










 Custom services
Custom services
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
Introducción
Leucaena leucocephala (Lam.) de Wit es una de las especies vegetales exóticas más importantes en los agroecosistemas ganaderos en la región de América Latina y el Caribe, pues es una planta que se debe considerar ante la erosión, la degradación y la compactación de los suelos, y la consecuente pérdida de la capacidad productiva de la tierra. De ahí que se considere un fitorecurso vital para la recuperación de la estabilidad, la fertilidad y el potencial agrícola de esta última (Calle et al., 2011). Es, además, un árbol multipropósito, que aporta abono verde al suelo, y del cual se obtiene madera (Brewbaker et al., 2016).
En la ganadería tropical sostenible, su uso en bovinos constituye un reto para producir de forma rentable en un ambiente de conservación de los recursos naturales, donde coexisten organismos diversos en ecosistemas saludables (Cortez-Egremy et al., 2016; Murgueitio-Restrepo et al., 2016). Se destaca también su inclusión en el silvopastoreo como alternativa a los sistemas con gramíneas exclusivamente, con el propósito de reducir las emisiones de los gases de efecto invernadero, en específico del CH4, que se origina en la fermentación ruminal de la fibra digestible (Molina et al., 2016; Morales-Velasco et al., 2016). Se ha demostrado que con los sistemas con árboles se logra el incremento productivo de los animales y de los servicios ambientales que se generan en el ecosistema (Chará et al., 2019).
Otro de los atributos más sobresalientes de la leucaena es que esta forrajera es capaz de producir hasta 1 t o más de semilla total (ST) ha-1. Específicamente, el cultivar Perú produce entre 450 y 610 kg de ST ha-1 sin poda, mientras que cuando se poda en junio, cada dos años, puede producir de 315 a 588 kg de ST ha-1 (Milera y Sánchez, 2017). Sin embargo, su producción de simientes se afecta por diferentes insectos fitófagos y patógenos, que impiden alcanzar estos rendimientos (Ahmed et al., 2016).
En ese grupo de insectos se destacan los que pertenecen a los ordenes Lepidoptera y Hemiptera, de las familias Cosmopterigidae y Pentatomidae, en cada caso, que afectan las inflorescencias y las legumbres, respectivamente. Los insectos de la familia Cosmopterigidae están incluidos en las más de 130 especies que existían en la región neotropical, según informes de una revisión de Landry (2001). Los de la familia Pentatomidae, al igual que los primeros, se consideran fitófagos plagas, de gran importancia económica en las leguminosas forrajeras de dicha región (Panizzi y Grazia, 2015; Grazia et al., 2015). El objetivo de este trabajo fue definir la potencialidad de dos insectos fitófagos para convertirse en plagas de la leucaena, durante su fenofase reproductiva, en Cuba.
Materiales y Métodos
Localización de las áreas experimentales. Las áreas experimentales se establecieron en la Estación Experimental de Pastos y Forrajes Indio Hatuey (EEPFIH), situada en la provincia de Matanzas, en la región oeste de Cuba.
Áreas experimentales evaluadas y sus característi-cas principales. Se evaluó un sistema silvopastoril (SSP) (cuartón) destinado a la ceba de toros, y establecido con la asociación de las variedades comerciales L. leucocephala cv. Perú (leucaena) y Megathyrsus maximus (Jacq.) B.K. Simon & S.W.L. Jacobs cv. Likoni (guinea likoni), aprobadas en Cuba por el MINAG (2017). Además, se estudió otro sistema, con similar composición de especies, pero destinado a la producción de carne bovina y semilla (SSPDP). Ambos se ubicaron en un suelo ferralítico rojo lixiviado (Hernández-Jiménez et al., 2015), con 1,3 ha de superficie y densidad de 396 árboles ha-1 (516 árboles en cada área) y seis años de explotación.
Principales elementos del manejo fitosanitario. No se aplicó fertilización inorgánica u orgánica, riego ni plaguicidas químicos o biológicos.
Método para el muestreo (recolección) de los insectos. Se utilizó la bolsa de nailon transparente.
Método para la evaluación de los insectos. Se aplicó el método del sobre en cinco puntos del cuartón.
Procedimiento para la caracterización de las lesiones del insecto fitófago asociado a las inflorescencias. En el SSP, en un año, en la etapa de floración plena (más del 70 % de la plantación florecida), se realizaron cinco muestreos durante tres semanas (dos en la primera y en la segunda, y uno en la tercera) en los meses de abril y agosto, que corresponden al período poco lluvioso (PPLL) y lluvioso (PLL), respectivamente. Se hicieron en los puntos de muestreo formados por uno o dos árboles, en dependencia de la cantidad de flores que poseían.
De los elementos del clima, se registraron las precipitaciones acumuladas en ambos meses: 19,1 mm en abril y 112 mm en agosto. Los datos procedieron de la estación meteorológica Indio Hatuey.
En cada muestreo, en los cuatro puntos cardinales de la planta, se tomaron cinco botones y cinco inflorescencias con pedúnculo. Las muestras se colocaron por separado en las bolsas de nailon transparente. Estas se cerraron con un cordón elastizado para evitar el escape de los insectos en el traslado del campo experimental al laboratorio de protección de plantas de la EEPFIH.
En el laboratorio, con un microscopio estereoscopio marca Zeiss, se observaron las 100 inflorescencias y los 100 botones de cada muestreo, con el propósito de contar el número de huevos, larvas, pupas y adultos del insecto. De la muestra total de inflorescencias, se tomaron, al azar, 30 con afectaciones visibles, para cuantificar, describir las lesiones, y determinar el número de huevos, larvas, pupas y adultos del insecto por flor individual, así como la cantidad de flores lesionadas.
Procedimiento para la caracterización de las lesiones del insecto fitófago asociado a las legumbres y semillas, y su relación con una enfermedad bacteriana. Durante tres años, en el SSP, se evaluó 5 % de las plantas de L. leucocephala cv. Perú, con el propósito de la recolección de legumbres con semillas verdes, en formación y maduras (en el momento de su aparición en cada caso), así como de los insectos presentes. La recolección se realizó cada quince días, para lo cual se utilizaron bolsas de nailon. Se cuantificaron los insectos y las lesiones que ocasionaron por legumbre. Además, se describieron dichas lesiones.
Se cuantificaron y se caracterizaron los síntomas bacterianos presentes en las legumbres recolectadas, asociados a las lesiones producidas por los insectos, para corroborar lo referido en estudios de Delgado et al. (1989) y Alonso et al. (1993). Además, se contabilizó el número de semillas infectadas por la bacteria. Posteriormente, se realizó el aislamiento e identificación de la especie bacteriana en el laboratorio de bacteriología vegetal del Centro Nacional de Sanidad Agropecuaria (CENSA) mediante pruebas morfológicas, fisiológicas y bioquímicas.
Para describir la relación entre el insecto y la presencia de los síntomas bacterianos en legumbres y semillas, se aplicó el análisis de regresión mediante el empleo del programa estadístico SPSS®, versión 15.0 para Windows®. Como norma de selección del modelo de mejor ajuste, se tuvieron en cuenta los criterios enunciados por Guerra et al. (2003).
Procedimiento para determinar la relación entre el número de insectos y las legumbres y semillas infectadas por la bacteria en cada época. En el SSPDP, durante dos años, se evaluaron cuatro picos de floración-fructificación de L. leucocephala cv. Perú (específicamente en momentos próximos a la cosecha). En el año l, el primer pico evaluado correspondió al período mayo-julio; el segundo, a octubre-noviembre. El tercer y cuarto pico, ambos correspondientes al año 2, se enmarcaron en abril-junio, y septiembre-noviembre, respectivamente. El primer y el tercer pico correspondieron al período lluvioso, y el segundo y cuarto, al poco lluvioso. La tabla 1 muestra los datos de los elementos del clima que se recopilaron.
Tabla 1 Elementos del clima durante los cuatro picos de floración-fructificación evaluados.

Fuente: Estación meteorológica Indio Hatuey
En los meses citados, cada siete días, se muestrearon al azar cinco plantas de leucaena (con el método que se informó). De estas, se cosecharon cinco racimos de legumbres con semillas en estado de formación o de cosecha (aproximadamente 25 % de las legumbres que puede producir un árbol), con síntomas o sin ellos. Se colocaron en bolsas transparentes de nailon, cerradas con un cordón elastizado, para trasladarlas conjuntamente con los insectos al laboratorio de protección de plantas de la EEPFIH, con el propósito de su observación en un microscopio estereoscopio, marca Zeiss.
En el laboratorio se cuantificó el número de insectos, legumbres totales e infectadas por la bacteria, así como de semillas también infectadas por dicho microorganismo. A los datos se les realizó una prueba de comparación de proporciones, con la utilización del Sistema de Comparación de Proporciones versión 2.1, creado en el CENSA (1998), para determinar en qué pico de floración-fructificación de cada época se favoreció más la presencia de los insectos y el desarrollo de la enfermedad bacteriana. Además, se usó el procedimiento antes descrito para el aislamiento e identificación de la bacteria, con vistas a corroborar la correspondencia con la especie encontrada. Para explicar la relación entre cantidad de insectos (ninfas y adultos) y legumbres y semillas infectadas por la bacteria, se realizó también un análisis de regresión, con el mismo programa estadístico y criterio de selección utilizado en el experimento anterior.
Estimación de las pérdidas de semillas por los insectos de las inflorescencias, y las legumbres. Se tomó como supuesto un punto intermedio (promedio) en los datos acerca de las características botánicas de la planta de L. leucocephala, ofrecidos por Hudges (1998). Esto es: una inflorescencia produce ocho legumbres, y una legumbre tiene 13 semillas, 1 kg posee 15 000 semillas, y su dosis de siembra es de 2 kg de semilla ha-1. Además, se consideró que una hectárea con 500 árboles produce 200 kg de semilla. También se tuvo en cuenta la cantidad de inflorescencias y legumbres muestreadas en el SSP en los experimentos antes descritos. El cálculo se realizó mediante una regla de tres, a partir de las inflorescencias y legumbres muestreadas, y según el supuesto citado.
Resultados y Discusión
Al evaluar las inflorescencias, se confirmó la presencia del microlepidóptero Ithome lassula Hodges (Lepidoptera: Cosmopterigidae) desde la fase de abotonamiento y en la inflorescencia propiamente dicha. Este insecto produjo lesiones que se manifestaron como perforaciones circulares en la base de las flores individuales. Generalmente, se observó una en cada flósculo, aunque en alguna ocasión se registraron dos. Estas observaciones resultan similares a lo informado por Alonso et al. (2015).
Cada larva se puede alimentar de una, y hasta de siete flores individuales a la vez, pues al mismo tiempo perfora la base de varias florecillas y consume las diferentes estructuras reproductivas. Además, puede llegar a devorar el capítulo. Si se alimenta de los botones, estos no abren, y se impide así la formación de la inflorescencia. De ahí que todas estas lesiones en su conjunto provoquen que no se formen las legumbres y, por ende, las semillas correspondientes. Las flores individuales se tornan de una coloración parda oscura con relación a las sanas, y las larvas crean una cápsula formada por hilos de seda y pequeñas partes de las florecillas que llevan consigo hasta el momento de empupar. Por lo general, se encontraron dos larvas como promedio por inflorescencia (tabla 2) y, en ocasiones, hasta 12, lo que supera la cifra de 7-9, referida por Elder (2008) en estudios relacionados con esta especie insectil, quien también trabajó con la leguminosa evaluada en esta investigación.
Tabla 2 Número total y promedio de lesiones y larvas de I. lassula en botones e inflorescencias de L. leucocephala.
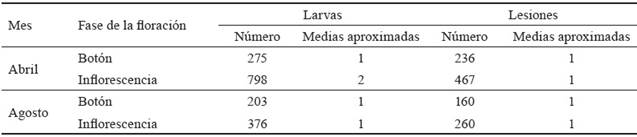
Los datos de la tabla 2 muestran incremento en el número de larvas y lesiones, de la fase de botón a la de inflorescencia, en ambos picos de floración. Dicho incremento fue superior durante las evaluaciones de abril con respecto a las de agosto, lo que se relaciona con el tamaño pequeño de los huevos (0,5 mm). Estos se insertan en las yemas durante los primeros momentos del desarrollo del capítulo y de las larvas neonatas (menos de 5 mm), lo que dificulta su detección de manera fácil (no se observaron en ninguno de los casos). De acuerdo con lo que plantea Elder (2008), no es hasta que abre el botón que se puede observar mayor número de larvas (con el aumento de su tamaño hasta 5 mm de longitud) y de sus lesiones.
La precipitación acumulada durante agosto resultó superior con respecto a abril. Esto pudo influir en la disminución de las larvas por el efecto directo de la lluvia, o debido a una mayor posibilidad de producir pudriciones de las inflorescencias por la alta humedad, la contaminación por las excretas de los insectos y el crecimiento de hongos saprófitos. De ahí que esta mayor afectación por I. lassula, en las inflorescencias durante el pico de floración en abril, período poco lluvioso, es de vital importancia, pues en dicha etapa es cuando la producción de semillas del cultivar Perú es mayor (Matías, 1999). Además, es importante señalar que la presencia de este cosmopterígido afecta las inflorescencias y, por ende, la producción de semillas en los cultivares Cunningham, Ipil Ipil y CNIA 250 de L. leucocephala (Alonso, 2009), así como en las especies Leucaena diversifolia (Schltdl.) Benth., y Leucaena spp. (Azani et al., 2017).
Se determinó que la lesión que causa el insecto Loxa viridis (Palisot de Beauvois) (Hemiptera: Pentatomidae) consiste en pequeñas perforaciones de forma circular, que miden 2 mm de diámetro, producidas con el estilete del insecto en la superficie de la legumbre, fundamentalmente en la zona donde la semilla está en formación (Alonso, 2009). Al inicio, aparece un ligero abultamiento de la epidermis de la legumbre, debido a la succión de la savia por el pentatómido, que en su parte terminal deja una gota de secreción salival. Posteriormente, se observa que el tejido donde se produjo la punción se necrosa, y da la impresión de un punto de color negro. Se aprecia que en cada legumbre se encuentra un número variable de picaduras.
Este pentatómido también se halla en las otras tres variedades comerciales de L. leucocephala (Cunningham, Ipil Ipil y CNIA 250) existentes en Cuba (Alonso, 2009).
En cuanto a los síntomas de la enfermedad bacteriana, se observa su aparición en las legumbres jóvenes, con úlceras pequeñas amarillo-parduscas, inicialmente, de las que emanan exudaciones con aspecto pegajoso. A medida que avanza la enfermedad, las úlceras se tornan pardo oscuras y aumentan en número y tamaño, hasta llegar a producir afectaciones similares en las semillas en formación, que impiden finalmente su maduración. Y si lo hacen, se pudren por la presencia de las exudaciones bacterianas, lo que se corresponde con lo descrito por Delgado et al. (1989). En Cuba se informaron estos síntomas por Alonso (2009), quien identificó la especie bacteriana Pectobacterium carotovorum subsp. odoriferum (Enterobacteriales, Enterobacteriaceae).
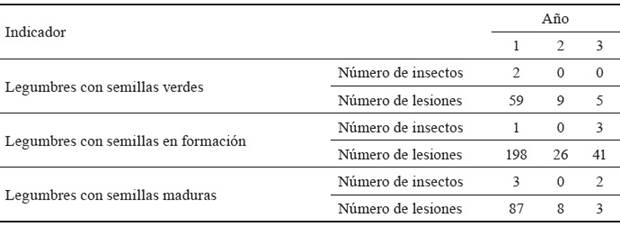
Como muestra la tabla 3, en los tres años de evaluación, a pesar de la baja detección de individuos de L. viridis, al cuantificar las lesiones se observó que el número de picaduras tendió al incremento en las legumbres con simientes en estado de formación con respecto a aquellas con semillas verdes. Y esto es porque poseen mayor cantidad de savia disponible, que hace que aumente la avidez de los insectos por ellas, además de que se acumulan las picaduras del estado inicial. Mientras, en las legumbres en estado de cosecha (con semillas maduras), las lesiones disminuyeron a causa del endurecimiento alcanzado por la maduración, que les confiere una consistencia coriácea, y también a consecuencia del cambio de coloración, de café a marrón, que enmascara la lesión, criterio que coincide con lo expuesto por Hudges (1998).
Tabla 3 Cantidad de insectos y lesiones por L. viridis en las legumbres de L. leucocephala cv. Perú.
El resultado del análisis de regresión para determinar la relación entre número de picaduras realizadas por los insectos y semillas infectadas por la bacteria, así como la relación entre las legumbres infectadas y el número de semillas que se afectaron por dicho microorganismo, indicó mejor ajuste con las ecuaciones cúbicas. En la figura 1, la curva muestra que inicialmente hubo valores ínfimos de picaduras en las legumbres con semillas verdes, lo que se debe a que estas no son tan preferidas por el insecto. Sin embargo, cuando alcanzaron el estado en formación, aumentó de forma abrupta el número de semillas infectadas por la bacteria. Esto no es solo por la acumulación de picaduras hasta la etapa de maduración, sino como expresión del desarrollo de la enfermedad, lo que evidencia el carácter transmisor del pentatómido referido por Alonso et al., (1993). Un comportamiento similar ocurrió entre las legumbres y las semillas infectadas, pues a medida que maduró la legumbre infectada, se produjo mayor número de semillas afectadas por la bacteria (figura 2).
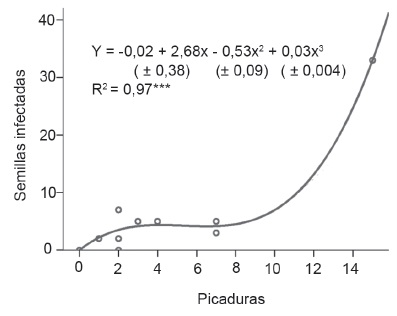
Figura 1 Relación entre picaduras por L. viridis y semillas infectadas por P. carotovorum subsp. odoriferum.

Entre el número de picaduras y las legumbres infectadas no se encontró una relación estrecha, lo que se pudo deber al hecho de que, aunque el insecto realiza varias picaduras en la misma legumbre, algunas de estas lesiones no se localizan sobre la zona de las semillas en formación.
De acuerdo con el resultado que se muestra en la tabla 4, se comprobó que en el PPLL se produjo incremento significativo en el porcentaje de legumbres infectadas por P. carotovorum subsp. odoriferum, con respecto al PLL, debido a que el número de individuos de L. viridis fue también superior. Esto confirma que el insecto se favoreció por un menor acumulado de lluvias, que además aumenta la posibilidad de transmisión de la bacteria, a pesar del menor número de legumbres en las plantas muestreadas en ese pico de floración-fructificación. Este efecto se observó de igual manera en el número de semillas afectadas por la bacteria en el PPLL (4 188), que resultó, aproximadamente, el doble de las semillas infectadas que en el PLL (1 962).
Tabla 4 Número de pentatómidos y su relación con las legumbres infectadas durante los picos de floración-fructificación por período.

*p ≤ 0,05
1Legumbres infectadas por P. carotovorum subsp. Odoriferum
A pesar del bajo número de individuos de L. viridis recolectados, su presencia en el campo se manifestó por las lesiones que permanecieron en las legumbres, lo que además propició que se produjeran putrefacciones por la bacteria, que al afectar a las semillas provocan una disminución de su valor agrícola.
En la figura 3 y 4 se observa la relación entre número de insectos, las legumbres y las semillas infectadas por la bacteria, como resultado del análisis de regresión, las ecuaciones cúbicas mostró el mejor ajuste (R2 = 0,90 y 0,91), se observó que inicialmente se produjo declinación en la curva, que pudo estar dada por la presencia de una generación que culmina su ciclo de vida, así como por un número considerable de puestas de huevos en este período. Esto implica que al eclosionar aparezca un grupo numeroso de ninfas en las legumbres con semillas verdes, en las que las lesiones por los insectos y los síntomas que ocasiona la bacteria son menos evidentes. Mientras, cuando estas ninfas alcanzan el estado adulto provocan mayor número de lesiones en la misma medida que las legumbres poseen semillas en formación, lo que explica el incremento de la afectación por la bacteria en dichos órganos. Finalmente, ocurre otro descenso en la curva, lo que indica que la afectación por la bacteria disminuye. Esto se debe a que, con el proceso de maduración, las lesiones por los insectos son menos posibles por el aspecto coriáceo de las legumbres antes mencionado y el endurecimiento de las semillas.

Figura 3 Relación entre el número de L. viridis y legumbres infectadas por P. carotovorum subsp. odoriferum
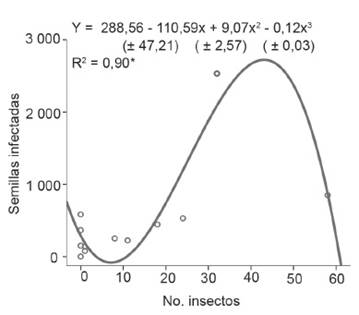
Figura 4 Relación entre el número de L. viridis y semillas infectadas por P. carotovorum subsp. odoriferum.
A pesar de que en la literatura actualizada disponible no se encontró ningún criterio acerca de los resultados descritos anteriormente, Lenné (1991) desde que detectó la presencia de insectos pentatómidos, asociados a bacterias en legumbres y semillas de leucaena, afirmó que existía la posibilidad de ese comportamiento, dado el vínculo directo entre ambos organismos. En Brasil, Panizzi y Rossi (1991) también indicaron la afectación de la especie insectil Loxa deducta (Walker) (Hemiptera: Pentatomidae) en leucaena y soya.
Segarra-Carmona et al. (2016) señalaron que, en México, L. viridis tiene como hospedante la leguminosa Senna papillosa (Britton & Rose). Esta planta se puede utilizar como componente del alimento animal, como cortina rompevientos, y en sistemas agroforestales para proporcionar sombra al cafeto, según indican Sánchez-Bernes y Moya-Caderón (2018).
Al estimar los daños que puede causar I. lassula en las 1 600 inflorescencias muestreadas (botones y abiertas) en ambos períodos, se produciría una pérdida de 8 kg de semillas, al dejarse de formar 8 984 legumbres y 116 972 semillas, lo que representaría 4 ha sin sembrar. En términos de producción, en una hectárea con 500 árboles podría dañarse 71 % de la producción semillera, pues se afectarían 20 500 inflorescencias, por lo que se dejarían de formar 164 000 legumbres, y se perderían 142 kg de semillas de los 200 kg posibles a obtener, según se tiene como referencia para esta unidad de superficie, sin la utilización de insumos externos. Esta cifra podría ser superior, si se considera que Partridge (1988) señaló que el cultivar Perú puede producir, aproximadamente, 100 kg de semilla ha-1 cuando no se emplea riego.
A partir de las afectaciones ocasionadas por el microlepidóptero I. lassula, es válido considerarlo como plaga del cultivo de L. leucocephala en Cuba, si se tiene en cuenta que el porcentaje de afectación (71 %) justificaría la toma de medidas de manejo fitosanitario, pues según Elder (2008) el control se realiza cuando en las inflorescencias el daño causado por las larvas de esta especie, es superior al 10 %. Por tanto, los resultados reafirman el criterio de este autor con relación a los daños que puede causar este insecto en la leguminosa en estudio, al señalar que se han registrado reducciones de hasta 10 veces la cantidad de legumbres que se pueden formar. Esto, a su vez, reduce de manera similar la producción de simientes.
La importancia de la especie I. lassula radica, además, en el hecho de que especies de este género (Ithome) se han informado en otros países, en los que atacan otras leguminosas de importancia similar para los sistemas silvopastoriles, como Acacia y Prosopis (Gallaher y Merlin, 2010). Esto implica que, de estar presente en otras leguminosas arbóreas, el daño que ocasionan puede ser notable, con impacto negativo para la ganadería del país. Como señaló Elder (2008), en Australia, I. lassula se considera abundante y frecuente, y de interés para los productores de semillas de leucaena, ya que un solo botón dañado impide la formación de la inflorescencia. Se registran entonces reducciones importantes en la cantidad de legumbres y, por consiguiente, en la producción semillera. De igual manera, Ananthakrishnan y Sivaramakrishnan (2017) señalan a este insecto como una plaga importante en dicha leguminosa en los Ghats occidentales, conocidos como las montañas Sahyadry, en la India.
En lo que respecta a las pérdidas por la afectación bacteriana de la especie P. carotovorum subsp. odoriferum, asociada al pentatómido L. viridis, se estimaron a partir de 1 175 legumbres muestreadas en total. De estas, 301 resultaron infectadas por la bacteria. Por tanto, en una hectárea con 500 árboles se podrían afectar 59 000 legumbres y 767 000 semillas, lo que representaría un déficit de 51 kg, y una merma en la producción de semilla de 26 %, porcentaje que se encuentra en el rango de 17 - 49, obtenido por Delgado et al. (1989). Estos autores evaluaron las afectaciones que provocó la bacteria en las semillas de diferentes variedades de L. leucocephala, en las que estaba incluido el cultivar Perú.
El daño que ocasiona la bacteria, debido en gran medida a su penetración a través de las lesiones del pentatómido, y a su posible transmisión por parte de este, resulta lo suficientemente significativo para considerar el comportamiento de este insecto como plaga de interés para el cultivo. Esto tiene relación directa con la importancia de los insectos como transmisores de enfermedades bacterianas, pues se cita que 15 de las más de 200 enfermedades que diseminaron los insectos y arácnidos en el 2015, eran de origen bacteriano (Ritacco, 2015).
Conclusiones
Asociados a los órganos reproductivos de la leucaena, se detectaron con potencialidad para convertirse en plagas, el microlepidóptero I. lassula, por su afectación directa en las inflorescencias, y el pentatómido L. viridis en las legumbres. Este último, indirectamente, afecta también la producción de semillas, al comportarse como un transmisor de la bacteria P. carotovorum subsp. odoriferum; además de las pérdidas que puede causar, según la estimación que se realizó.
Esto sugiere la necesidad de continuar con la ejecución de nuevas investigaciones que permitan obtener más información acerca del ciclo biológico de los insectos estudiados y su sincronía con la fenofase reproductiva de la forrajera, con vistas a diseñar de manera eficiente una estrategia agroecológica que sea efectiva para su manejo, y evitar así pérdidas en sus rendimientos.

