










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión On-line ISSN 2224-4697
Rev. Protección Veg. v.22 n.1 La Habana ene.-abr. 2007
Trabajo original
CARACTERIZACIÓN MOLECULAR DE AISLADOS DE CAMPO DEL VIRUS DEL ENCRESPAMIENTO AMARILLO DE LA HOJA DEL TOMATE (TYLCV)
MOLECULAR CHARACTERIZATION OF FIELD ISOLATES OF TOMATO YELLOW LEAF CURL VIRUS (TYLCV)
Madelaine Quiñones, D. Fonseca y Yamila Martínez
Grupo de Fitopatología, Dirección de Protección de Plantas, Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, La Habana. Cuba. Correo electrónico: madeqp@censa.edu.cu
RESUMEN
Los begomovirus constituyen «virus emergentes» de plantas. Debido a las infecciones persistentes en los campos de tomate, las pérdidas ocasionadas por estos virus a la agricultura mundial y a la gran variabilidad informada para muchos de sus miembros, por lo que el objetivo de este trabajo fue caracterizar la variabilidad genética de la población natural del TYLCV predominante en Cuba. Para esto, se secuenciaron 21 aislados de campo del TYLCV, obtenidos de diferentes años y regiones del país. Se utilizaron cebadores específicos para la amplificación y secuenciación de la región intergénica (RI) y del extremo 5` terminal del gen c1 o proteína de la replicación (rep). Las secuencias obtenidas se analizaron mediante programas bioinformáticos para determinar las relaciones filogenéticas con otros aislados de TYLCV informadas en bases de datos y predicción de posibles estructuras secundarias. Los resultados evidenciaron la presencia de una única población del TYLCV en el país, donde todos los aislamientos mostraron entre 97-99% de identidad con el TYLCV previamente aislado en Cuba. Se determinó la presencia de tres estructuras secundarias diferentes con elevada estabilidad termodinámica, donde la estructura predominante es similar a la determinada para el TYLCV aislado en Egipto y diferente al TYLCV-Isr previamente aislado en el país. Los resultados obtenidos en este trabajo aportan nuevos elementos para la caracterización de la diversidad genética y el control del TYLCV; los que pudieran explicar las posibles causas de la amplia diseminación de este virus observada en los campos de tomate cubanos.
Palabras clave: TYLCV; begomovirus; tomate; variabilidad genética; PCR; secuenciación; estructura secundaria.
ABSTRACT
Begomoviruses are plant emergent viruses. Due to the persistent TYLCV infections in tomato fields, the losses caused by theses viruses in the worldwide agriculture and the high variability reported for most of their members, the objective of this work was to characterize the genetic variability of the natural population of TYLCV in Cuba. Twenty one isolates of TYLCV, collected from several years and regions of the country, were amplified and sequenced using specific primers that amplify the intergenic region (IR) and the 5` end of c1 gene or replication protein (rep). The sequences obtained were analyzed by bioinformatic softwares to determine the phylogenetic relationship with other TYLCV isolates reported in Genkbank, and predict possible secondary structures. Results evidenced the presence of an only one TYLCV population in the country, where all the isolates showed between 97-99% identity of their nucleotide sequences with that TYLCV previously isolated in Cuba. The presence of three different secondary structures with high thermodynamic stability was determined, being the predominant structure similar to that of TYLCV-Egypt and different from that of TYLCV-Isr. These results provide new elements for the characterization of the genetic diversity and control of TYLCV, and could explain the possible causes of the wide dissemination of the virus observed in the Cuban tomato fields.
Key words: TYLCV; begomoviruses; tomato; genetic variability; PCR; sequencing; secondary structure.
INTRODUCCIÓN
Los begomovirus, pertenecientes a la familia Geminiviridae, constituyen el principal problema fitosanitario del cultivo del tomate (Lycopersicon esculentum Mill)en las regiones tropicales y subtropicales (3). La presencia de miembros de este género, como el virus del chino del tomate (CdTV) en México y el virus del encrespamiento amarillo de la hoja del tomate (TYLCV) en la mayoría de los países de la cuenca del Caribe, han provocado pérdidas de hasta un 100% de las cosechas (15). En Cuba, se han informado pérdidas entre el 20-100% en este cultivo debido al azote de plagas y enfermedades, dentro de estos los begomovirus constituye uno de los principales problemas identificados (6, 23).
A inicios de la década de los 90´ la elevada incidencia de estos virus hizo necesario la puesta en marcha de un programa de manejo integrado del complejo mosca blanca-geminivirus y la ejecución de proyectos de investigación encaminados a la identificación, caracterización y obtención de métodos de diagnóstico para la detección del agente causal. Como resultado de estos proyectos se identificaron en el país tres begomovirus en el cultivo del tomate: el virus monopartito del encrespamiento amarillo de la hoja del tomate identificado en Israel (TYLCV-Isr) (13), los virus bipartitos taíno moteado del tomate (ToMoTV) (24) y mosaico Habana del tomate (ToMHV) (14).
Los estudios de distribución de begomovirus en áreas de la provincia de La Habana mostraron la presencia del TYLCV, ToMHV y de infecciones mixtas de estos virus en las principales variedades comerciales en producción, donde la infección por el TYLCV resultó ser la de mayor distribución (12). Recientes informes muestran una predominancia de este virus en varios países (República Dominicana, Jamaica, Estados Unidos y España), el cual ha sido capaz de desplazar a los virus endémicos y se ha convertido en la mayor causa de pérdidas de la agricultura de estos países (2, 17, 27, 28), el que infectó no solo el tomate, sino otros cultivos de importancia económica, como el frijol (Phaseolus vulgaris) y pimiento (Capsicum annuum) (17, 22, 25, 27), además de malezas encontradas frecuentemente en las áreas de producción de muchos cultivos de interés agrícola (29).
Por otra parte, los resultados obtenidos a partir de los estudios de evolución y variabilidad de las poblaciones de virus de ADN, son pocos hasta la fecha, y en el caso específico de los begomovirus han sido muy escasos. En la actualidad los trabajos realizados con este fin se basan fundamentalmente en el estudio de las regiones intergénica (RI) y el extremo 5´ terminal del gen c1 o proteina rep de los begomovirus, por su función la replicación de estos virus y la información taxonómica que estas ofrecen para la caracterización de nuevos aislados (5,17). Estos resultados, unidos a la amplia diversidad de los begomovirus presentes en toda la región de América Latina y el Caribe, así como a los recientes informes de emergencia de muchos de los miembros de esta familia y el alto potencial de recombinación informado, para dar lugar a nuevos virus, en algunos casos de mayor severidad, conllevaron a la necesidad de profundizar en el estudio de estos patógenos. El objetivo de este trabajo ha sido caracterizar la variabilidad genética de la población natural del TYLCV-Isr en Cuba, mediante la amplificación, secuenciación y análisis molecular de la región intergénica (RI) y el extremo 5` terminal del gen de la replicasa viral de nuevos aislados de campo colectados. Estos resultados aportan nuevos elementos cognoscitivos de la enfermedad en el país y son de gran valor para el manejo y control de esta entidad.
MATERIALES Y MÉTODOS
Selección de los aislados del TYLCV-Isr presente en Cuba
Con el objetivo de determinar la variabilidad genética dentro de la población natural del TYLCV-Isr presente en Cuba se seleccionaron 21 aislados del virus. Para esta selección se tuvieron en cuenta los siguientes aspectos: año de colecta, región de procedencia y la variedad de tomate en que fue aislado el virus (Tabla 1).
Reacción en Cadena de la Polimerasa (PCR)
Los ADN seleccionados se analizaron mediante la PCR para lo cual se amplificó la región intergénica y el fragmento correspondiente al extremo 5´ terminal del gen rep del TYLCV-Isr con los cebadores IRTY2353+ (análogo a la posición 2353 de la cadena viral) / IRTY 255- (complementario a la posición 255 de esta secuencia) descritos previamente (22). Las reacciones de amplificación se ajustaron a un volumen final de 50 mL, se utilizó 1U de la enzima Taq ADN Polimerasa en la solución amortiguadora suministrada por la casa comercial (Promega) y suplementado con 200 mM de cada dNTP, 2 mM de MgCl2 (Promega), 0.4 mM de cada cebador y 100ng de la muestra.
Las amplificaciones se realizaron en un termociclador (MJ Research, Inc.). El programa de amplificación consistió en un paso inicial de desnaturalización a 950C durante 5 min, seguida por 35 ciclos de reacción (30s a 950C de desnaturalización, 30 s a 600C de anillamiento de los cebadores y 30 s a 720C de extensión), seguido por un paso de extensión final durante 7 min a 720 C. Los productos de la PCR se analizaron en geles de agarosa al 1%, se utilizó como patrón de peso molecular el marcador de 1Kb (GIBCO) (29).
La banda de interés de cada producto de la PCR (684pb) se purificó, mediante el juego de reactivos Gene Clean II de acuerdo a las condiciones recomendadas por la casa comercial (Bio101. Inc) y la concentración de los fragmentos purificados se determinó mediante la lectura de la D.O a 260 nm en un espectrofótometro LassosSpec III (Lasso Biotech, LTDA) (29).
Análisis de las secuencias nucleotídicas
Los productos obtenidos en la PCR se secuenciaron. Para esto se utilizaron los cebadores específicos IRTY2353+/ IRTY 255- anteriormente descritos. La secuenciación se realizó automáticamente mediante el uso del sistema "ABI PRISM Dye terminator", con el juego de lectura de ciclos de secuencia con la enzima ADN polimerasa (AmpliTaq DNA polymerase FS, Perkin Elmer) de acuerdo con las condiciones recomendadas por la casa comercial. Las reacciones de secuenciación se prepararon según el protocolo adjunto al juego de reactivos. Las secuencias y el cromatograma obtenido para las mismas se analizaron mediante el programa informático Chromas (versión 1.62) (http://www.technelysium.com.au/Chromas.html).
Para el análisis y comparación de las secuencias de la RI se escogieron de forma representativa 11 de los nuevos aislados de campo, mientras que para el estudio del extremo 5' Terminal del gen c1 se seleccionaron nueve aislados. Las secuencias nucleotídicas se compararon con las de distintos aislamientos del TYLCV-Isr (TYLCV-Cu (Cuba); TYLCV-Jp (Japón); TYLCV- Isr (Israel); TYLCV-Esp (España); TYLCV-Port (Portugal); TYLCV- RD (Rep Dominicana), así como de otros begomovirus relacionados (TYLCV-Sar (TYLCV aislado en Cerdeña, TMoHV, TToMV) informadas en la base de datos NCBI (http://www.ncbi.nlm.nih.gov/blast/blast.cgi ). El alineamiento de las secuencias se realizó con el programa CLUSTAL W (versión 1.75) (30) y se calculó la matriz de distancias evolutivas de acuerdo al método de Kimura (11). El árbol filogenético se construyó por el método del vecino más cercano (28) con el programa "Windows Easy Tree" (WET, versión 1.31) (4) y se realizó un remuestreo para 1000 replicaciones.
Predicción de las estructuras secundarias del extremo 5´ de la RI
La predicción de las estructuras secundarias óptimas de los nucleótidos (1-200) del extremo 5´ de la RI de los nuevos aislados se realizó con el programa MFOLD (versión 3.25) del paquete GCG (32) (http://www.bioinfo.math.rpi.edu /mfold) a 37°C, y estas se compararon con la obtenida para el TYLCV-Isr y los aislados de República Dominicana, Egipto e Irán informados en la base de datos del NCBI.
RESULTADOS Y DISCUSIÓN
Variabilidad de la población natural del TYLCV-Isr en el cultivo del tomate en Cuba
Al comparar la secuencia completa de los fragmentos de 684pb de los 21 aislamientos cubanos del TYLCV-Isr, se separaron los alineamientos correspondientes a la región 5´ terminal de la rep y la secuencia de la región intergénica. El alineamiento de las secuencias de la región 5' terminal del gen rep de los aislados secuenciados y su comparación con la misma región del TYLCV aislado previamente en Cuba (13) mostró entre 95 y 99% de identidad. Mientras que al comparar las secuencias de la RI de estos nuevos aislamientos con la del TYLCV aislado previamente mostró entre ellos porcentajes de homología desde 96 hasta 100%. Estos resultados evidencian la poca divergencia encontrada entre las secuencias de los aislados estudiados y el TYLCV-Isr previamente identificado en el país (13) y de acuerdo a los criterios taxonómicos establecidos (21) sugieren la presencia de un único aislamiento del TYLCV-Isr en el cultivo del tomate en Cuba. Las observaciones hechas para el virus del mosaico amarillo dorado del frijol (BGYMV) en Brasil apoyan estos resultados, donde los estudios de comparación de secuencias de nuevos aislados con los previamente informados mostraron porcentajes de homología entre el 93 y 100% (5).
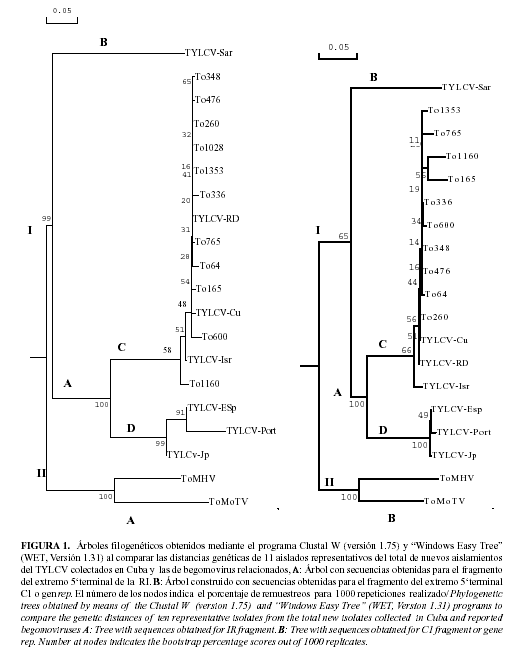
La Figura 1 (A y B) muestra las relaciones filogenéticas obtenidas al realizar el alineamiento de los fragmentos de la RI y del extremo 5' de la rep, de 10 de las aislados secuenciados.
Puede observarse (Fig. 1A), que los aislados comparados se separan en dos grupos diferentes (I y II); el primero agrupa a los aislados del TYLCV (aislado de Israel, los aislados caribeños del mismo (TYLCV-Cu y TYLCV-RD) y los aislados de España (TYLCV-Esp), Portugal (TYLCV-Port), Japón (TYLCV-Jp) y Cerdeña (TYLCV-Sar). Mientras que en el grupo II se encuentran los begomovirus bipartitos identificados en Cuba en los últimos años (TMoHV; TToMV). Dentro del grupo I se distinguen dos subgrupos (A y B), donde se integran en el subgrupo A los aislados del TYLCV de Israel, Cuba, República Dominicana, España, Portugal y Japón junto con los nuevos aislados cubanos en estudio. Sin embargo el TYLCV-Sar se separa independiente en el subgrupo B. Por otra parte, el subgrupo A se divide en dos nuevos subgrupos (C y D), encontrándose en el C los aislados del TYLCV caribeños y en el D los aislados de España, Portugal y Japón, resultados estos que coinciden totalmente con lo esperado.
La comparación de las secuencias del extremo 5´ del gen rep (Fig. 1B) mostró resultados similares a los obtenidos al analizar los fragmentos de la RI secuenciadas de los diferentes aislados y comparadas con las secuencias de referencia utilizadas en el trabajo. De igual forma el análisis filogenético mostró la presencia de dos grandes grupos (I y II). El grupo I une a todos los aislados de TYLCV comparados y en el grupo II se integran los begomovirus bipartitos aislados en Cuba.
Estos resultados confirman que las secuencias analizadas en este trabajo constituyen nuevos aislados del TYLCV-Isr, ya que son idénticos al resto de los aislados del virus utilizados como referencia en este trabajo y evidencian una rápida diseminación de este virus en el país previo a su introducción a inicios de la década del 90, a través, posiblemente del intercambio de material vegetal con estas Islas del Caribe o con Israel (15,12).
Comparaciones más detalladas de la secuencia de nucleótidos del extremo 5´ de la RI mostraron que en 11 aislados (To 336, To 165, To 418, To 476, To 600, To 1023, To 1160, To 933, To 227, To 64 y To 729) se observaron pequeñas sustituciones de nucleótidos a lo largo de las secuencias analizadas al compararlos con la secuencia del TYLCV previamente aislado en Cuba. No obstante, la secuencia de nonanucleótidos (TAATATTAC) típica de esta familia viral, capaz de formar una estructura de tallo-lazo y ubicada entre los nucleótidos 138-170 de la RI, está perfectamente conservada en todos los aislados, tal como ha sido informado para todos los geminivirus caracterizados hasta la fecha (9,12,31). Estos resultados son similares a los obtenidos por Faria y Maxwell (5) al caracterizar la región RI del virus del mosaico dorado amarillo del frijol (BGYMV) en Brasil.
Los cambios más significativos en estas secuencias ocurrieron en el aislado To227, donde se observó una sustitución del nucleótido T por C en la posición 151 (extremo 5' de la RI y cercano al tallo de la secuencia de nonanucleótidos conservada). Mientras que en los aislados To 600, To 933, To1160 y To1023 ocurrió una sustitución del nucleótido G por C en la posición 180 de esta secuencia (en la zona del tallo posterior a la secuencia de nonanucleótidos). De igual forma los aislados To 729 (G por A) y To 227 (T por C) mostraron sustituciones de nucleótidos en la zona del tallo adyacente a la secuencia de nonanucleótidos conservada. En los aislados To64, To336, To 418 y To165, también se observaron pequeños cambios en otras regiones de menor importancia dentro del extremo 5´ de la RI.
Por otra parte, la comparación de las secuencias repetidas o iterones (ATCGGTGATCGGTG) (motivos supuestos específicos de unión con la proteína rep), ubicados de forma adyacente a la secuencia TATA a partir de las posiciones 11, 38 y 46 del extremo 5`terminal de la cadena viral y a partir de la posición 82 (en dirección contraria a la cadena viral) (12) mostró que la localización de estos concuerda con lo informado para el TYLCV-Isr aislado en el país previamente (13). Los motivos localizados en la dirección de síntesis de la cadena viral se mantuvieron conservados en todos los aislados analizados, sin embargo no ocurrió lo mismo con el motivo detectado en la dirección contraria a la cadena viral donde se observó una sustitución del nucleótido C por una T en el aislado To 476. Un cambio en este iterón, puede influir en la secuencia de los aminoácidos (aa) conservados de la proteína rep, que actúan como sitio de acoplamiento del ADN en el proceso de replicación, tal como ha sido planteado para este y otros iterones (1, 26). Por esta razón, los cambios de nucleótidos encontrados en algunas de las secuencias obtenidas para la RI sugirieron un análisis más detallado de las secuencias obtenidas para los fragmentos rep de estos nuevos aislados.
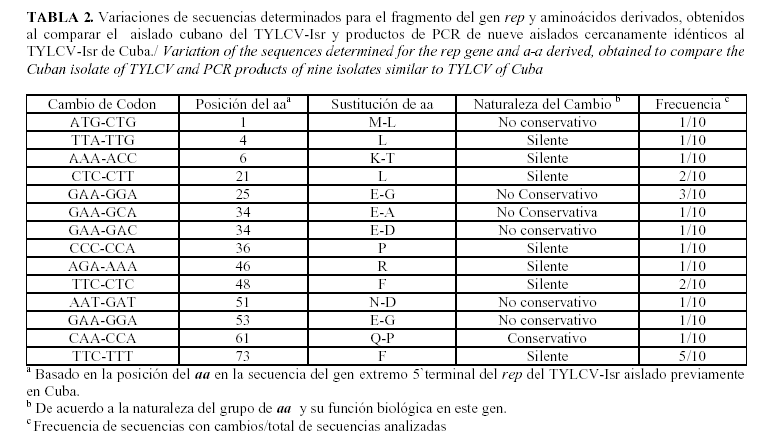
En la Tabla 2 se observan las sustituciones de nucleótidos y sus residuos de aminoácidos (aa), detectados al alinear las secuencias de nucleótidos y de proteínas del extremo 5´ terminal del gen rep del aislado cubano del TYLCV-Isr y el fragmento correspondiente a este gen en nueve de los aislados estudiados. En cinco de los aislamientos estudiados (To 1023, To 1353, To 165, To 765, To933) se encontraron sustituciones de nucleótidos. De forma general, se observaron 22 sustituciones de nucleótidos en las secuencias de estos aislados. De estas, sólo una conllevó a una sustitución de aminoácidos conservativa que se manifestó en el aislado (To 933) y se localizó en el motivo II (HLHxUUQ) de esta proteína, el cual es conservado para la mayoría de los geminivirus estudiados (19). Este cambio implicó la sustitución del aa Q (residuo de glutamina) por el P (residuo de prolina). Sin embargo, algunos autores plantean la importancia de la presencia en esa posición de un aa hidrofóbico, tal como sucede en este aislado, donde aparece la sustitución por prolina (18). En este motivo se mantiene además la presencia de dos histidinas que están involucradas con los iones metálicos requeridos para el corte del ADN por la proteína iniciadora de la replicación.
De las restantes sustituciones nueve resultaron no conservativas, ya que estas no implicaron cambios de gran importancia que puedan repercutir en la función biológica de este gen y 12 fueron sustituciones silentes que no conllevaron cambios en la secuencia de aminoácidos. Las sustituciones ocurridas (conservativas y no conservativas) en la secuencia 5' del gen rep, están presentes en la misma posición de todos los aislamientos en que se observaron estas, por lo que las mismas no pueden ser atribuidos a mutaciones puntuales ocurridas en este fragmento durante la reacción de amplificación y en la lectura de la secuencia.
Los motivos I (FLTYP) y III (xxYxxK) (10) están absolutamente conservados en todos los aislados. Sin embargo, el aislado To476 mostró un cambio en la secuencia de aa de la zona estructural M..X4,X3,X2,X 1,FX1L*X3AKNY FLTYPQC, que constituye el mayor elemento de reconocimiento del origen de replicación (1). Esta divergencia encontrada puede determinar cambios en la preferencia del ADN para iniciar la replicación en este aislamiento en particular.
Predicción de Estructuras Secundarias
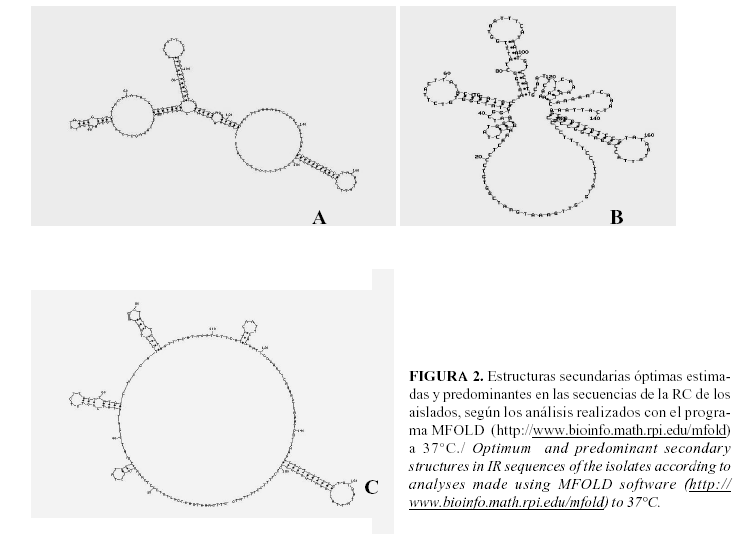
El análisis de predicción de la estructura secundaria óptima de los primeros 200 nucleótidos del extremo 5´ de la RI de los aislamientos (Fig. 2) permitió agruparlos en 3 estructuras óptimas diferentes (A, B y C). La estructura de la Fig. 2A correspondió al aislado cubano del TYLCV-Isr con una estabilidad termodinámica de DG= -20.2 Kcal/mol a 370C. Con estructura similar estuvo el aislamiento To476, que a pesar de presentar pequeños cambios, los cuales provocaron una ligera disminución de su estabilidad termodinámica (DG= -18.3 Kcal/mol), es el único que posee una estructura similar al TYLCV-Isr caracterizado anteriormente.
Los aislados To1160 y To 64 mostraron una estructura (Fig. 2B) con valores DG de -16.7 y -18 Kcal/mol, respectivamente, ligeramente similar a la obtenida para el aislado de TYLCV de República Dominicana, que mostró DG=-20 Kcal/mol. Los resultados obtenidos con el análisis de la estructura óptima confirmaron que las divergencias encontradas en las comparaciones filogenéticas de estos aislados repercuten en la estructura secundaria de la RI, con valores de energía libre que indican la posibilidad de que sean aislados más inestables en el campo.
El resto de los aislados mostró una estructura óptima con DG entre -15,4 y -19 Kcal/mol, la que se observa en la Figura 2C. La estructura obtenida para estos aislados fue similar a la del TYLCV- Eg y TYLCV-IR (DG=-20.8 y -20.2 Kcal/mol). Los aislamientos To1360, To1028, To1353, To260, To1023, To1118, To933 y To227 presentaron la misma estructura pero con cambios en los valores de energía libre (DG = -15.4 Kcal/mol). Los aislados To765 y To600 tuvieron valores de DG entre -17.2 y -17.7 Kcal/mol. La estructura predominante determinada está presente en aislados del año 1995 y se continúa manifestando en la mayoría de los aislados colectados hasta la fecha, lo que indica que a pesar de presentar una estabilidad termodinámica inferior a la obtenida para el TYLCV-Isr aislado en Cuba y la de otros aislados del TYLCV; esta se ha mantenido en el tiempo, lo que pudiera ser indicio de la adaptación de una nueva variante biológica al hospedante, que pudiera llegar a ser tan importante como lo es el aislado cubano del TYLCV-Isr.
La estructura secundaria óptima determinada por este programa confirmó que la mayoría de los aislados colectados muestran la estructura de la Fig. 2 C. Esta estructura posee un lazo grande, alrededor del cual se ubican varios tallos-lazos, donde el mayor de estos contiene la secuencia nonámera conservada en todos los geminivirus. La estructura obtenida para estos es diferente a la encontrada para el TYLCV previamente identificado en Cuba (Fig. 2 A), en la cual el lazo grande es muy reducido y presenta el tallo-lazo característico en la misma posición, lo cual coincide con lo descrito para los geminivirus (20).
En los análisis realizados se pudo determinar que cambios por encima de un 20% de la energía libre con respecto al aislado cubano del TYLCV-Isr, provocaron alteraciones en la estructura secundaria óptima. Resultados similares han sido descritos por otros autores que sólo después de un 10% de diferencia en la DG, han observado variaciones en la estructura secundaria (7,8). Por su parte, Navas-Castillo et al. (16), al caracterizar los aislados de TYLCV de España y Portugal determinaron una DG=-16.1 Kcal/mol al compararlos con el aislado leve del TYLCV-Isr, y observaron un aumento de la energía libre (DG =-6.8 Kcal/mol) y por tanto una disminución de la estabilidad interna del aislado. Estos resultados pudieran estar relacionados con una posible implicación de estas mutaciones en el incompleto apareamiento de bases en esta zona, considerada fundamental para la replicación viral, lo cual hace pensar que la ligera severidad detectada para este aislado podría deberse a la presencia de una estructura de tallo-lazo alterada. Los resultados obtenidos muestran la presencia de tres estructuras diferentes en los aislados caracterizados, lo cual pudiera estar relacionado con la gran explosión que ha tenido la problemática de los begomovirus, fundamentalmente del TYLCV en Cuba, en los últimos años. Los mismos sientan las bases para profundizar en el mecanismo de interacción entre la replicasa viral y estas estructuras.
Por otra parte, estos resultados evidencian la presencia de una única población circulante del TYLCV en las principales áreas de producción de tomate en el país. La razón de la presencia de una única población viral mantenida en el tiempo de estudio y en todas las áreas de producción del país, podría ser explicada por el uso en la actualidad de variedades e híbridos parcialmente resistentes, las cuales podrían crear una presión de selección hacia los aislados más agresivos del virus.
El presente trabajo constituye el primer estudio de variabilidad de las poblaciones naturales del TYLCV-Isr en Cuba y para la región de América Latina y el Caribe. Aporta además elementos cognoscitivos en el estudio de esta entidad viral y sienta las bases para futuros estudios de interacción hospedante-patógeno. Así mismo, la escasa variabilidad del TYLCV-Isr encontrada hasta la fecha podría beneficiar enormemente la estrategia a seguir en los programas de mejoramiento genético para resistencia a la enfermedad causada por el TYLCV en la obtención de nuevas variedades e híbridos por métodos tradicionales y alternativas de ingeniería genética con secuencias derivadas del patógeno.
AGRADECIMIENTOS
Los autores agradecen al Ministerio de Asuntos Exteriores de Italia por la concesión de la beca en la cual se realizó parte de este trabajo y al Dr. Gian Paolo Accotto (Instituto de Fitovirología Aplicada, Turín, Italia) en la dirección y apoyo para ejecutar el trabajo de secuenciación de genes.
REFERENCIAS
1. Argüello-Astorga GR, Ruiz-Medrano R. An interon- related domain is associated to motif 1 in the replication proteins of geminivirus: identification of potential interacting amino acid-base pairs by a comparative approach. Arch Virol. 2001;(146):1465-1485.
2. Czosnek H, Laterrot H. A worldwide survey of tomato yellow leaf curl viruses. Arch Virol. 1997;(142):1391-1406.
3. Dopazo J. A new index to find regions showing an unexpected variability or conservation in sequence alignments. Comput Appl Biosci. 1997;13(3):313-317.
4. Faria JC, Maxwell DP. Variability in geminivirus isolated associated with Phaseolus spp. in Brazil. Phytopathology. 1999;(89):262-268.
5. Gómez O, Martínez Y, Piñón M. Nuevas líneas de Tomate resistentes al virus del encrespamiento amarillo de la Hoja (TYLCV), asintomáticas a la infección. Rev Protección Veg. 2002;12(2): 119.
6. González Lien. Caracterización y diagnóstico molecular del Viroide del Tubérculo Ahusado de la Papa .PSTVd. en Cuba. Tesis presentada en opción al grado científico de Doctor en Ciencias Agrícolas.2000;102 p.
7. Gultyaev AP, Van Batenburg FHD, Pleij CWA. Dynamic competition between alternative structures in viroid RNAs simulated by an RNA folding algorithm. J Mol Biol. 1998;(276):43-55.
8. Gutiérrez C. Geminivirus DNA replication. Cell Mol Life Sci.1999;(56):313-329.
9. Hoogstraten RA, Hanson SF, Maxwell DP. Mutational Analysis of the Putative Nicking Motif in the Replication-Associated Protein AC1 of Bean Golden Mosaic Geminivirus. MPMI. 1996;9(7):594-599.
10.Kimura M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J Mol Evol. 1980;(16):111-120.
11.Martínez Yamila Contribución al conocimiento de geminivirus que afectan el cultivo del tomate. Lycopersicon esculentum Mill. en Cuba. Tesis en opción al grado de Doctor en Ciencias Agrícolas. CENSA, La Habana. 1998;119 p.
12.Martínez YM, Zabalgogeazcoa I, De Blas C, Sánchez F, Peralta EL, Ponz F, Romero J. Geminivirus associated with diseased tomatoes in Cuba. J Phytopathology. 1996;(144):277-279.
13.Martínez Y, De Blas C, Quiñones M, Castellanos C, Peralta EL, Romero J. Havana tomato virus, a new bipartite geminivirus infecting tomatoes in Cuba. Arch Virol. 1998;(143):1757-1772.
14.Morales FL, Anderson PK. The emergence and dissemination of whitefly-transmitted geminivirus in Latin America. Arch Virol. 2001;(146):415-441.
15.Navas-Castillo J, Sánchez-Campos S, Noris E, Louro D, Accotto GP, Moriones E. Natural recombination between tomato yellow leaf curl virus-IS and tomato leafcurl virus. J Gen Virol. 2000;(81):2797-2801.
16.Navas-Castillo J, Sánchez-Campos S, Díaz JA, Saez-Alonso E, Moriones E. Tomato Yellow leaf curl virus-Is cause a novel disease of common bean and severe epidemics in tomato in Spain. Plant Dis.1999;(83):29-32.
17.Orozco BM, Hanley-Bowdoin L. Conserved sequence and structural motifs contribute to the DNA binding and cleavage activities of a geminivirus replication protein. J Biol Chem. 1998;(273):448-456.
18.Orozco BM, Miller AB, Seattlage SB, Bowdoin HL. Functional Domains of a Geminivirus Replication Protein. J Biol Chem. 1997;272(15):9840-9846.
19.Orozco BM, Bowdoin HL. A DNA structure is required for geminivirus replication origin function. J Virol. 1996;70(1):148-158.
20.Padidam M, Beachy RN, Fauquet CM. Classification and identification of geminivirus using sequence comparison. J Gen Virol. 1995;(76):249-263.
21.Quiñones M, Fonseca D, Martínez Y, Accotto GP. First Report of Tomato Yellow Leaf Curl Virus (TYLCV)-infecting Pepper Plants in Cuba. Plant Dis. 2002;(86):73.
22.Quiñones M, Martínez Y, Fonseca D, Gómez O. Virus del Encrespamiento Amarillo de la Hoja del Tomate (TYLCV): Aportes a los estudios de variabilidad y diagnóstico para Cuba y la región de América Latina y el Caribe. En: 44th Reunión de la Sociedad de Fitopatología Americana-División Caribe. Ciudad Habana, Cuba. AP 3.7;2004.
23.Ramos PL, Guerra O, Peral R, Oramas P, Guevara RG, Rivera- Bustamante R. Taino tomato mottle virus, a new bipartite Geminivirus from Cuba. Plant Dis. 1997;(81):1095.
24.Reina J, Morilla G, Bejarano ER, Rodríguez MD, Janssen D, Cuadrado I M. First Report of Capsicum annuum Plants Infected by Tomato Yellow Leaf Curl Virus. Plant Dis. 1999(83):1176.
25.Roye ME, McLaughlin WA. Genetic diversity among Geminivirus associated with the weed species Sida spp., Macroptilium lathyroides, and Wissadula amplissima from Jamaica. Plant Dis. 1997;(81):1251-1258.
26.Roye ME, Wernecke WA, McLaughlin WA, Nakhla KM, Maxwell DP. Tomato dwarf leaf curl virus, a new bipartite geminivirus associated with tomatoes and peppers in Jamaica and mixed infection with tomato yellow leaf curl virus. Plant Pathol. 1999;(48):370-378.
27.Saito N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987;(4):406-425.
28.Salati R, Nahkla MK, Rojas MR, Guzmán P, Jaquez J, Maxwell DP, Gilbertson RL. Tomato yellow leaf curl virus in the Dominican Republic: Characterization of an infectious clone virus monitoring in whiteflies, and identification of reservoir hosts. Virology. 2002;92(5):487-496.
29.Sambrook J, Fritsch EF, Maniatis T. Molecular cloning. Second Edition. Cold Spring Harbor Laboratory Press, editor; 1989.
30.Thompson JD, Higgins DG, Gibson TJ, CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position specific gap penalties and weight matrix choice. Nucleic Acids Res.1994;(22):4673-4680.
31.Zhou X, Liu Y, Calvertz L, Muñoz C, Otim-Nape GW, Robinson DJ, Harrison BD. Evidence that DNA-A of a geminivirus associated with severe cassava mosaic disease in Uganda has arisen by interspecific recombination. J Gen Virol. 1997;(78):2101-2111.
32.Zuker M, Mathews DH, Turner DH. Algorithms and thermodynamics for RNA secondary structure prediction: a practical guide. En: RNA biochemestry and biotechnology. Barciszewski J, Clark BFC, editors. NATO AI Series, Kluwer Academic Publishers; 1999.
(Recibido 5-11-2005; Aceptado 6-10-2006)


{kind=link}
{kind=link}
{kind=link}
{kind=link}