










 Servicios personalizados
Servicios personalizados
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
Chascolytrum subaristatum (Lam.) Desv. (= Briza subaristata Lam.) es una especie de la familia Poaceae, subfamilia Pooideae, tribu Poeae, subtribu Calotthecinae, del género Chascolytrum (Anton y Zuloaga, 2020), del que se citan tres especies introducidas en Argentina (Zuloaga et al., 1999).
En la provincia de Entre Ríos, Argentina, se puede encontrar en áreas naturales. Veintitrés botánicos junto a sus colaboradores colectaron esta especie, al menos 53 veces desde 1903, en once departamentos de la provincia. En la Carta de Suelos de Entre Ríos se encuentra citada en 13 departamentos. Según estas colectas y registros, se halló en los 17 departamentos de esta provincia, en áreas de vegetación xerófila a hidrófila, pastizales y praderas.
Las excursiones botánicas realizadas a estos departamentos (Galussi et al., 2015; 2016) permitieron localizar Ch. subaristatum en áreas naturales reducidas, generalmente bajas y húmedas. Se halló en dos sitios en Paraná y en cuatro sitios en Villaguay, junto a 37 especies, en su mayoría gramíneas herbáceas. La frecuencia relativa en que se encontró estuvo entre 14 y 35 % y la cobertura, entre 1,8 y 5,8 %.
Se trata de una especie perenne, cespitosa, que tiene de 10 a 80 cm de altura. Presenta hojas lineales largas de 2 mm, algo ásperas, lígula membranosa, partida y glabra. Según los estudios de anatomía foliar realizados por Moya et al. (2019), Ch. subaristatum tiene un patrón anatómico de gramíneas C3, con células de parénquima incoloro en el mesofilo. Las paredes periclinales externas de las células epidérmicas de las caras adaxial y abaxial se encuentran impregnadas de lignina. Los tallos muestran sección circular. Posee una epidermis uniestratificada, sin pelos y con cutícula gruesa, y un anillo esclerenquimático subepidérmico. Los haces vasculares colaterales cerrados se disponen en 3 o 4 anillos concéntricos, con médula parenquimática o hueca.
Ch. subaristatum es buena forrajera de campos altos en flor, desde fines de julio hasta marzo (Burkart, 1969). La digestibilidad del limbo de las hojas es de valor nutritivo medio, con relativamente bajo contenido de materia seca, buen contenido proteico y energía metabólica. Por su crecimiento en otoño-invierno-primavera, es de considerable importancia en áreas naturales porque aporta alimento en el sistema productivo silvopastoril.
Como inflorescencia presenta una panoja contraída, erecta, algo mutante en la madurez, con ramas laterales breves, espiguillas medianas y multifloras, de 4,5 a 8 mm de longitud (Burkart, 1969). En la época de floración, en octubre, sus flores son casmógamas, y cleistógamas en noviembre y diciembre. Las plantas son muy variables, en el tamaño de su panoja como en el de sus espiguillas y antecios (Rosengurtt et al., 1968). Según la descripción de Pohl (1994) y Calderón-de-Rzedowski y Rzedowski (2001), la inflorescencia es una panícula de 5 a 15 cm de largo, contraída, con pedicelos de 5 a 15 mm de largo. Sus espiguillas son más o menos ovoides, algo comprimida, de 3,5 a 8,0 mm por 2,5 a 6,0 mm de ancho, péndulas en la madurez.
Las observaciones de Galussi et al. (2017) en un período de crecimiento permitieron definir, entre otras características de Ch. subaristatum, la constitución de la inflorescencia (trofotagma y unidad de floración), que resultó ser una aparente panícula.
Según Arciniegas-Torres y Flórez-Delgado (2018), los sistemas silvopastoriles constituyen una solución factible para la ganadería con enfoque de sostenibilidad, por lo que representan una opción viable para la conservación y adaptación al cambio climático, pues permiten intensificar la producción ganadera mediante la optimización y uso racional de los recursos naturales.
Dado que Ch. subaristatum forma parte de los sistemas silvopastoriles de la provincia de Entre Ríos, Argentina, se hace útil su estudio, a fin de promover y conservar la presencia de esta especie en el pastizal. Desde esta perspectiva, el objetivo de la investigación fue caracterizar aspectos morfológicos y reproductivos de Ch. subaristatum, en la provincia de Entre Ríos, Argentina.
Materiales y Métodos
Localización. Para las evaluaciones previstas, se cultivaron 15 plantas que provenían de cariopsis cosechados de noviembre a diciembre de 2015, en Oro Verde, provincia de Entre Ríos, Argentina (Galussi et al., 2016). Esta región se halla ubicada a 31º 50’ 37,31” S - 60º 32’ 27,92” O.
Procedimiento experimental. La siembra se realizó en papel de germinación (ISTA, 2014), una vez obtenidas las plántulas, con longitud de 5 cm en la parte aérea. En un ambiente protegido, se trasplantaron en macetas con tierra mullida, en las que se mantuvo el sustrato húmedo. Posteriormente, las macetas con las plantas se llevaron al medio exterior, con riego permanente.
Al inicio de la primavera, durante dos años consecutivos (2016 y 2017), se evaluaron 10 plantas (las mismas en los dos años). De ellas se tomaron datos del inicio de la floración, la madurez y el momento de cosecha, el número de inflorescencias, de nudos y paracladios, el número de espiguillas, de antecios con cariopsis y la germinación de los mismos.
Para ambas cosechas, las inflorescencias se cortaron despúes de que el raquis central cambiara de color, de verde amarillento a castaño claro, y se colocaron en sobres de papel abierto. Una vez producida la desarticulación de los antecios, se determinó el peso, la humedad y la germinación,, según ISTA (2014; 2018), con algunas modificaciones (Galussi et al., 2016).
Los antecios con cariopsis se almacenaron en el laboratorio, en sobres de papel, a temperatura ambiente. Se evaluó la germinación en períodos posteriores, según el año de cosecha, en tres repeticiones de 100 cariopsis (con lema y palea) a 25 °C, sobre papel de germinación (ISTA 2014; 2018). En una tercera cosecha de las mismas plantas, solo se registró el número de inflorescencias, y en antecios con cariopsis se atendió al contenido de humedad, se determinó el contenido de humedad, el peso y la germinación.
Análisis estadístico. Se calcularon los estadísticos promedio, desviación estándar y coeficiente de variación. Los datos, relativos al número de nudos y antecios, se compararon por medio de la prueba de diferencias de medias para muestras independientes. Se aplicó la prueba chi cuadrado para la homogeneidad de proporciones (Di Rienzo et al, 2016).
Resultados y Discusión
Para los dos años evaluados, la inflorescencia, desde el nudo de la hoja bandera hasta su ápice, alcanzó de 40 a 47 cm de longitud total, con poca variabilidad según las plantas y los años, característica referida por Burkart (1969). El eje principal de la inflorescencia (trofotagma más la unidad de floración) que porta a los paracladios tuvo de 100 a 160 mm de longitud. Se observaron de seis a nueve nudos, entrenudos proximales de 30 a 35 mm y distales de 5 mm. Los paracladios que integran la inflorescencia fueron de primer, segundo y tercer orden, en número de 11 a 17 los de primer orden para ambas cosechas. Los paracladios, de origen mayormente profilar, pueden ser largos (40 mm), intermedios (25 mm) o cortos (15 mm). Los de segundo y tercer orden fueron menos (1 a 4) y mostraron longitud inferior.
Se observó que la aparición de los paracladios del trofotagma, de primer, segundo y tercer orden, es acrópeta, siendo igual la antesis y maduración de las cariopsis en las espiguillas. El cambio de color de las espiguillas, de verde claro a amarillento amarronado, es expresión del grado de madurez del disemínulo. Las espiguillas se mostraron comprimidas, con antecios múltiples, de 8 a 10 como promedio, dispuestas dísticamente y fértiles.
Las observaciones mostraron que al madurar los antecios, se desarticulan de las espiguillas. Este tipo de desarticulación es basípeta, lo que se puede deber al menor tamaño de las cariopsis superiores, que conlleva a que el secado sea mucho más rápido.
En la figura 1 se muestran las características de los antecios y las cariopsis, que coinciden con las referidas por Calderón-de-Rzedowski y Rzedowski (2001), y permiten describir la lema. Se trata de una estructura de 1,8 a 3,5 mm de largo por 2,5 a 6,0 mm de ancho, subtrilobada, engrosada en el dorso, con margen membranáceo dilatado. Tiene un ápice agudo o acuminado, mucronado o brevemente aristado, pálea ovalada a suborbicular, de 1 a 2 mm de largo y de color amarillento claro. Su forma es alagrimada o acorazonada, dorsiventralmente comprimida. Los frutos semillas son cariopsis, de 1,0 a 1,5 mm de largo, dorsiventralmente comprimidos, con un embrión puntiforme recubierto por finas glumelas.

En las 10 plantas observadas, que tenían de siete a nueve meses de edad (cosecha de 2016), el número de inflorescencias en la primera floración (principios de septiembre) fue de 112, y en la segunda (mediados de octubre) de 59, para 65 y 35 %, respectivamente, con relación al total de inflorescencias por planta (17 inflorescencias como promedio). En tanto, para la cosecha de 2017, en las mismas plantas, con edad de 19 a 21 meses, de un total de 180 inflorescencias, la primera floración (principios de septiembre) representó 7 %, y la segunda, a los 55 días posteriores significó 23 %, para un total de 18 inflorescencias por planta como promedio. En la tercera cosecha (2018), en las mismas plantas y cantidad, pero de 21 a 33 meses de edad, se contabilizaron 268 inflorescencias, para 26 inflorescencias por planta como promedio.
En la primera floración de ambas cosechas, de las 10 inflorescencias registradas por cosecha, el número de espiguillas varió entre 24 y 58, según la inflorescencia y de acuerdo con el número de nudos y paracladios. El promedio de antecios con cariopsis fue de 314 y 290, y la variabilidad relativa registrada, de 20,3 y 16,4 para las cosechas de 2016 y 2017, respectivamente. El menor y el mayor número de antecios con cariopsis fue de 207 y 391, y de 208 y 365, para la cosecha de 2016 y 2017, respectivamente. En total se cosecharon 3 139 y 3 246 antecios, con cariopsis en primera y segunda cosecha, respectivamente, lo que depende de las condiciones ambientales, del tamaño de la planta, número de inflorescencias y espiguillas con antecios fértiles.
El número de nudos y antecios fértiles en la inflorescencia, según cada cosecha (2016 y 2017), se muestra en la tabla 1. Con relación al número de nudos, en la primera cosecha (2016), el valor medio (6,8) fue comparativamente menor al de la segunda (8,9), y presentó además menor variabilidad relativa (9,3 con respecto a 9,8). En cuanto al número de antecios fértiles (con semillas) por inflorescencia, el valor promedio para la primera cosecha (313,9) fue mayor que el de la segunda (290,1). Sin embargo, la variabilidad relativa resultó menor en esta última con respecto a la primera (16,4 de 20,3). Se encontraron diferencias estadísticas significativas (p ≤ 0,001) en el número de nudos entre cosechas. En la cosecha de 2017, el número de nudos fue significativamente mayor que en la de 2016. Con respecto a los antecios fértiles, no se encontraron diferencias estadísticas (p = 0,3574).
Tabla 1 Nudos y antecios con semillas en inflorescencias de Ch. subaristatum en dos cosechas.
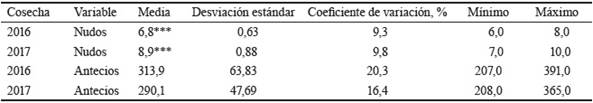
***p ≤ 0,001
Las inflorescencias (aparente panícula) están constituidas por paracladios (ramas floríferas) de distintas longitudes (largo, intermedio y corto), característica referida por Galussi et al. (2016). Los paracladios largos se ubican cerca de la base de la inflorescencia, después los intermedios y distalmente los cortos. Los resultados del análisis comparativo entre años se muestran en la tabla 2.
Tabla 2 Longitud de los paracladios con inflorescencias en Ch. subaristatum (mm)
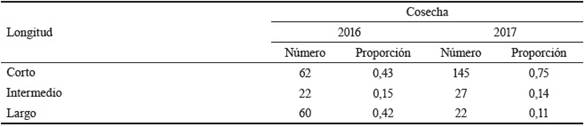
n= 10 inflorescencias de 10 plantas
La proporción de inflorescencias en paracladios intermedios entre años no se afectó (0,15 vs 0,14). En la segunda cosecha, hubo incremento de inflorescencias en paracladios cortos (0,43 a 0,75) en detrimento de los largos (0,42 a 0,11).
Finalizada la maduración, entre los 40 y 45 días, y al momento de la desarticulación de los disemínulos, el contenido de humedad, peso y germinación de los antecios con cariopsis fue variable, según años de cosecha (tabla 3). En los atributos humedad y peso, hubo baja variabilidad entre repeticiones de la misma muestra, aunque se registraron diferencias significativas entre los promedios de las muestras de cada año (p ≤ 0,05).

La germinación promedio entre años fue diferente al inicio y a través del tiempo, fluctuando con el mecanismo de dormancia, aunque de buena longevidad (tabla 4). La germinación de los disemínulos con cariopsis, 25 días posteriores a la cosecha de 2016, fue de 1 %, y presentó porcentaje elevado de semillas frescas. Transcurrido el tiempo, las semillas de la misma muestra manifestaron aumento de la germinación, que alcanzó 88 % de plántulas normales a los 240 días de almacenamiento. Posteriormente disminuyó de manera paulatina hasta 67 %, a los 780 días de almacenamiento.
Tabla 4 Germinación de antecios con semillas de Ch. subaristatum según período de almacenamiento (%).

En plantas del segundo año, las cariopsis cosechadas y sembradas 25 días posteriores a la cosecha (2017), tuvieron un comportamiento diferente con respecto a las del año anterior. Presentaron elevada germinación inicial (87 %), con variabilidad en el tiempo. Igualmente, su máxima germinación la manifestaron a los 240 días, período similar al pico máximo alcanzado en las semillas de la primera cosecha. Este comportamiento evidenció un estado de dormancia esporádica en esta especie.
Los ejemplares de Ch. subaristatum, de la misma edad y año de cosecha, presentaron escasa variabilidad en cantidad y tamaño de inflorescencias, paracladios y cantidad de antecios fértiles. La morfología estructural de la inflorescencia, en las plantas del segundo año, fue de características similares a las citadas para el primer año de cosecha por Galussi et al. (2016), con ligeras variaciones en longitud y características cuantitativas.
Entre las plantas de la primera y la segunda cosecha, el porcentaje de inflorescencias aumentó en 37 %, lo que sugiere más cantidad de macollos reproductivos en plantas de mayor edad. Aunque en un estudio del comportamiento en dos gramíneas C3, Chicahuala et al. (2018) sugirieron que el inicio del ciclo reproductivo estaría controlado por factores externos, poco variables entre años (fotoperíodo), mientras que su final podría estar regulado, en mayor medida, por factores con mayor variabilidad interanual (precipitaciones y temperaturas).
En este estudio, estos factores no se midieron, y podrían ser la causa de la posible variabilidad en la producción de semillas, además de la edad de las plantas. La cantidad de semillas por planta alcanzó potencialmente hasta 7 820 disemínulos. En otra gramínea, C3: raigrás (Lolium spp.) se han registrado, en condiciones de campo, en la región pampeana, producciones de 1 500 a 7 500 semillas por planta, en dependencia de la sensibilidad a glifosato y de las condiciones hídricas (Yanniccari et al., 2016). Potencialmente, la producción por planta en dicha especie puede llegar hasta las 12 mil semillas (Pop et al., 2010).
El peso, contenido de humedad y germinación de las cariopsis cosechadas de Ch. subaristatum presentaron diferencias significativas entre años de cosecha. El comportamiento registrado en la germinación mostró cierta coincidencia con lo referido para esta especie (Postulka et al., 2002) y para otras gramíneas estivales (Bolaños et al., 2015). Se confirma la ruptura de la dormancia innata sin tratamiento alguno, después de ocho meses de almacenamiento.
Las evidencias halladas en este estudio permiten afirmar la presencia de dormancia fisiológica no profunda o intermedia, según sea el ambiente de maduración. Según informan Baskin y Baskin (1998), esta se puede encontrar en gramíneas, y puede ser superada principalmente por almacenamiento en seco o aplicación de giberelina. No obstante, según lo observado en este experimento, las semillas pueden entrar en dormancia nuevamente después de la pérdida de esta condición. Hubo años, incluso, que después de la cosecha no presentaron dormancia, manifestándose después de algún tiempo de almacenamiento con la conservación de sus embriones viables. Por ello, se sugiere realizar a las semillas el ensayo de viabilidad por tetrazolio (ISTA, 2018), aunque se debe estandarizar el patrón para semillas de la especie, viables y no viables. Este comportamiento indicaría que la evaluación de la germinación no debe ser puntual sino periódica.
Conclusiones
Con respecto a la relación de la inflorescencia con los aspectos productivos, cosecha y calidad de las semillas, se sugiere realizar un manejo apropiado del pastizal, y no pastorear en el momento de desarrollo de los macollos reproductivos.
En la región de Entre Ríos, la producción de semillas es posible, y varía según la conformación de la inflorescencia, edad de las plantas y año de cosecha. Resulta conveniente hacer la cosecha previa a la desarticulación de los disemínulos.
Las semillas pueden presentar dormición esporádica, desde el inicio de la cosecha o durante el almacenamiento, lo que indica que la evaluación de la germinación debe ser periódica, y resulta conveniente sembrar después de que se rompa la latencia en la mayor cantidad de semillas.

