










 Servicios personalizados
Servicios personalizados Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La biota edáfica comprende los organismos que pasan una parte o todo su ciclo de vida en el interior del suelo o sobre su superficie inmediata. Incluye artrópodos, nemátodos, moluscos, entre otros. De acuerdo con el tamaño de su cuerpo, la fauna edáfica se clasifica en microfauna, mesofauna y macrofauna (Mekonen, 2019).
La edafofauna realiza múltiples servicios ambientales provechosos para el bienestar y la salud humana: descomposición de la materia orgánica, suministro de nutrientes para las plantas, mantenimiento de la estructura del suelo, movimiento y retención de agua en el perfil edáfico, control biológico de plagas y enfermedades y secuestro y liberación de carbono (Tanjung et al., 2020).
Las comunidades de la macrofauna a menudo se consideran como bioindicadores de la calidad del suelo, por ser sensibles a los cambios ambientales que pueden ocasionar variación en su abundancia y composición (Machado-Cuellar et al., 2020; Morel y Ortiz-Acosta, 2022). La calidad del suelo depende de propiedades físicas, químicas y biológicas. De acuerdo con su variabilidad espacial y temporal, sensibilidad a los cambios de uso del suelo y facilidad en su interpretación y ejecución, se pueden utilizar como indicadores de calidad.
El estudio de la relación entre las propiedades físicas y químicas del suelo, su intensidad de uso y la abundancia y diversidad de la biota edáfica, se utiliza para seleccionar indicadores de calidad, por lo que constituye un tópico actual en las investigaciones acerca de la ecología del suelo (Díaz-Porres et al., 2014). En Cuba, se han identificado varios grupos de la macrofauna edáfica como indicadores de la calidad del suelo. No obstante, se recomienda para fortalecer las acciones de investigación, validación y generalización de resultados en el país, extender la utilización de los indicadores ya generados a distintos suelos y ecosistemas. (Cabrera-Dávila et al., 2022). El objetivo de este estudio fue determinar la relación existente entre algunas propiedades químicas, físicas y biológicas del suelo y el componente vegetal, en agroecosistemas de pastizales de la provincia Granma, Cuba.
Materiales y Métodos
Localización. La investigación se desarrolló en cinco agroecosistemas de pastizales de la provincia Granma, ubicada en la porción suroeste de la región oriental de Cuba, entre las coordenadas 20°23′00″N y 76°39′09″O. En la tabla 1 se muestran las principales características de los agroecosistemas. Los muestreos se realizaron dos veces al año, en el período lluvioso, (PLL) y en el período poco lluvioso (PPLL), desde julio de 2014 hasta marzo de 2017.
Tabla 1 Principales características de los agroecosistemas.
| Agroecosistema | Triángulo y Progreso | Cupeycito | Ojo de agua | Estación de pastos |
|---|---|---|---|---|
| Municipio | Bayamo | Jiguaní | Guisa | Bayamo |
| Afiliación | UBPC Francisco Suárez Soa | Empresa Genética Manuel Fajardo | Finca de Rafael Almaguer, CCS Braulio Coroneaux | Jorge Dimitrov |
| Propósito | Producción de leche | Cría de terneros | Ceba de toros | Ceba de toros |
| Tipo de suelo | Vertisol pélico | Pardo mullido carbonatado | Pardo mullido carbonatado | Fluvisol |
| Método de pastoreo | Continuo | Rotacional | Continuo | Rotacional |
| Área total de pastoreo, ha | T:18,5 P: 20,4 | 14,2 | 6,7 | 0,8 |
| Área de muestreo, ha y porcentaje que representa del área total | T: 2 11 % P: 2 10 % | 1,8 13 % | 1,2 18 % | 0,8 100 % |
| Tipo de pastos predominantes |
|
|
|
Sistema silvopastoril de hierba de |
| Tiempo de explotación, años | 20 | 10 | 7 | 10 |
| Raza y carga animal, UGM ha-1 | Mestizo Siboney 1,5 | Criollo 1,7 | Mestizo 2,2 | Mestizo Siboney 1 |
| Condiciones generales | Área de pastoreo totalmente deforestada, sin cuartones. Se encharca en la época de lluvia | Buen nivel de sombra por árboles y acuartonamiento, alta pedregosidad. Especies de árboles: |
Buen nivel de sombra por árboles, sin cuartones, relieve con pendiente (10 %). Susceptibilidad a la erosión. Especies de árboles: |
Buen nivel de sombra, zona de intensa sequía |
T: El Triángulo P: El Progreso, UBPC: Unidad básica de producción cooperativa, CCS: Cooperativa de crédito y servicio
Muestreo e identificación de la macrofauna y mesofauna edáficas. Se utilizaron dos métodos: el recomendado por el programa Tropical Soil Biology and Fertility (Anderson e Ingram, 1993) y las trampas de caída o pitfall (Moreira et al., 2012). Para el primer método, se limpió la hojarasca previamente y se extrajo todo tipo de cuerpos extraños, como piedras y residuos vegetales. En la diagonal del área de muestreo, se extrajeron cinco monolitos por hectárea, de 25 x 25 x 20 cm, a una distancia de 20 m. Manualmente se recolectaron y contaron los individuos de la macrofauna in situ. Las lombrices se conservaron en formaldehido al 4 % y los invertebrados restantes en etanol al 70 %.
Para el segundo método de muestreo, se colocaron en cada área de estudio nueve trampas, dispuestas en las dos diagonales en forma de cruz, con una trampa en el centro. Se utilizaron recipientes plásticos de 8 cm de diámetro y 10 cm de profundidad, los que se enterraron a ras del suelo, con el menor disturbio posible en el área circundante. Después se añadió una solución acuosa detergente al 0,003 %, preparada con detergente líquido comercial de LABIOFAM y se taparon con hojas secas y restos vegetales propios de cada agroecosistema. Al cabo de siete días, se colectó el contenido de las trampas en frascos de cristal y se trasladaron al laboratorio. Con la utilización del estereoscopio se extrajeron y contaron los individuos de la solución y se colocaron en viales con etanol al 70 %.
Para la identificación de los especímenes conservados se consultaron los trabajos de Hickman et al. (2008) y los de Brusca y Brusca (2003). También se revisó la colección entomológica perteneciente al Laboratorio Provincial de Sanidad Vegetal en Granma. Se definieron las variables en los monolitos y en las trampas de caída: número de individuos pertenecientes a los órdenes Araneae, Hymenoptera, Hymenoptera-Formicidae, Coleoptera, Isopoda, Hemiptera y Haplotaxida.
La mesofauna edáfica se determinó en las trampas de caída. Después de aclarar los individuos con hidróxido de sodio al 5 % y pasarlos por lactofenol con un ligero calentamiento, se montaron finalmente en líquido FOR o Hoyer para su clasificación. La identificación taxonómica se realizó según los trabajos de Brusca y Brusca (2003) y Díaz-Azpiazu et al. (2004). Se definieron las variables número de ácaros y colémbolos.
Microflora. Se tomaron cinco muestras compuestas por diez submuestras, a profundidad de 0-20 cm, con ayuda de una piqueta. Se utilizó el método de las diluciones seriadas y siembra profunda en placas Petri (Mayea et al., 1998). Se definieron como variables el número de unidades formadoras de colonias (UFC) de bacterias, hongos y actinomicetos.
Las condiciones de aislamiento de la microflora se muestran en la tabla 2.
Tabla 2 Condiciones de aislamiento de la microflora.
| Grupo microbiano | Dilución | Medio de cultivo | Temperatura, ºC | Tiempo de incubación |
|---|---|---|---|---|
| Bacterias | 106 | Agar nutriente | 30 | 24 horas |
| Hongos | 104 | Agar extracto de malta | 30 | 5 a 7 días |
| Actinomicetos | 105 | Agar almidón amoniacal | 30 | 5 a 7 días |
Determinación de las propiedades químicas y físicas del suelo. Para la determinación de los indicadores químicos y físicos (tabla 3), se tomaron cinco muestras compuestas (diez submuestras tomadas en zig-zag) en toda el área de muestreo, a profundidad de 0-20 cm, con la ayuda de una barrena helicoidal. Estas muestras se secaron al aire y posteriormente se molinaron y tamizaron (1 mm) para la realización de los análisis agroquímicos. Para la determinación de la densidad aparente se tomaron muestras de suelo no disturbado.
Tabla 3 Propiedades físicas y químicas del suelo. Métodos para su determinación.
| Indicador | Métodos |
|---|---|
| pH (H2O) (1:2) | Potenciometría, NC ISO 13.080.10 :2015 |
| MO, % | NC ISO-51, 1999 |
| Na, K, Ca, Mg¥ | Extracción con AcNH4, por el método de Maslova |
| Na, K, cmol kg-1 | Determinación por fotometría de llama |
| Ca, Mg, cmol kg-1 | Determinación por volumetría con EDTA |
| P2O5, ppm | Oniani (1964) |
| CE, dS m-1 | Conductimétrico NC 112:2001 |
| Composición granulométrica y microestructura, % | NRAG 408, 1981 |
| Humedad higroscópica, % | Método gravimétrico NC 110, 2001 |
| Densidad aparente, g cm-3 | NRAG 370, 1980 |
| Densidad real, g cm-3 | NC 11 508, 2000 |
| Tamizado en seco y estabilidad estructural, % | Método de Savinov (Orellana |
| Coeficiente de estructura en seco | Cálculo |
| Porosidad total | Pt = (1- Da/Dr)*100 |
| Carbono orgánico, % | CO=MO*0,58 |
NC: Norma cubana, NRAG: Norma ramal agrícola, ¥ Cationes intercambiables
Determinación de la composición botánica y la producción de biomasa. La composición botánica se determinó en 80 marcos ha-1, distribuidos al azar, en cada área de muestreo mediante el método de t´Mannetje y Haydock (1963). La composición botánica se estimó a partir del valor de importancia relativa, que estuvo en función de la frecuencia y dominancia relativa del pasto, las plantas arvenses y el suelo desnudo o la despoblación, considerada por la ausencia de vegetación.
La disponibilidad de biomasa se determinó en cada área, en 100 marcos de 0,25 m2, tomados al azar según la metodología de Haydock y Shaw (1975). Los patrones de muestras del pasto se cortaron a altura aproximada de 10 cm. Se definieron cuatro variables: porcentaje de pastos, de arvenses y suelo desnudo y producción de biomasa (t ha-1).
Análisis estadístico. Se realizaron análisis de correlaciones canónicas entre los grupos de variables de la macrofauna edáfica en los monolitos y en las trampas de caída. La mesofauna y la microflora se determinaron con las variables físicas y químicas del suelo y las variables del componente vegetal definidas anteriormente. Este análisis se realizó para dos grupos de variables en cada caso. Se utilizaron dos criterios de significación de las relaciones canónicas, de acuerdo con Badii et al. (2007): el nivel de significación estadística de las funciones (p < 0,05) y la magnitud de la correlación canónica (Rcanónico > 0,75). El análisis se realizó con el paquete estadístico Statistica V 8.0 para Windows (Statsoft, 2008).
Resultados y Discusión
Correlaciones canónicas de las variables de la biota edáfica con las propiedades físicas y químicas de los suelos. El análisis de correlación canónica fue significativo entre las variables de la macrofauna y la microflora edáficas y las propiedades físicas y químicas del suelo (tabla 4). Al analizar la estructura de los factores obtenidos en la correlación canónica se observó que, en todos los casos, el orden Isopoda estuvo entre las variables de la macrofauna edáfica que más aportó a la varianza compartida por ambos grupos de variables. Para la macrofauna determinada por el método de los monolitos, se le adicionó el orden Haplotaxida, que solo se observó por este método. Lo anterior permite inferir que en los agroecosistemas objeto de estudio estos órdenes son los que más se relacionan con las propiedades físicas y químicas de estos suelos. No existió correlación canónica significativa de las variables de la mesofauna edáfica con las propiedades físicas y químicas del suelo.
Tabla 4 Correlaciones canónicas entre variables de la biota edáfica y las propiedades físicas y químicas de los suelos.
| Grupo 1 | Grupo 2 | R canónico | χ2 | p | Variables que más contribuyen |
|---|---|---|---|---|---|
| Macrofauna-t | Físicos | 0,89 | 67,01 | 0,008 | Isopoda, Dr (-), Da |
| Macrofauna-t | Químicos | 0,92 | 77,83 | 0,004 | Isopoda, Mg, pH (-) |
| Macrofauna-m | Físicos | 0,89 | 69,36 | 0,005 | Isopoda, Haplotaxida, Hy (-), Arcilla (-) |
| Macrofauna-m | Químicos | 0,91 | 72,95 | 0,012 | Haplotaxida, Isopoda, Ca, K |
| Microflora | Físicos | 0,99 | 130,87 | 0,000 | Bacterias, Hy (-) |
| Microflora | Químicos | 0,99 | 137,86 | 0,000 | Actinomicetos, Na, Ca |
Macrofauna-t: macrofauna capturada en las trampas; Macrofauna-m: macrofauna capturada en los monolitos
Hubo diferencias en cuanto a las variables físicas y químicas de mayor contribución a la varianza cuando se realizó el análisis con la macrofauna, determinada por el método de los monolitos y por las trampas de caída. En la macrofauna que se determinó por el método de las trampas, hubo influencia positiva del Mg y la Da, y por el pH y la Dr de forma negativa. En la macrofauna determinada por el método de los monolitos, se relacionó positivamente con el Ca, K y, de forma negativa, con la Hy y el porcentaje de arcilla. Esto se pudiera deber a las diferencias de la macrofauna colectada en ambos métodos, pues la fauna con características de mayor movilidad (actividad diurna o nocturna), se captura más fácil mediante las trampas; mientras que los monolitos concentran su acción en aquellos organismos menos móviles, con actividad diurna fundamentalmente (Chávez, 2020).
Las figuras 1 y 2 muestran las correlaciones simples más importantes entre los organismos de la biota edáfica y las variables físicas y químicas de los suelos. El orden Isopoda se relacionó de forma positiva con la Dr, la humedad higroscópica y la porosidad total y, de forma negativa, con la Da, el Ca y el Mg. Los organismos pertenecientes a Haplotaxida se relacionan positivamente con la humedad higroscópica y el K y, de forma negativa, con el porcentaje de arcilla. Todas las relaciones de las bacterias fueron negativas con respecto a la estabilidad estructural, el porcentaje de arcilla, la humedad higroscópica y el Ca y Na.
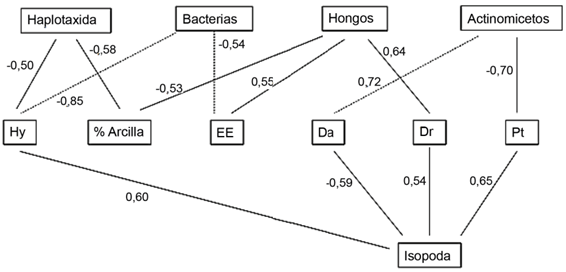
Fig. 1 Correlaciones entre la biota edáfica y variables físicas del suelo (Hy: humedad higroscópica, EE: estabilidad estructural, Da: densidad aparente, Dr: densidad real, Pt: porosidad total).
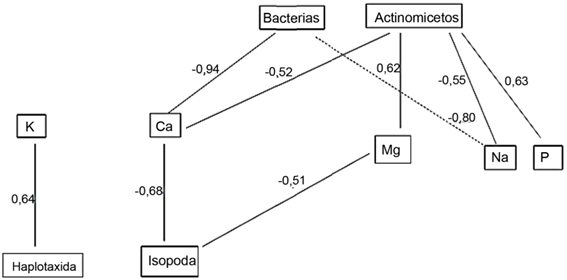
Los hongos solo se relacionaron con las variables físicas de forma positiva (estabilidad estructural y Dr); mientras que los actinomicetos solo se relacionaron con las variables químicas (de forma negativa con el Ca y el Na, y positivamente con el Mg y el P).
La distribución de la macrofauna del suelo depende de varios factores: tipo de suelo, contenido de nutrientes, materia orgánica, pH, textura y estructura (Pollierer et al., 2021 ). Además, son muy importantes los factores relacionados con la vegetación, el clima, el uso del suelo y el manejo antrópico desarrollado (Tanjung et al., 2020; Valkay-Halkova et al., 2022). Las correlaciones entre la biota edáfica y las propiedades físicas y químicas de los suelos halladas en el presente estudio han sido informadas por otros autores. Díaz-Porres et al. (2014) encontraron correlación significativa entre las variables físicas y químicas del suelo y la densidad de grupos taxonómicos de la macrofauna edáfica. Estos autores concluyeron que, al analizar la estructura de los factores obtenidos en la correlación canónica, la relación C/N y el porcentaje de materia orgánica fueron las variables edáficas que mejor explicaron la distribución de los macroartrópodos. Ello difiere de los resultados presentados, pues estos factores no fueron los de mayor contribución.
Desde el punto de vista de las correlaciones simples, también se registraron resultados similares a lo revisado en la literatura. Rosa et al. (2015) comprobaron que las propiedades químicas del suelo que mejor se correlacionaron con los grupos de la macrofauna edáfica fueron la materia orgánica, el Ca, el Mg, el P y el K, cuando realizaron un estudio que incluyó pastizales, en Santa Catarina, Brazil. Gholami et al. (2016) informaron también un resultado coincidente, al constatar una correlación negativa del contenido de arcilla y la conductividad eléctrica, con algunos índices de la macrofauna edáfica (abundancia, uniformidad, riqueza y diversidad) en un estudio realizado en un bosque al suroeste de Irán. Estos índices tuvieron correlación positiva con el contenido de materia orgánica y de limo. Hani y Suhaendah (2019) señalaron que los factores que influyeron en la diversidad de la macrofauna del suelo fueron el pH, la temperatura y la humedad del suelo. Ge et al. (2021) distinguieron al carbono orgánico como el principal factor entre las propiedades del suelo, el cual determina la composición de la macrofauna edáfica.
Li et al. (2020) informaron que los grupos de la biota Bourletiella (Collembola), Symphyla y Armadillidae (Isopoda) estuvieron influenciados principalmente por el contenido de K. En tanto, las larvas de Staphylinidae (Coleoptera) se afectaron fundamentalmente por la temperatura del suelo. Otros grupos, como Muscidae (Diptera), Chironomid, Psychodidae, y Scydmaenidae (Coleoptera), mostraron influencia mínima de los factores del suelo.
Tulande et al. (2018) constataron igualmente que la macrofauna mostró respuesta positiva al contenido de P, K y Na del suelo, en los Andes colombianos. Otros autores informaron que el P y el Na determinan la presencia de algunos grupos de la macrofauna edáfica, como Araneae, Coleoptera, Isopoda y Haplotaxida (Ott et al., 2014). Sin embargo, en campos de arroz en Indonesia, Tanjung et al. (2020) establecieron que la disponibilidad de P no influyó en el incremento de la diversidad de los grupos de la macrofauna edáfica.
En un estudio realizado en sabanas naturales con pastoreo en los llanos venezolanos, Morales-Márquez et al. (2018) observaron que la macrofauna edáfica se correlacionó positivamente con la porosidad del suelo y de forma negativa con la densidad aparente, como sucedió con el orden Isopoda en esta investigación (fig. 1).
Varias investigaciones en regiones tropicales y subtropicales han relacionado directamente, a escala local, la distribución espacial, densidad y biomasa de las lombrices de tierra y comunidades de la macrofauna del suelo con el contenido de materia orgánica, nitrógeno y otros nutrientes, relación C/N, pH, textura (contenido de arena y arcilla), capacidad de intercambio catiónico, retención de agua, aireación, porosidad y estructura del suelo (Rodríguez-Suárez et al., 2019; Sofo et al., 2020; Panklang et al., 2022).
La literatura evidencia la importancia de los nutrientes del suelo para el desarrollo de la macrofauna edáfica, como el calcio, que interviene en varios mecanismos de regulación osmótica en los invertebrados, así como en los procesos de crecimiento (Rosa et al., 2015). Mientras, en los oligoquetos, forma parte del arreglo espacial de las subunidades polipeptídicas de la hemoglobina (Moreira et al., 2011).
Según De-la-Cruz-Lozano (2005), el potasio y el fósforo pueden ser factores limitantes del crecimiento de los insectos y el magnesio es esencial como cofactor enzimático.
Si se analiza la mayor contribución de los órdenes Haplotaxida e Isopoda a los factores obtenidos en la correlación canónica junto a Dr, Da, Hy, contenido de arcilla, pH, Ca, K, Mg y Na, también se encuentran elementos que coinciden con la literatura. La dependencia de las lombrices de tierra de la textura, densidad real, porosidad y carbono orgánico, manifiesta cambios de composición y abundancia en una corta escala de tiempo (Zhukova y Mytiai, 2022). Según estos autores, las lombrices de tierra prevalecen en ambientes edáficos húmedos, no compactados y con alto contenido de materia orgánica.
Rodríguez (2020) en un estudio con varios niveles de diversificación e intensificación en la rotación de cultivos en Argentina, constató que la abundancia y la biomasa de las lombrices de tierra se relacionaron de forma positiva, fundamentalmente con el contenido de materia orgánica. De igual forma, Hoeffner et al. (2021), en 24 pastizales de Francia, pudieron comprobar que el incremento de la materia orgánica fue el factor más favorable para la abundancia y la biomasa de las lombrices. Nanganoa et al. (2019) registraron una correlación fuerte y negativa del pH y el número de lombrices en cinco usos intensivos de la tierra en el trópico húmedo de Camerún.
También se reconoce que, organismos detritívoros, como los pertenecientes al orden Isopoda, son muy sensibles a los cambios físicos y químicos del suelo, así como a los cambios bruscos de temperatura y humedad en sus hábitats, por lo que se pueden utilizar para indicar el estado de perturbación en el medio edáfico. Para estos organismos es indispensable la cobertura vegetal diversa, pues trae consigo una hojarasca más heterogénea y mayor concentración de macronutrientes en el suelo. Al respecto, Isopoda se ha asociado con una mayor cobertura vegetal, materia orgánica y humedad del suelo (Cabrera-Dávila, 2019). Otros autores enfatizaron la importancia de Isopoda en la descomposición de la hojarasca y la interacción con los microorganismos del suelo (Pey et al., 2019).
Correlaciones canónicas entre variables de la biota edáfica. Entre los diferentes grupos de la biota edáfica, también se establecieron correlaciones canónicas significativas. Este es el caso de la macrofauna determinada por las trampas y la mesofauna (Rcanónico = 0,79; χ2 = 29,12; p = 0,0038). Aquí, los coleópteros y ácaros fueron las variables que más influyeron negativamente. La microflora (bacterias, hongos, actinomicetos) se correlacionó con la macrofauna determinada por los monolitos (R canónico=0,95; χ2=29,12; p=0,0099). El número de bacterias y el orden Isopoda fueron las variables que tuvieron mayor contribución negativa a la varianza.
Las correlaciones individuales mostraron que las bacterias se relacionaron positivamente con el orden Isopoda, Hemiptera, Haplotaxida y con los actinomicetos, que tuvieron una relación positiva con los hongos (fig. 3).
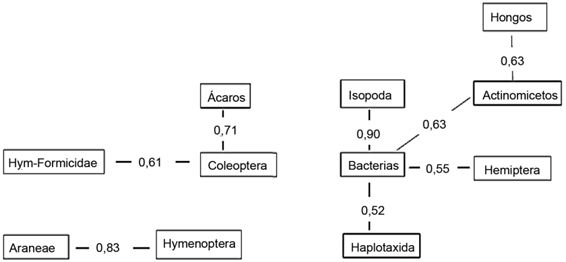
Varios grupos de la biota se relacionaron entre sí: ácaros y coleópteros, arañas e himenópteros, hormigas y coleópteros, presumiblemente por su similar movilidad. Resulta interesante que todas las relaciones que se establecieron entre los componentes de la biota edáfica fueron positivas. Díaz-Porres et al. (2014) encontraron también este tipo de correlación entre Araneae y Coleoptera y otros grupos de la macrofauna edáfica: Isopoda con Hemiptera y Lepidoptera con Chilopoda.
Correlaciones canónicas entre las variables de la biota edáfica y las variables del componente vegetal. Las variables de la biota edáfica que mostraron correlaciones canónicas significativas con las variables del componente vegetal fueron la macrofauna determinada por los monolitos (Rcanónico = 0,89; χ2 = 54,39; p = 0,00038) y la microflora (Rcanónico = 0,93; χ2 = 59,71; p = 0,0000). De las primeras, las variables que más contribuyeron a la varianza fueron el porcentaje de despoblación y el orden Haplotaxida. De las segundas, el porcentaje de arvenses y los hongos demostraron mayor contribución. Las bacterias y los individuos pertenecientes a Isopoda se relacionaron de forma positiva con la producción de biomasa y el porcentaje de pastos (fig. 4). Los actinomicetos y las bacterias se relacionaron negativamente con el porcentaje de arvenses. Mientras que, los hongos lo hicieron de forma positiva; al parecer los suelos con mayor cobertura de arvenses fueron los que brindaron mejores condiciones para estos microorganismos y, a su vez, se relacionaron negativamente con el porcentaje de pastos.
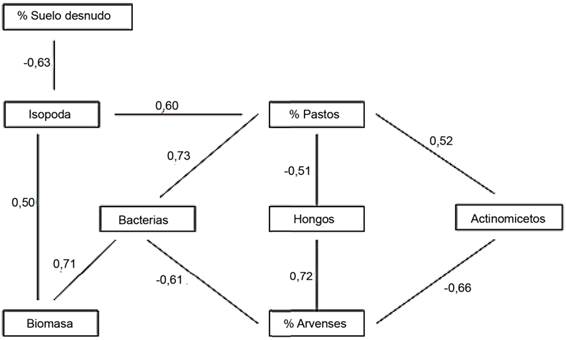
Se ha informado ampliamente en la literatura que la diversidad de la vegetación, así como la cobertura del suelo, se asocian con la diversidad y la abundancia de la fauna edáfica (Lo-Sardo y Silva, 2019; Sabatté et al., 2021; Panklang et al., 2022). Lo anterior avala los resultados que mostraron la relación entre las variables del componente vegetal y la macrofauna, determinada por los monolitos y la microflora.
Rodríguez-Suárez et al. (2018) en la evaluación de la macrofauna edáfica en usos diferentes de la tierra de la Amazonía colombiana, incluidos pastizales y sistemas silvopastoriles, refirieron que el análisis de componentes principales separó al bosque natural como el uso de la tierra con mayor diversidad de la macrofauna, donde abundaron los órdenes Isopoda y Gastropoda. Según estos autores, los resultados indican que la mayor diversidad de árboles puede resultar en mayor heterogeneidad y cantidad de hojarasca y, consecuentemente, mayor disponibilidad de alimentos y microhábitats, así como mejores condiciones edáficas y microclimáticas para el desarrollo de la macrofauna.
Rodríguez (2020) señaló que la abundancia y la biomasa de las lombrices de tierra se relacionaron positivamente con los índices intensidad de la rotación y proporción de leguminosas en los agroecosistemas de Argentina. Singh et al. (2021) informaron que, en Alemania, en diferentes tipos de pastizales, las comunidades de lombrices se afectaron significativamente. La menor abundancia y biomasa se pudo ver en el pastizal sometido a uso intensivo, debido a la menor diversidad de plantas presentes. Guaca et al. (2019) refirieron efecto significativo de la vegetación (bosques, plantación boscosa, silvopastoreo y pastizal) en las propiedades microbiológicas del suelo en la Amazonía colombiana.
Selección de indicadores. Los órdenes de la macrofauna edáfica Isopoda y Haplotaxida se seleccionaron como indicadores de buena calidad del medio edáfico, pues según el análisis de correlaciones canónicas fueron los que tuvieron mayor relación con las propiedades físicas y químicas de los suelos, así como con las variables del componente vegetal en los pastizales estudiados. Además, como indicadores biológicos cumplen determinadas características que los avalan para este fin: gran aptitud para la especiación, ciclo de vida corto, poco poder de dispersión por su adaptación a la vida edáfica y a diferentes tipos de suelo, hábitos alimentarios relacionados con la degradación de la materia orgánica y respuesta predecible a los cambios en el ambiente (Cabrera-Dávila, 2019). Estos organismos son fáciles de colectar e identificar, pues se pueden ver a simple vista. Además, no se necesita equipamiento de laboratorio, ni reactivos, ni personal especializado para su determinación, lo que constituye una limitación para los indicadores físicos y químicos.
Varios estudios sugieren el uso de las lombrices de tierra como indicadores de la calidad del suelo (Mekonen, 2019; Siebert et al., 2019; Rodríguez, 2020). En Cuba, Cabrera-Dávila (2019) propuso como indicador faunístico la relación lombrices/hormigas. En los usos de la tierra con mayor cobertura del suelo, como es el caso de los bosques, esta relación fue mayor que uno; mientras que en sistemas de uso con mayor grado de disturbio los valores fueron próximos a cero. Los autores señalan la utilidad práctica de este indicador, pues los grupos involucrados son de fácil identificación y no requieren conocimiento especializado. De igual forma, se definen las familias pertenecientes a Isopoda como taxones indicadores: Philosciidae en bosques primarios y Trachelipidae y Armadillidae, en sistemas agroforestales.
No obstante, Ramírez et al. (2019) reconocen que la identificación y estudio de estos organismos como bioindicadores de la calidad del suelo y de la biodiversidad de los ecosistemas permanece como un problema universal, y que las comunidades de los macro-invertebrados varían en su composición, abundancia y riqueza, en dependencia del estado de perturbación que causa el cambio de uso de suelo. Por tanto, se trata de indicadores de la perturbación e impacto de las diferentes formas de manejo.
Conclusiones
Existieron correlaciones canónicas significativas entre las propiedades físicas y químicas del suelo con la biota edáfica, y de esta con el componente vegetal y entre las propias variables biológicas. A partir de estas últimas fue posible sugerir relaciones entre estos elementos en los agroecosistemas de pastizales estudiados, las que ayudan a explicar la dinámica de estos organismos del suelo con las propiedades físicas y químicas y con el componente vegetal. Se proponen los órdenes Haplotaxida e Isopoda como indicadores de calidad del suelo, seleccionados y validados a partir de los análisis de correlaciones canónicas entre las propiedades edáficas físicas y químicas y el componente vegetal. A esto se adiciona, la facilidad en su muestreo e identificación, posible de realizar por productores, investigadores y otros especialistas.

