del sustrato (Baskin y Baskin, 2014).
La variabilidad en la germinación es tan diversa que, en ocasiones, se clasifica erróneamente como la respuesta de una especie ante un elemento del ambiente; por ejemplo, cuando este proceso ocurre en presencia de la luz (Górski et al., 2013). Esto indica, por una parte, las diferentes estrategias regenerativas de una especie vegetal para enfrentar la incertidumbre ambiental; y, por otra, la selección artificial que han sufrido los cultivos durante varios siglos (Baskin y Baskin, 2014; Mitchell et al., 2016).
La morera (Morus alba L., Moraceae) es una planta cultivada multipropósito, originaria de la India y Asia Central, que ha sido introducida en casi todo el mundo, principalmente por su importancia en la industria de la seda y por su valor forrajero (Martín et al., 2014); y el método de propagación asexual (por estacas/esquejes) es el más empleado para esta especie (Olivera y Noda, 2010).
No obstante, la propagación por semilla (vía sexual) no solo constituye el método más económico y simple, sino también asegura la diversidad genética y con esto la supervivencia y producción de la especie bajo continuos cambios del ambiente (Fenner y Thompson, 2005; Finch-Savage y Bassel, 2015). Sin embargo, la información que existe sobre las clases de dormancia y los requerimientos germinativos de las semillas de M. alba es contradictoria. Por una parte, se plantea que las semillas frescas (cultivadas/silvestres) presentan dormancia fisiológica (Barbour et al., 2008), que es la misma presente en su congénere Morus nigra (Koyuncu, 2005) y en la mayoría de las especies de la familia Moraceae (Willis et al., 2014); también se ha informado la presencia de dormancia física en semillas frescas de M. alba (Barbour et al., 2008).
Górski et al.(2013) señalaron que la germinación se reduce por la baja relación rojo:rojo lejano y Permán et al.(2013) reconocieron que las semillas frescas no requieren pretratamiento para germinar (Permán et al., 2013).
En Cuba, la propagación por esquejes es el método más extendido para M. alba (Martín et al., 2014), y solo recientemente se introdujo el procedimiento de su reproducción por vía sexual. Por tanto, el objetivo de este estudio fue determinar el efecto de la temperatura del sustrato y de la alternancia de luz-oscuridad, sobre la respuesta germinativa de semillas frescas de cinco variedades de M. alba cosechadas en el país, procedentes de dos lotes diferentes.
MATERIALES Y MÉTODOS
Material vegetal. Las semillas frescas (recién cosechadas) de M. alba procedían del banco de recursos genéticos de la Estación Experimental de Pastos y Forrajes Indio Hatuey. Se emplearon las correspondientes a cinco variedades (cubana, tigreada, universidad, universidad mejorada y yu-62) que fueron introducidas desde Etiopía, Costa Rica y China (Martín et al., 2014), y cosechadas en marzo de 2014 (lote 1) y febrero de 2015 (lote 2) de una plantación destinada para la producción de semilla. La extracción y limpieza de las simientes se realizó manualmente, mediante despulpado y separación con un tamiz y enjuague con agua corriente; y seguidamente estas se secaron a temperatura ambiente y a la sombra durante dos o tres días. Su porcentaje de viabilidad se determinó mediante la prueba de tetrazolium (ISTA, 2007).
Requerimientos germinativos. Para lograr la respuesta germinativa en condiciones de laboratorio se utilizó un diseño completamente al azar. Se ensayaron diez tratamientos con la combinación de cinco termoperíodos [temperatura constante de 25 ºC y en regímenes alternos de 25/30 ºC, 25/35 ºC, 25/40 ºC y 25/45 ºC (8 horas para la temperatura más alta del termoperíodo y 12 horas a 25 ºC, con transición entre estas de 4 horas)] y la alternancia de luz-oscuridad [exposición a la luz durante 8 h a 40 mmol m-2 s-1 aproximadamente y a una longitud de 400-700 nm, coincidente con el período de mayor temperatura dentro de cada tratamiento; y a oscuridad total (constante), que se logró envolviendo las placas con dos capas de papel de aluminio]. La iluminación fue la recomendada en las normas establecidas internacionalmente (ISTA, 2007). Las condiciones a las que se sometieron las simientes simulaban las posibles variaciones de estos elementos del clima en el suelo de un área de campo, según el criterio de Sánchez et al. (2011). Las placas en las incubadoras se cambiaron de posición regularmente.
Las semillas, antes de la siembra, se esterilizaron con bicloruro de mercurio (1 g L-1) para prevenir la contaminación fúngica. Se utilizaron por tratamiento cinco réplicas de 25 semillas cada una. La siembra se realizó sobre agar hidrostático al 1 % en placas Petri de 9 cm de diámetro. En el caso de las semillas iluminadas, el conteo de germinación se efectuó diariamente durante 30 días. El conteo de germinación en las semillas expuestas a condiciones continuas de oscuridad total se determinó tres días después de haber concluido la germinación a la luz. El criterio para la germinación fue la emergencia de la radícula. En los tratamientos a la luz, el agar se mantuvo húmedo durante todo el experimento. Se determinó el porcentaje de germinación final a la luz y a la oscuridad, y para la condición de luz, el tiempo medio de germinación (días) según la metodología propuesta por Ranal et al. (2009).
El porcentaje de germinación final a la luz y a la oscuridad se empleó para calcular el índice de germinación relativa a la luz (GRL), que evidenció los requerimientos de luz para la germinación (Milberg et al., 2000). Este índice fue calculado según la fórmula: GRL = GL/(GD + GL), donde GL = porcentaje de germinación a la luz, y GD = porcentaje de germinación a la oscuridad. Para obtener el índice de GRL de cada variedad se utilizó el rango de temperatura óptimo (i.e., aquel donde se registró el mayor porcentaje de germinación en luz u oscuridad). Los valores de GRL variaron entre 0 (semillas que germinaron solo en la oscuridad) y 1 (semillas que germinaron solo a la luz). Si el índice GRL resultó superior a 0,75 se consideró que la especie/variedad fue dependiente de la luz (fotoblástica positiva), si fue menor a 0,25 se consideró repelente de la luz (fotoblástica negativa), y si el valor estuvo entre 0,25 y 0,75 se estableció como indiferente a la luz (Funes et al., 2009).
Al concluir las pruebas de germinación, se determinó el porcentaje de semillas vivas no germinadas presionando las semillas con una aguja para establecer si contenían un embrión blanco y firme (semillas vivas) o un embrión suave o gris, indicativo este último de no viabilidad seminal semillas muertas (Baskin y Baskin, 2014). Para las no germinadas en las temperaturas 25 ºC y 25/30 ºC, esta variable se estableció a través de la resiembra en la temperatura óptima de germinación (véase sección Resultados) y bajo luz blanca. La resiembra se realizó para determinar si las semillas adquirieron termoinhibición o termodormancia debido a condiciones inadecuadas de temperatura; se consideró que estas adquirieron termoinhibición si existían altos porcentajes de germinación, de acuerdo con el criterio de Geneve (2005).
Asignación de clases de dormancia. Se consideró que la especie poseía semillas dormantes si su germinación comenzaba después de 28 días en condiciones óptimas de germinación (Baskin y Baskin, 2014), o si al menos un 20 % de las semillas permanecieron vivas y sin germinar al final del experimento de germinación (Sánchez et al., 2015).
Análisis estadístico. El procesamiento estadístico se realizó de forma independiente para cada año de cosecha (lote de semilla). Los datos de porcentaje de germinación final y tiempo medio de germinación se procesaron mediante análisis de varianza (ANOVA) de clasificación simple, con arreglo factorial de los tratamientos; los del porcentaje de germinación se transformaron mediante el arcoseno de la raíz cuadrada de la proporción. No se aplicaron pruebas de comparaciones múltiples de medias a posteriori, debido a que algunas variables independientes representaron combinaciones de tratamientos cuantitativos (alternancia de luz-oscuridad y temperatura). Los análisis estadísticos se realizaron por el programa InfoStat v. 2015 (Di Rienzo et al., 2015).
RESULTADOS
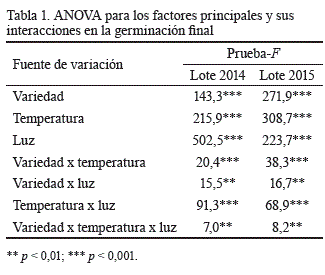
Requerimientos germinativos. El análisis de varianza demostró que los porcentajes de germinación final en ambos lotes de semillas frescas de M. alba fueron el resultado de la interacción altamente significativa que se estableció entre las variedades, las temperaturas del sustrato y la alternancia de luz-oscuridad (tabla 1). Además, las interacciones de primer orden (variedad x temperatura, variedad x luz, y temperatura x luz) y los efectos principales también desempeñaron un papel significativo en la germinación.
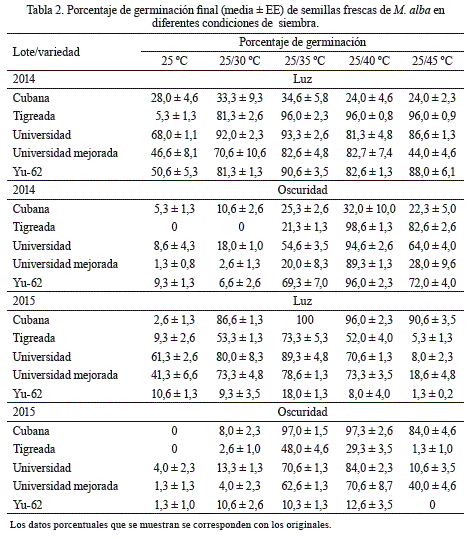
Los mayores porcentajes de germinación final en todas las variedades en cada lote se obtuvieron en el rango de temperatura alterna de 25/35 ºC con iluminación (tabla 2). Sin embargo, en el caso de la variedad tigreada de 2014, con iluminación, también las temperaturas de 25/40 ºC y 25/45 ºC fueron adecuadas para incrementar la germinación, aunque en este último rango no ocurrió la emergencia de las plántulas. Por su parte, en la variedad universidad de 2014 se obtuvieron incrementos significativos de la germinación en la siembra a 25/30 ºC bajo luz. En condiciones de oscuridad total igualmente se obtuvieron altos porcentajes de germinación final para todas las variedades cuando la siembra se realizó a 25/40 ºC para las semillas del lote de 2014, y a 25/35 ºC y 25/40 ºC para las semillas del lote de 2015.
En cambio, los menores porcentajes de germinación final se alcanzaron, para todas las variedades y lotes, en la temperatura constante de 25 ºC y la alterna de 25/30 ºC, efecto que fue mayor en las variedades cubana y tigreada, tanto en condiciones de luz como en oscuridad total (tabla 2). En estas condiciones de siembra también apareció un gran número de semillas vivas no germinadas que alcanzaron altos porcentajes de germinación cuando se coloraron a temperatura alterna de 25/35 ºC.
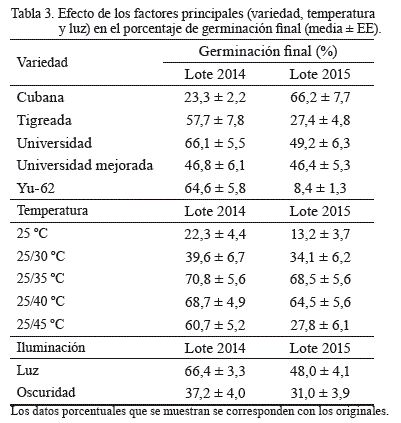
En general, las semillas frescas de las variedades cubana de 2014 y yu-62 de 2015 presentaron los menores porcentajes de germinación final (tabla 2), comportamiento que también se evidenció cuando se analizaron los efectos de los factores principales por separado (tabla 3); las variedades tigreada y universidad del lote 2014 mostraron una germinación superior que aquellas colectadas en 2015. Universidad mejorada mostró un comportamiento similar entre los años. Finalmente, el efecto del factor luz, como estimulante de la germinación, fue superior para ambos años, aunque su resultado fue más marcado en las semillas de 2014 (tabla 3).
El tiempo medio de germinación (o tiempo necesario para alcanzar el 50 % de las semillas germinadas) para ambos lotes de semillas fue afectado significativamente por la interacción del factor variedad y temperatura, y también por los efectos principales (tabla 4). El menor tiempo para germinar, independientemente de la variedad y el tiempo de colecta, siempre se alcanzó en la temperatura alterna de 25/35 ºC (tabla 5), con un rango desde 2,4 ± 0,1 días (en yu-62 de 2014) hasta 7,1 días en tigreada y yu-62 de 2015. También se apreció que a partir de 25/35 ºC la germinación fue más lenta, en particular en el rango de 25/45 ºC, donde tendió a concentrarse en unos pocos días. Lo anterior evidenció que el termoperíodo de 25/35 ºC fue el óptimo para la germinación de todas las variedades, pues con él se lograron los máximos porcentajes de germinación final en el menor tiempo posible. Cabe señalar que, para todas las variedades de 2014, el inicio de la germinación comenzó después del segundo día de siembra a 25/30 ºC y 25/35 ºC; y para las variedades de 2015, bajo las mismas temperaturas, la germinación se inició en el tercer día para cubana y universidad, en el cuarto día para tigreada y universidad mejorada, y en el quinto día para yu-62.
El índice de germinación relativo a la luz (GRL) en la temperatura de 25/35 ºC fue superior en todas las variedades de 2014, excepto en yu-62, respecto a los valores obtenidos para los lotes de semillas de 2015 (fig. 1). Las variedades tigreada y universidad mejorada presentaron valores de GRL superiores a 0,75; por tanto, fueron fotoblásticas. En el resto de las variedades, independientemente del tiempo de colecta, hubo valores de GRL desde 0,50 hasta 0,63, por lo que fueron consideradas como fotoblásticas indiferentes.
Asignación de clases de dormancia seminal
Las semillas no presentaron dormancia seminal; en la temperatura óptima el proceso germinativo comenzó antes de los 7 días, y alcanzó para la mayoría de las variedades más de un 70 % de germinación final, de acuerdo a la viabilidad inicial de los lotes (97 %). Además, en la temperatura considerada como óptima las semillas que no germinaron estuban muertas, según los resultados de la prueba de corte.
DISCUSIÓN
En las condiciones experimentales se evidenció que las semillas frescas de M. alba no presentaron dormancia física, ni fisiológica, pues se hidrataron y su germinación comenzó en menos de 28 días; resultado que coincide con lo informado por Permán et al. (2013) para esta especie. También se evidenció que la germinación de las variedades puede ocurrir en un amplio rango de temperaturas bajo condiciones de luz y oscuridad. Este aspecto se debe, posiblemente, al proceso de domesticación de la especie, que durante siglos ha favorecido que las semillas germinen con mayor rapidez y que exista una pérdida de la dormancia. Similar síndrome de domesticación se ha informado para un gran número de cultivos (Fuller y Allbay, 2009; Dürr et al., 2015) y representa una ventaja para el manejo y el rápido establecimiento de las plantas cosechadas (Smýkal et al., 2014). Sin embargo, la diversidad en la respuesta germinativa podría también ser una ventaja para enfrentar la incertidumbre o estrés ambiental que pueden hallar las semillas una vez que sean sembradas en condiciones de vivero
(Cochrane et al., 2015; Mitchell et al., 2016). De hecho, las condiciones de estrés, tanto en sistemas productivos como en sistemas naturales, son las que comúnmente encuentran las semillas cuando llegan al suelo (Finch-Savage y Bassel, 2015; Gardarin et al., 2016).
Los resultados de este estudio demostraron que la germinación de M. alba puede ser reducida por las condiciones inadecuadas de temperatura del sustrato y de iluminación. La germinación final disminuyó significativamente a temperatura fija de 25 ºC y alterna de 25/30 ºC, pero cuando se resembró a 25/35 ºC se lograron altos porcentajes de germinación, lo que evidenció el proceso de termoinhibición, tal como ha sido informado para numerosas especies (Geneve, 2005; Baskin y Baskin, 2014). Igualmente, cabe señalar que la germinación siempre se estimuló en condiciones de luz blanca, lo cual se debe a la fotosensibilidad reconocida para las semillas de diferentes genotipos de M. alba (Smýkal et al., 2014).
La temperatura del sustrato parece ser el factor principal que regula la germinación de las variedades estudiadas, pues la termoinhibición ocurrió tanto en las semillas sembradas bajo luz blanca como en aquellas en condiciones de oscuridad total. Sin embargo, también se demostró que en la temperatura óptima de siembra (25/35 ºC) las semillas frescas de tigreada y universidad mejorada se comportaron como fotoblásticas positivas, de acuerdo al índice de germinación relativa a la luz (Milberg et al., 2000). Lo anterior evidencia que dichas variedades fueron las más sensibles al estrés lumínico y a las bajas temperaturas del sustrato, resultado que podría tener gran implicación práctica en la reproducción de estas variedades en condiciones de vivero y de campo.
CONCLUSIONES
Existió gran diversidad de respuestas germinativas en las variedades de M. alba estudiadas, en cada fecha de cosecha; lo que indica que el proceso de germinación puede ocurrir en diferentes condiciones ambientales, en particular donde haya amplia fluctuación de la temperatura. Además, se pudo constatar que los rangos de temperatura muy altos, como el de 25/45 ºC, podrían ser efectivos para germinar, pero no para lograr emergencia de la plántula. Estos elementos deberán tenerse en consideración si se pretende reproducir la especie por vía sexual, pues dicha diversidad podría atentar contra la obtención de plantaciones uniformes, y también podría favorecer la adaptación de la especie a diversas condiciones climáticas.
Se recomienda determinar los efectos de los tratamientos pregerminativos de hidratación-deshidratación de estas simientes, para sincronizar e incrementar la germinación y el establecimiento de la especie en condiciones estresantes (calor, sequía, salinidad, entre otras).
REFERENCIAS BIBLIOGRÁFICAS
1. Barbour, J. R.; Read, R. A. & Barnes, R. L. Morus L. In: F. T. Bonner and R. P. Karrfalt, eds. The woody plant seed manual. Agriculture Handbook 727. Washington, DC: Department of Agriculture, Forest Service. p. 728-732, 2008.
2. Baskin, Carol C. & Baskin, J. M. Seeds: ecology, biogeography and evolution of dormancy and germination. 2 ed. San Diego, USA: Academic Press, 2014.
3. Cochrane, A.; Yates, C. J.; Hoyle, G. L. & Nicotra, A. B. Will among-population variation in seed traits improve the chance of species persistence under climate change? Glob. Ecol. Biogeogr. 24:12-24, 2015.
4. Di Rienzo, J. A.; Casanoves, F.; Balzarini, Mónica G.; Gonzalez, Laura; Tablada, Margot & Robledo, C. W. InfoStat versión 2015 Córdoba, Argentina: Grupo InfoStat, FCA, Universidad Nacional de Córdoba. http://www.infostat.com.ar. [13/12/2015], 2015.
5. Dürr, Carolyne; Dickie, J. B.; Yang, X. Y. & Pritchard, H. W. Ranges of critical temperature and water potential values for the germination of species worldwide: Contribution to a seed trait database. Agric. For. Meteorol. 200:222-232, 2015.
6. Fenner, M. & Thompson, K. The ecology of seeds. Cambridge, United Kingdom: Cambridge University Press, 2005.
7. Finch-Savage, W. E. & Bassel, G. W. Seed vigour and crop establishment: extending performance beyond adaptation. J. Exp. Bot. 67 (3):567-591, 2015.
8. Fuller, D. Q. & Allbay, R. Seed dispersal and crop domestication: shattering, germination and seasonality in evolution under cultivation. Ann. Plant Rev. 38:238-295, 2009.
9. Funes, G.; Díaz, Sandra & Venier, Paula. La temperatura como principal determinante de la germinación en especies del Chaco seco de Argentina. Ecol. Austral. 19 (2):129-138. http://www.scielo.org.ar/scielo.php?script=sci_arttext&pid=S1667-782X2009000200005&lng=es. [13/12/2015], 2009.
10. Gardarin, A.; Coste, F.; Wagner, Marine-Héléne & Dürr, Carolyne. How do seed and seedling traits influence germination and emergence parameters in crop species? A comparative analysis. Seed Sci. Res. 26 (4):317-331, 2016.
11. Geneve, R. L. Some common misconceptions about seed dormancy. Combined Proceedings International Plant Propagators' Society. USA: International Plant Propagators' Society, 2005.
12. Górski, T.; Górska, Krystyna & Stasiak, H. Inhibition of seed germination by far red radiation transmitted through leaf canopies. Pol. J. Agr. 13:10-38, 2013.
13. ISTA. International rules for seed testing. Bassersdorf, Switzerland: International Seed Testing Association, 2007.
14. Koyuncu, Fatma. Breaking seed dormancy in black mulberry (Morus nigra L.) by cold stratification and exogenous application of gibberellic acid. Acta Biol. Cracov. Series Botánica. 47 (2):23-26, 2005.
15. Lamz, A. & González, María C. Indicadores de crecimiento inicial y del estado nutricional para la selección temprana de genotipos de arroz (Oryza sativa L.) tolerantes a la salinidad. Cultivos Tropicales. 36 (2):41-48, 2015.
16. Martín, G. J.; Pentón, Gertrudis; Noda, Yolai; Contino, Y.; Díaz, Maykelis; Ojeda, F. et al. Comportamiento de la morera (Morus alba L.) y su impacto en la producción animal y la crianza de gusanos de seda en Cuba. Rev. cubana Cienc. agríc. 48 (1):73-78, 2014.
17. Milberg, P.; Andersson, L. & Thompson, K. Large-seeded species are less dependent on light for germination than small-seeded ones. Seed Sci. Res. 10 (1):99-104, 2000.
18. Mitchell, J.; Johnston, I. G. & Bassel, G. W. Variability in seeds: biological, ecological, and agricultural implications. J. Exp. Bot. 68 (4):809-817, 2017.
19. Olivera, Yuseika & Noda, Yolai. Origen, distribución, características botánicas, especies y variedades. En: Milagros Milera, comp. Morera. Un nuevo forraje para la alimentación del ganado. Matanzas, Cuba: EEPF Indio Hatuey. p. 9-19, 2010.
20. Permán, J.; Navarro, R.; Nicolás, J. L.; Prada, María & Serrada, R. Producción y manejo de semillas y plantas forestales. Madrid: Organismo Autónomo Parques Nacionales, Ministerio de Agricultura, Alimentación y Medio Ambiente, 2013.
21. Ranal, Marli A.; Santana, Denise G. de; Ferreira, Wanessa R. & Mendes-Rodrigues, C. Calculating germination measurements and organizing spreadsheets. Revista Brasil. Bot. 32 (4):849-855, 2009.
22. Sánchez, J. A.; Montejo, Laura; Gamboa, A.; Albert-Puentes, Delhy & Hernández, F. Germinación y dormancia de arbustos y trepadoras del bosque siempreverde de la Sierra del Rosario, Cuba. Pastos y Forrajes. 38 (1):11-28, 2015.
23. Sánchez, J. A.; Muñoz, Bárbara & Fresneda, J. Combined effects of hardening hydration-dehydration and heat shock treatments on the germination of tomato, pepper and cucumber. Seed Sci. Technol. 29 (3):691-697, 2001.
24. Sánchez, J. A.; Suárez, A. G.; Montejo, Laura & Muñoz, Bárbara C. El cambio climático y las semillas de las plantas nativas cubanas. Acta Bot. Cub. 214:38-50, 2011.
25. Shalimu, D.; Li, K.; Baskin, Carol C.; Baskin, J. M. & Liu, Y. Seed germination biology of four Pomegranate (Punica granatum) cultivars from Xinjiang, China. HortScience. 50 (6):826-829, 2015.
26. Smýkal, P.; Vernoud, Vanessa; Blair, M. W.; Soukup, A. & Thompson, R. D. The role of the testa during development and in establishment of dormancy of the legume seed. Front. Plant Sci. 35:1-19.https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4102250/pdf/fpls-05-00351.pdf, 2014.
27. Tribouillois, Hélène; Dürr, Carolyne; Demilly, D.; Wagner, Marine-Hélène & Justes, E. Determination of germination response to temperature and water potential for a wide range of cover crop species and related functional groups. PLoS One. 11 (8):e0161185, 2016.
28. Willis, C. G.; Baskin, Carol C.; Baskin, J. M.; Auld, J. R.; Venable, D. L.; Cavender-Bares, Jeannine et al. The evolution of seed dormancy: environmental cues, evolutionary hubs, and diversification of the seed plants. New Phytol. 203 (1):300-309, 2014.
Recibido el 24 de noviembre del 2016
Aceptado el 20 de septiembre del 2017
*Este resultado corresponde al proyecto: «Investigaciones con Morus sp. para el desarrollo de tecnologías sostenibles de alimentación y salud humana y animal en Cuba». Programa Nacional de Ciencia y Técnica «Producción de alimento animal», del Ministerio de Ciencia, Tecnología y Medio Ambiente, Cuba.
*This result corresponds to the project «Studies with Morus sp. for the development of sustainable human and animal feeding and health technologies in Cuba». National Program of Science and Technology «Feed production», of the Ministry of Science, Technology and Environment, Cuba.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}