










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión On-line ISSN 2224-4697
Rev. Protección Veg. v.22 n.2 La Habana Mayo-ago. 2007
ARTÍCULO RESEÑA
ESTRATEGIAS DE INGENIERÍA GENÉTICA PARA LA OBTENCIÓN DE PLANTAS RANSGÉNICAS RESISTENTES A GEMINIVIRUS. EXPERIENCIA DEL CENSA
STRATEGIES OF GENETIC ENGINEERING FOR THE OBTAINMENT OF TRANSGENIC PLANTS RESISTANT TO GEMINIVIRUSES. EXPERIENCE AT CENSA
Madelaine L. Quiñones*, A. Vega**, Yamila Martínez* y E. Rodríguez***
* Grupo de Fitopatología, Dirección de Protección de Plantas y ** Grupo de Virología Animal, Dirección de Microbiología. Centro Nacional de Sanidad Agropecuaria (CENSA). Apartado 10, San José de las Lajas, La Habana. Cuba. Correo electrónico: madeqp@censa.edu.cu; ***Facultad de Ciencias. Universidad de Málaga, España.
RESUMEN
Los begomovirus constituyen el principal problema para la producción del cultivo del tomate (Lycopersicon esculentum Mill.) en la mayoría de las regiones tropicales y subtropicales, causando pérdidas de un 100% del rendimiento. Para el control de los mismos se han aplicado varias estrategias convencionales de lucha, las que prometen obtener resultados alentadores a largo plazo. La aplicación de la transgenesis en plantas para conferir resistencia a virus hace de esta alternativa una herramienta de gran importancia para el desarrollo de una agricultura más sostenible. Entre las estrategias más utilizadas se encuentran: la resistencia conferida por la sobreexpresión del gen de la proteína de la cápsida viral (CP), uso de ácidos nucleicos defectivos interferentes, expresión de los ARNs de genes de interés utilizando la expresión de ARNs antisentido, así como de pequeños fragmentos de ARN viral como inductores del silenciamiento génico post-transcripcional. En Cuba, se han identificado tres begomovirus afectando este cultivo, el Virus del Encrespamiento Amarillo de la Hoja del Tomate (TYLCV), Taíno Moteado del Tomate (ToMoTV) y Mosaico Habana del Tomate (ToMHV), pero el TYLCV ha sido el de mayor diseminación e incidencia. Dado el impacto que conllevan las pérdidas ocasionadas por los geminivirus y el aporte de la ingeniería genéticapara su control, el presente artículo refiere aspectos sobre las principales estrategias desarrolladas para la obtención de plantas transgénicas resistentes a estos, el impacto de esta tecnología en la agricultura actual y los resultados específicos obtenidos en el CENSA.
Palabras clave: ingeniería genética; plantas transgénicas; resistencia derivada del patógeno; geminivirus.
ABSTRACT
Begomoviruses constitute the principal problem for the production of tomato crop (Lycopersicon esculentum Mill.) in tropical and subtropical regions, causing yield damages of 100%. Conventional strategies for the control of these viruses which promise to obtain long term results have been developed. The application of trangenesis in plants for conferring resistance to virus is a tool of great importance for the development of a more sustainable agriculture. Among the main strategies developed are: resistance by the over expression of the coat protein (CP), use of interfering defective nucleic acids, expression of RNAs from genes of interest, as well as the use of antisense RNAs and short RNAs as inductors of post-transcriptional gene silencing. In Cuba, three begomoviruses have been identified in tomato crops: Tomato Yellow Leaf Curl Virus (TYLCV), Tomato Mosaic Havana Virus (ToMHV) and Taíno Tomato Mottle Virus (TToMoV), but TYLCV has had the greatest incidence and dissemination. Due to the impact on damages caused by geminiviruses and the role played by the genetic engineering in their control, the present article reviews the aspect related to the main strategies developed for obtainment of transgenic plants resistant to geminiviruses, the impact of this technology in the current agriculture and the specific results obtained at CENSA.
Key words: genetic engineering; transgenic plants; pathogen-derived resistance; geminiviruses.
INTRODUCCIÓN
Las enfermedades virales causan pérdidas significativas en todos los cultivos de importancia económica, que estas afectan. Las mismas provocan un impacto negativo en la agricultura mundial, por lo que durante muchos siglos los agrónomos y fitopatólogos han realizado un considerable esfuerzo para controlar las mismas. Su control se basa fundamentalmente, en la detección a tiempo de la enfermedad para evitar el establecimiento y diseminación de sus agentes causales. En este sentido, la prevención mediante la evaluación frecuente de los semilleros y el uso de variedades con altos niveles de tolerancia o resistencia a virus constituyen los elementos de mayor importancia para su control. Para la prevención de estos patógenos se hace necesario el uso de un grupo de medidas de manejo integrado, que comprenden, desde la aplicación de prácticas culturales tradicionales, hasta la lucha contra el vector y otras posibles fuentes de virus, así como, la aplicación de nuevas tecnologías de avanzada que permitan de conjunto con las técnicas tradicionales realizar un control más eficiente de estas entidades patógenas (36).
Los geminivirus causan pérdidas devastadoras en varios cultivos de importancia económica, particularmente en los países tropicales y subtropicales (25,34). Estos se caracterizan por presentar partículas isométricas geminadas (del latín, gemini: dobles), variando su tamaño entre 30-36nm de largo y 18-20nm de ancho, teniendo como principales vectores, la mosca blanca y diferentes especies de saltahojas (Homoptera: Cicadellidae), además de algunas chinches de los árboles (Membraceae). Estos virus integran la segunda familia más extensa de virus de plantas y está compuesta por cuatro géneros: Mastrevirus, Curtovirus, Topocuvirus y Begomovirus que se distinguen por su vector, gama de hospedantes y características de su genoma. El genoma viral está compuesto por una cadena sencilla de ADN circular, de aproximadamente 2.7-2.8 Kb, que se presenta en algunos miembros como un solo componente (genoma monopartito) y en otros, como dos componentes (genoma bipartito). La mayoría de los begomovirus tienen dos componentes genómicos el ADN-A y ADN-B, ambos esenciales para la infectividad. El ADN-A presenta seis genes: Ac1 codifica para una proteína esencial para la replicación viral (Rep) en asociación con la ADN polimerasa del hospedante, Ac2 codifica una proteína activadora de la transcripción (TrAP), Ac3 codifica una proteína potenciadora de la replicación (REn), Av1 y Av2 codifican la proteínas de la cápsida y la proteína de pre-cápsida, respectivamente. La función del gene Ac4 aun no está completamente esclarecida. En el componente B se localizan los genes Bv1 y Bc1 que codifican proteínas relacionadas con el transporte nuclear y el movimiento, respectivamente. A diferencia los begomovirus monopartitas presentan un único componente genómico donde se localizan todos los genes esenciales para la replicación, transcripción, encapsidación y movimiento viral (43).
Tres begomovirus se han identificado en el cultivo del tomate (Lycopersicon esculentum Mill.) en el país: Virus del Encrespamiento Amarillo de la Hoja (de las siglas en inglés TYLCV) (22), Mosaico Habana (ToMHV) (23) y Moteado Taíno del Tomate (ToMoTV) (37). La alta incidencia del TYLCV constituye el principal problema para la producción de tomate en muchos países (46). Cuba no es una excepción, donde se ha podido cuantificar pérdidas desde el 40-100% en campos comerciales de tomate, además de haberse informado en los últimos años su presencia en nuevos hospedantes de importancia económica, tales como pimiento (34) frijol (26) y calabaza (24).
Antes del advenimiento de la ingeniería genética, las metodologías de mejoramiento tradicional fueron aplicadas al desarrollo de plantas resistentes a virus de cultivos importantes agronómicamente. En adición, técnicas estándares de fitopatología, que incluyen cuarentena, erradicación, rotación de cultivos y uso de semilla certificada libre de virus se han utilizado como herramientas de valor para el control de enfermedades virales.
Para el caso específico de los geminivirus, se han aplicado métodos de control tradicionales y de nueva generación, obteniéndose resultados alentadores en muchos casos (6,16,20,27,29). Estrategias no convencionales son desarrolladas en la actualidad en muchos laboratorios del mundo. Los avances en el campo de la biología molecular y la ingeniería genética, han permitido desde hace más de una década, la obtención de plantas transformadas que portan información genética foránea y les confiere resistencia a virus (6,10, 27,30).
Varios métodos que emplean el uso de la ingeniería genética han sido evaluados y demostrados satisfactoriamente para la obtención de plantas trangénicas resistentes a geminivirus (42). Entre los más relevantes se pueden mencionar, la sobreexpresión en plantas de genes virales (gen que codifica para la proteína de la cápsida cp) (4), uso de ácidos nucleicos defectivos interferentes (41) y la expresión de los ARN de genes de interés (rep, AV2) utilizando construcciones antisentido (6,30,31,32,33), así como un pequeño fragmento del ARN viral que desencadena el denominado mecanismo de silenciamiento génico post-transcripcional (de las siglas en inglés PTGS) (29,31).
En este sentido, la aplicación de estas estrategias han posibilitado en gran medida apoyar el control de estos patógenos de interés y obtener en menor tiempo; y de forma más específica variedades resistentes a geminivirus, cuyo proceso reduce a la mitad el tiempo necesario destinado a obtener una variedad mejorada por la vía clásica. No obstante, a pesar del desarrollo alcanzado, la aplicación e introducción de esta tecnología para obtener plantas con los caracteres deseados, aún continúa siendo un tema de gran debate a nivel internacional. Los riesgos asociados con la liberación de las plantas al medio ambiente natural que pudiera traer consigo el escape de los transgenes y la introgresión de estos en el pool de genes silvestres, el impacto del producto de los transgenes sobre otros organismos y el ecosistema parecen ser los principales obstáculos, por los cuales aún esta tecnología no ha logrado una aceptación pública adecuada y una aprobación regulatoria a nivel mundial (12).
En Cuba, desde hace algunos años el Centro de Ingeniería Genética y Biotecnología (CIGB) trabaja en la obtención de plantas transgénicas con resistencia a diferentes plagas y enfermedades que afectan a las plantas, así como el uso de plantas como biofábricas de enzimas de uso industrial, productos biofarmacéuticos u otros obteniendo resultados novedosos en este campo. En el caso específico del Centro Nacional de Sanidad Agropecuaria (CENSA), se trabajó en la obtención específica de vectores que contienen construcciones genéticas para expresar el gene c1 que codifica para la proteína de la replicación del TYLCV y su expresión transitoria en protoplastos obtenidos a partir de plantas de Nicotiana bentahmiana (35). Este artículo reseña tiene como objetivo dar una breve panorámica sobre las diferentes estrategias desarrolladas para la obtención de plantas transgénicas resistentes a geminivirus a nivel mundial, el impacto de esta tecnología en la agricultura actual y los resultados específicos obtenidos en el CENSA.
Estrategias desarrolladas para el control de enfermedades causadas por geminivirus mediante técnicas de ingeniería genética.
Sanford y Johnston (39), propusieron el concepto de resistencia derivada del patógeno como vía alternativa a la creación de genes de resistencia. Ellos sugirieron que los genes de los patógenos, cuando eran expresados por un organismo "hospedante potencial" podrían conllevar a que el mismo se hiciera resistente al patógeno. Teniendo en cuenta esta teoría, en la actualidad las técnicas de ingeniería genética que involucran el uso de genes derivados del patógeno para el control de virus están siendo de amplia aplicación (27,32).
Diversas estrategias se han diseñado y obtenido con resultados altamente satisfactorios. Entre las principales a destacar se encuentran: la resistencia conferida por la sobreexpresión en plantas de genes virales (CP) (4), la cual muestra que la sobreexpresión de un gen desencadena en estas plantas un mecanismo de competencia por la maquinaria celular y las hace mucho más resistentes a la infección contra el virus del cual se obtuvo la secuencia de esta proteína. Otra estrategia se basa en la resistencia mediada por el uso de moléculas defectivas (41) y establece que los ácidos nucleicos defectivos interferentes (DI) son mutantes de deleción de las secuencias genómicas virales, las cuales dependen de sus parentales para la replicación y que al igual que los ARN satélites, son capaces de aminorar los síntomas producidos e interferir en la replicación del virus.
Una de las estrategias más trabajadas en la actualidad lo constituye el uso de fragmentos del ARN viral, en el caso específico de los geminivirus se trabaja con la resistencia mediada por la replicasa viral, o sea, uso del gen c1 que codifica para la proteína Rep con el propósito de interferir con la replicación viral y así conferir resistencia. La interferencia con la acción de la proteína Rep representa una de las opciones más promisorias para la resistencia derivada del patógeno contra las enfermedades causadas por geminivirus. Dos vías son empleadas para su uso: la primera está relacionada con la resistencia mediada por la expresión del gen c1 ubicado en antisentido en una construcción genética (6) y la segunda involucra el silenciamiento génico post-transcripcional (de las siglas en inglés PTGS) utilizando pequeños fragmentos de ARN interferentes (ARNsi) que se basan en el silenciamiento génico dependiente de homología (29,31,32).
- Resistencia conferida por la sobreexpresión de genes virales (proteína de la cápsida).
La resistencia mediada por la proteína de la cápsida es uno de los primeros métodos que se aplicó al control de muchos virus de plantas (45) y es la fuente de resistencia más frecuente derivada del patógeno. Las plantas transgénicas que expresan el gen de la cubierta proteica viral, son mucho más resistentes a la infección contra el virus del cual se obtuvo la secuencia de la proteína de la cápsida (PC). El primer reporte de este tipo de resistencia se obtuvo en 1986, mediante la introducción del gen de la PC del Virus del Mosaico del Tabaco (TMV) en el genoma de las plantas de tabaco (1). Siguiendo este primer ejemplo, la resistencia mediada por la proteína de la cápsida ha sido efectiva al conferir resistencia contra al menos 30 virus de plantas. Sin embargo, el mecanismo responsable de esta resistencia en el hospedante aun no está bien entendido en todos los casos.
Se han postulado dos teorías que explican la resistencia mediada por la proteína de la cápsida: la primera plantea que la PC impide el desnudamiento del ARN viral tan pronto como los capsómeros del virus son removidos, de forma que el desensamblaje del virus no ocurrirá o estará retardado, si los niveles de la PC están por encima de los máximos normales (38). La otra teoría señala que la PC se une a un receptor celular, que permite la entrada y/o el desensamblaje del virus. Al unirse al receptor queda bloqueado el sitio de entrada del virus a la célula y eliminada también la posibilidad de que se formen complejos para el desensamblaje.
En el caso específico de los geminivirus un ejemplo de esta estrategia lo muestra la sobreexpresión en plantas de tomate del gen v1 que codifica para la PC de TYLCV. El mismo se situó bajo el control transcripcional del promotor 35S del Virus del Mosaico de la Coliflor (CaMV) y se clonó en vectores Ti de Agrobacterium, luego se transformaron plantas de tomates sensibles a la infección por el TYLCV. Las plantas transgénicas expresando el gen v1 se inocularon con TYLCV utilizando moscas blancas alimentadas sobre plantas infectadas con el virus, estas respondieron con retardo en la aparición de los síntomas y se recuperaron de la enfermedad con un incremento de la resistencia en inoculaciones repetidas. Todas las plantas transformadas que se recuperaron expresaron la PC del TYLCV (19).
Trabajos realizados que involucran el uso de este método aplicado al control de otros virus de plantas sugieren la posibilidad de ocurrencia de dos posibles riesgos que parecen estar asociados a esta alternativa: la encapsidación de genes heterólogos y la recombinación. Esto fue demostrado al infectar con el Virus de las Venas Necróticas Amarillas de la Remolacha (BNYVV), plantas transgénicas que expresaban el gen de la CP del Virus de la Sharka o Plum Pox Virus (PPV) (21). Sin embargo, en el evento de encapsidación heteróloga el cambio en la progenie del virus es limitado a una generación porque la información genética del virus permanece intacta. En contraste, la recombinación entre los genomas virales y los transgenes conteniendo proteínas de cápsida resultaron en cambios permanentes en la constitución genética de la progenie viral (14).
- Resistencia mediada por ácidos nucleicos defectivos interferentes (DI).
Los ácidos nucleicos interferentes defectivos (DI), son mutantes de deleción de secuencias genómicas virales, que dependen de sus virus parentales para su replicación. Al igual que los ARN satélites, son capaces de aminorar los síntomas producidos por el virus al cual se encuentra asociado y de interferir en su replicación.
Frishchmuth y Stanley (15) descubrieron un brillante acercamiento a la resistencia de geminivirus, al introducir un clon dimérico que contiene el genoma B (ADN-B) del Virus del Mosaico Africano de la Yuca (ACMV) en el genoma de plantas de N. benthamiana utilizando técnicas de agroinoculación. En este experimento las plantas transgénicas desarrollaron síntomas atenuados al compararlos con las plantas no transformadas utilizadas como controles. La transmisión sucesiva de savia entre plantas transgénicas evidenció una disminución en los niveles de ADN viral y acrecentó la atenuación de los síntomas y el número de plantas asintomáticas con muy bajo contenido de ADN viral, lo cual demostró que el efecto del ADN DI en la proliferación del virus es acumulativo.
La infección por el Virus del Encrespamiento de la Remolacha (de las siglas en ingles BCTV) está asociada con la síntesis de novo de una población heterogénea de ADNs viral subgenómico. Otro ejemplo del uso de estas partículas interferentes se demostró al agroinocular con BCTV plantas de N. benthamiana transformadas con una construcción que contenía una repetición parcial de un ADN subgenómico y se observó una disminución de los síntomas típicos a la infección (13). Esta disminución en los síntomas está asociada con la movilización del ADN subgenómico de la copia integrada. En un intento por mejorar la resistencia, plantas de N. benthamiana se transformaron con una construcción que contenía un dímero parcial de un ADN subgenómico mucho más pequeño. Estas plantas no mostraron resistencia a pesar de que el ADN subgenómico se movilizó del genoma del hospedante. Para un mejor entendimiento de las bases del fenómeno de interferencia, ellos compararon la habilidad del BCTV de replicarse y acumularse en discos de hoja de plantas transgénicas resistentes y no resistentes. En estas, ambos ADNs subgenómicos interfirieron con la replicación del virus pero solo en las resistentes el ADN de interferencia suprimió eficientemente la acumulación viral (13).
- Resistencia mediada por el ARN viral.
Inicialmente se pensó que las plantas transgénicas que expresaban las proteínas virales podrían conferir protección contra el virus. En algunos estudios se ha demostrado que la expresión de las proteínas ha sido la responsable de la resistencia al virus, pero en diversos casos también se ha demostrado que esta resistencia ocurre a nivel del ARN (43).
Un método de regulación de genes en el organismo se realiza a través del uso de moléculas de ARN complementarias las cuales son capaces de unirse al ARN transcripto de genes específicos y de este modo previenen su traducción. Los investigadores del tema han desarrollado dos teorías que argumentan y demuestran la resistencia obtenida en experimentos in vitro con vistas a obtener plantas altamente resistentes. La primera relaciona la resistencia mediada por la expresión del gen c1 de los geminivirus en dirección antisentido en una construcción genética (6) y la segunda está asociada con el PTGS mediado por un pequeño fragmento de ARN y se basa en el silenciamiento génico dependiente de homología (29,31,32).
ARN antisentido
Secuencias antisentido apropiadas incorporadas en el genoma de la planta han mostrado que estas bloquean la actividad de genes específicos, por lo que constituyen otra variante para la obtención de resistencia; a este ARN se le ha denominado ARN antisentido o ARN mic (ARN complementario interfiriendo con el ARN mensajero). En este caso se piensa que la resistencia conferida por el ARN antisentido está dada por una interacción entre el ARN viral y el ARN antisentido, el complejo que se forma puede ser rápidamente degradado y el ARN viral estar incapacitado para la traducción (5).
Utilizando este basamento se han obtenido plantas transgénicas de tabaco que contienen una construcción genética que incluye una secuencia de ADN antisentido del gen c1 del geminivirus del Mosaico Dorado del Tomate (TGMV); donde c1 codifica para una proteína absolutamente requerida para la replicación del ADN de TGMV. Cuando estas plantas se infectaron con el virus mediante agroinoculación, la frecuencia de desarrollo de síntomas se redujo significativamente en un número de líneas antisentido, lo cual estuvo correlacionado con la abundancia de ARN antisentido transcripto y con una reducción en el ADN viral en los tejidos de hojas infectadas (9).
Bendahmane y Gronenborn (6), informaron la obtención de plantas transgénicas resistentes a un aislado de TYLCV empleando casetes de expresión que contenían ARNs antisentido. El blanco del ARN antisentido es una porción del ARN mensajero de la proteína Rep. Las plantas transgénicas de N. benthamiana obtenidas expresando el ARN antisentido de c1 mostraron resistencia a la infección por el TYLCV. Algunas de las líneas resistentes fueron asintomáticas y la replicación de TYLCV casi completamente suprimida. La resistencia mediada por transgenes demostró ser efectiva en al menos dos generaciones de la progenie. Así mismo se ha informado recientemente por otros autores el uso de una construcción antisentido conteniendo el gen rep del Virus del Encrespamiento de las Hojas del Tomate (ToLC) (30,33).
A pesar de los altos niveles de resistencia obtenidos utilizando esta variante, dos problemas principales o desventajas son atribuidos al mismo, el primero relacionado con un sustancial exceso de ARN antisentido que requiere frecuentemente una efectiva reducción de los niveles de ARNm blanco y el otro problema es que este método solo es factible de aplicarlo a virus que se replican en el núcleo de las células y no para virus que se encuentran en el citoplasma ya que las moléculas antisentidos transcriptas son producidos en el núcleo. En el caso de los geminivirus donde el ADN viral se encuentra abundante en el núcleo esta estrategia es de mucho valor (17).
Silenciamiento génico postranscripcional (PTGS)
El PTGS es un proceso altamente específico en el cual el ARN con homología al ARN transgénico inductor es específicamente degradado en el citoplasma. Diversos mecanismos se han propuesto para explicar como este sistema de degradación del ARN secuencia-específica podría ser activado, esto incluye el aumento de los niveles del ARNm del transgen por encima del "umbral" requerido, lo que trae como consecuencia la inducción del sistema de degradación, la metilación de los transgenes, lo que resulta en una producción de ARNs aberrantes prematuros para iniciar el mecanismo de degradación y la formación de los ARNs de doble cadena que son capaces de inducir una degradación directa de lo ARNs. Un modelo de cuatro etapas se ha propuesto para esclarecer la vía por la cual ocurre el silenciamiento del ARN (43).
Evidencias genéticas recientes muestran la posibilidad de que la ruta del silenciamiento del ARN está ramificada y que las ramas que se forman convergen en la producción de los ARNs de doble cadena (ARNdc). Estos ARNdc pueden ser producidos por las ARN polimerasas dependientes de ARN codificadas por el hospedante o bien codificadas por el virus, por la transcripción de promotores convergentes, o a partir de loci reorganizados. Las largas moléculas de ARNdc son procesadas en pequeñas moléculas de 21-25 nucleótidos, denominadas ARN interferentes (de las siglas en inglés siRNAs), por una enzima endonucleasa ARNdc- específica similar a la ARNasa III y denominada DICER (43).
Los ARNs interferentes del ARN molde, con ambas cadenas sentido y antisentido son incorporados en una endonucleasa multi-subunidad, denominado complejo del silenciamiento inducido del ARN (de las siglas en ingles RISC) o complejo de interferencia específica del ARN, que es guiado a este ARN molde por apareamiento de bases y por consiguiente ocurre el corte del ARN de secuencias similares, lo que trae consigo la producción del ARNdc (3,18,44). Los ARNs pequeños de interferencia no solo se han encontrado y asociado con el PTGS en plantas. Un proceso similar denominado ARN de interferencia (de las siglas en ingles RNAi) se ha determinado en Caenorhabditis elegans y utilizado como marcador para el silenciamiento génico. También se han encontrado evidencias de que los virus pueden a la vez ser blancos e inductores del PTGS y pueden conllevar al PTGS, lo que sugiere que el PTGS actúa como un mecanismo de defensa natural de las plantas ante la invasión de ARN virales o secuencias de ADN tales como elementos transponibles o el ADN-T de Agrobacterium (43).
Los resultados obtenidos a partir de la recuperación natural de plantas infectadas con la cepa de Camerún del Virus del Mosaico Africano de la Yuca (ACMV-CM) y la correlación encontrada entre esta respuesta natural y los altos niveles de ARN citoplásmasticos pequeños e interferentes (ARNsi) apoyan el mecanismo del silenciamiento génico post-transcripcional como una estrategia de control efectiva para los geminivirus (8). En los geminivirus, se ha demostrado el silenciamiento de un transgen conteniendo el gen GUS puesto bajo la regulación del promotor del TYLCV por la infección de virus homólogos pero no de heterólogos. Este fenómeno ocurre debido a la hipermetilación de secuencias derivadas del promotor TYLCV.
Una estrategia de control basada en el PTGS fue demostrada cuando células de plantas que fueron transfectadas simultáneamente con ACMV (CM) y con un ARNsi sintético diseñado como molde del gen Ac1 del virus mostró una reducción en los niveles de acumulación viral del ARNm de Ac1 en más del 90% y en el 70% del ADN viral, comparado con los controles utilizados. Consecuentemente la expresión transgénica del ARN de doble cadena del gen Ac1 revela una resistencia durable al Virus del Encrespamiento de la Hoja del Algodón en tabaco (2) y al TYLCV en tomate (46).
Otro ejemplo concreto de este tipo de estrategia fue demostrado recientemente donde se analizó el efecto de los ARNsi sobre la inhibición de la expresión del gen de la proteína de la cápsida del TYLCV, donde se evidenció en las plantas transgénicas de tomate la ausencia de síntomas típicos de la enfermedad, aún luego de siete semanas de la inoculación con el virus, mientras que las no transgénicas se infectaron a las dos semanas postinoculación, demostrándose la factibilidad de esta alternativa para conferir resistencia a este virus (47).
Impacto de la transgénesis en plantas en la agricultura mundial
No es menos cierto que el crecimiento de la población mundial tiende a la duplicación en los próximos 10 años y que además de satisfacer el aumento de la demanda de alimentos como resultado de ese crecimiento, el incremento de la producción de alimentos tendrá que satisfacer el aumento del consumo per cápita debido al mejoramiento de la calidad de vida, calculándose que para el año 2025 habrá que aumentar la producción de alimentos en al menos 50% de la que actualmente se produce. La conclusión inevitable derivada de esto es que para poder satisfacer la demanda, la humanidad tendrá que aumentar el rendimiento de las áreas sembradas. Además este incremento tendrá que ser el resultado del uso de menos tierra, agua y menos plaguicidas, por lo que varios factores bióticos y abióticos de estrés, así como varias condiciones desfavorables de post-cosecha, también darán lugar a pérdidas significativas en las producciones agrícolas. Por lo tanto, se necesitará de variedades de cultivos más estables y con mejores rendimientos, así como mejores formas de manejo con el propósito de aumentar la productividad y la sostenibilidad de los cultivos (40).
A fin de enfrentar estos retos será necesario disponer de nuevos conocimientos derivados del avance científico ininterrumpido, el desarrollo de nuevas tecnologías adecuadas y una amplia difusión de dichos conocimientos y tecnologías, así como la capacidad de introducirlos y utilizarlos racionalmente en todo el mundo. En este contexto es que el uso de nuevas tecnologías que al ser aplicadas junto a métodos tradicionales de mejoramiento y manejo de los cultivos cobra una importancia primordial en la producción sostenible de alimentos para la población existente. La biotecnología de plantas podría contribuir sustancialmente al rescate de las cosechas perdidas por el ataque de plagas y enfermedades, así como en el mejoramiento de la producción y calidad de los productos obtenidos.
La necesidad urgente de métodos sustentables para la producción agrícola del mundo con el objetivo de satisfacer las demandas de una población mundial en constante crecimiento hace de la tecnología del uso de plantas transgénicas o de los también denominados, organismos genéticamente modificados (OGMs) en la agricultura; una opción de valor a tener en cuenta. Las plantas transgénicas con caracteres tan primordiales como la resistencia a las plagas, enfermedades y herbicidas, son sumamente importantes y necesarias cuando no se ha detectado resistencia inherente en las especies locales. Por otra parte, estas poseen entre sus beneficios la mayor flexibilidad para administrar el cultivo, menor dependencia de insecticidas químicos, mayores rendimientos, facilidad de cosecha y mayor proporción de esta para su comercialización. Para el consumidor esto significa menor costo del alimento y más valor nutritivo en estos.
A pesar de los resultados demostrados en el uso de la transgénesis en plantas para conferir resistencia a diversas plagas y enfermedades, así como para la expresión de proteínas de interés terapéutico e industrial; y las ventajas que esta tecnología podría traer para el desarrollo biotecnológico y socieconómico, no ha conseguido tener una aceptación favorable por el público. Los principales problemas están relacionados con preocupaciones intrínsecas sobre las plantas y los genes en sí, así como preocupaciones extrínsecas relacionadas con la salud de los consumidores, el medio ambiente y la socioeconomía (28).
Como toda tecnología nueva, la transgénesis enfrenta importantes desafíos como resultado de la percepción del público en general, de tendencias ambientalistas y económicas, así como la predominancia de las compañías multinacionales en el desarrollo y beneficios de esta tecnología. Estudios realizados han identificado riesgos potenciales inherentes a esta novedosa tecnología, que incluye riesgos para la salud y el medio ambiente. Este último, se refiere especialmente a los riesgos potenciales que pudieran atentar contra la biodiversidad. Aquí se han propuesto como riesgos, la adquisición por parte del cultivo de rasgos de "maleza", el desarrollo de resistencias en las plagas y la dispersión o flujo del transgén. En cuanto a los riesgos potenciales para el ambiente se encuentran el flujo e introgresión de genes. De todos los riesgos potenciales atribuidos a los cultivos transgénicos, el flujo de genes y su impacto en la biodiversidad, continúa siendo estudiado y evaluado, especialmente en regiones o países que son centros de origen o centros de diversidad genética.
El flujo vertical de cultivo a cultivo o a parientes silvestres es relevante. Este flujo involucra la producción de híbridos viables que expresan el transgén y con capacidad de adaptación en hábitats naturales o de cultivo. Hay que tener en cuenta que el flujo de genes es una de las fuerzas del proceso de evolución y domesticación de los cultivos y que no es extraño que algunos cultivos, ej. trigo, canola, han resultado de la recombinación de más de un genoma. Para determinar el potencial del flujo de genes en la agrobiodiversidad, cada cultivo debe ser analizado con respecto a los siguientes factores: biología reproductiva, modificación genética (naturaleza del transgén, número de copias insertadas, expresión), dispersión del polen, flujo de genes de cultivo a cultivo (hibridización-posibles consecuencias), definición y estatus de la planta silvestre o maleza, flujo de genes del cultivo a parientes silvestres (compatibilidad y distribución), hibridación, posibles consecuencias del flujo. Entre los factores que afectan la dispersión del polen y la polinización cruzada se encuentra: tamaño (área) del cultivo fuente y receptor de polen, vectores de polen, factores ambientales (clima, barreras biológicas y físicas), viabilidad del polen, grados de alogamia en el cultivo, grado de sincronización en la floración. En caso de que el flujo de genes haya sido exitoso, tiene lugar entonces la hibridación y la introgresión.
Han sido reportadas tasas de flujo de genes de un 1% entre cultivos y parientes silvestres, pero puede llegar hasta 100% cuando el cultivo y los silvestres se encuentran muy cercanos. En algunos casos, las poblaciones de especies silvestres introgresadas han mostrado mayor diversidad genética que aquellos que se encuentran separados del cultivo. La introgresión por flujo de genes ha resultado ser importante en la evolución y domesticación de una docena de especies estudiadas (11).
A pesar de los riesgos asociados al uso de esta tecnología de avanzada es importante señalar que la biotecnología puede ofrecer un gran potencial para desarrollar una agricultura más compatible con el uso sostenible de los recursos naturales. Esto se puede lograr conectando la conservación y uso de la biodiversidad con el desarrollo sostenible de la agricultura. La biotecnología está contribuyendo a revelar en detalles y precisión la estructura genética de las plantas y de la biodiversidad, y este conocimiento está contribuyendo a aumentar la productividad de áreas intervenidas, a reducir la contaminación ambiental y agregar valor a la agrobiodiversidad.
Construcción de un vector quimérico conteniendo el gen c1 del TYLCV aislado en Cuba y expresión transitoria en protoplastos de N. benthamiana. Experiencia del CENSA.
En los laboratorios de Virología Vegetal del CENSA y el de genética de la Facultad de Ciencias de la Universidad de Málaga se construyeron los plásmidos que contienen los genes virales de interés del TYLCV aislado y caracterizado en Cuba (35), los que fueron utilizados para la construcción del vector que se empleó en la transfección de células de protoplastos de N. benthamiana y podrá ser de utilidad en la generación de plantas transgénicas resistentes a este aislado viral posteriormente.
El vector (pTYCU11) obtenido (Fig. 1) está compuesto por una construcción genética que contiene un dímero del origen de replicación del virus y un transgen integrado por el gen c1 (en la orientación antisentido) del virus TYLCV, bajo el control de las secuencias: promotora del CaMV 35 S, secuencia ocs poly A y secuencia amplificadora de la transcripción del transgen (EN), así como un marcador para la selección, en este caso un gen de resistencia a antibióticos (Kanamicina).

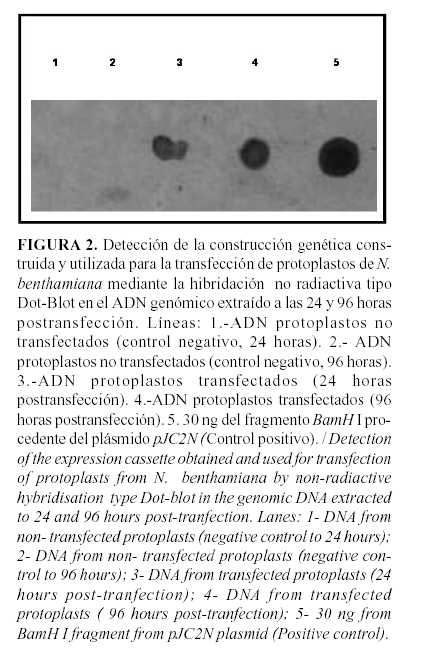
Los protoplastos se aislaron a partir de cinco hojas jóvenes donde se obtuvo un rendimiento de 10 X 106 protoplastos (7,35). Estos fueron tranfectados con la construcción obtenida (pTYCU11) y la detección de la construcción insertada se realizó mediante la hibridación no radiactiva de ácidos nucleicos tipo "dot-blot". Se empleó como sonda un fragmento del plásmido pJC2ENa que contiene el promotor 35S, amplificador y la secuencia señal de terminación de la transcripción del gen ocs. El fragmento se marcó utilizando el juego de reactivos para marcaje no radiactivo empleando la digoxigenina (DIG Nucleic Acid Detection Kit, Boehringer Mannheim), de acuerdo a las instrucciones recomendadas por la casa comercial (35).
La hibridación se realizó utilizando una fracción de los ADN genómicos (125 ng) obtenidos a las 24 y 96 horas post-inoculación a partir de protoplastos transfectados y no transfectados. Se utilizó como control positivo de esta detección el fragmento utilizado como sonda (30 ng) (35). La detección mostró señales positivas en los ADN extraídos a las 24 y 96 horas a partir de los protoplastos transfectados (Fig. 2: Líneas 3 y 4); sin embargo, no se observó señal positiva en los ADN extraídos a partir de protoplastos no transfectados (Fig. 2, Líneas 1 y 2), lo cual evidencia que los protoplastos fueron eficientemente transfectados mediante la metodología utilizada y que la misma puede ser de gran utilidad en los ensayos de expresión transitoria con las construcciones que se obtengan por esta tecnología.
El uso de dos orígenes de replicación viral permite que al producirse la infección y siempre que el virus sea capaz de expresar la proteína del gen c1, sobrepasando la inhibición producida por el ARN antisentido se producirá la formación de partículas extragénicas, a partir de la construcción genómica insertada. Estas partículas contendrán una copia de la construcción genética del ARN antisentido y se pondrían en marcha dos mecanismos adicionales para aumentar la resistencia, uno vinculado a la competencia de las partículas al tener el mismo origen de replicación que el virus por la maquinaria de replicación y el segundo la posibilidad de obtener mayor nivel de expresión de los cassettes en células de la planta hospedante (Bejarano, 2000, comunicación personal).
Teniendo en cuenta lo anteriormente expuesto la construcción genética pTYCU11 incrementará en células transformadas la expresión del gen c1 en la orientación antisentido, a través del aumento del número de copias de la construcción genética por la presencia de las dos secuencias IR, por el control transcripcional bajo el promotor 35 S del CaMV y el amplificador contenido en este promotor el cual aumenta los niveles de expresión, elementos que se han utilizado ampliamente en la ingeniería genética de plantas.
El montaje de la transfección de protoplastos de N. benthamiana con la construcción pTYCU11, permitió la introducción de esta metodología en nuestro laboratorio para la evaluación de las construcciones genéticas, con vistas a la posterior transformación de plantas y la obtención de plantas transgénicas resistentes al aislado cubano de TYLCV. Estos resultados permitirán continuar los trabajos de transformación de plantas siguiendo la estrategia de producción de ARN antisentido, resultados prometedores se han obtenido con la aplicación de esta estrategia como alternativa al control de enfermedades producidas por geminivirus en cultivos de interés económico (6).
Conclusiones y perspectivas futuras
En la actualidad el desarrollo alcanzado por la biología molecular y la ingeniería genética unido a la práctica de métodos convencionales para el control de estas enfermedades han permitido la obtención de plantas transgénicas con alto potencial de resistencia a enfermedades virales. Diversas estrategias se han diseñado, entre las que se encuentran, la resistencia conferida por la proteína de la cápsida viral, resistencia mediada por moléculas defectivas y la mediada por el uso del ARN viral ya sea por del ARN antisentido y por el denominado PTGS que interfieran con la replicación viral, para conferir resistencia. Esta última destacándose en la actualidad como una de las opciones más promisorias para la resistencia derivada del patógeno contra las enfermedades causadas por geminivirus. De forma destacada, la tecnología de transformación tiene un gran potencial de aplicación en muchas áreas de las ciencias de las plantas, tanto básicas como aplicadas. La introducción de transgenes podría facilitar nuevas alternativas en los campos de la genética, la bioquímica y la fisiología, y estimular progresos en estos.
Los resultados alcanzados al transfectar protoplastos con la construcción pTYCU11 obtenida y la alta expresión de esta quimera en estos indican que la misma se replica de forma eficiente en ellos, lo cual garantiza su posterior uso para la transformación de plantas y la consiguiente obtención de plantas transgénicas de tomate y otros hospedantes alternativos del virus, con alta resistencia a este virus.
Es una realidad que el advenimiento de los OGMs ofrece nuevas opciones en la alimentación y agricultura de los países en desarrollo, pero en la actualidad también se habla de la afectación a la diversidad y ecosistemas naturales que podría conllevar un uso desorganizado, desmedido y de lucro de esta tecnología. Es importante destacar que es necesario tener en cuenta los riesgos medioambientales potenciales y los beneficios de esta alternativa en el momento de tomar decisiones sobre el uso de OGMs. Las interacciones complejas que pueden ocurrir entre los OGMs y factores medio ambientales necesitan de una fuerte capacidad científico-técnica para evaluar y manejar los riesgos que estos organismos pudieran conllevar. Por otra parte, es necesario el establecimiento de un marco regulatorio internacional de bioseguridad legislativa basado en el protocolo de Cartagena sobre la bioseguridad de la convención de Diversidad biológica.
AGRADECIMIENTOS
Los autores de este artículo desean agradecer a la Comisión Interministerial de Ciencia y Tecnología de España, por el financiamiento concedido para la ejecución del proyecto bilateral que permitió la obtención de los resultados presentados.
REFERENCIAS
1. Abel PP, Nelson RS, De B, Hoffmann N, Rogers SG, Fraley RT, et al. Delay of disease development in transgenic plants that express the tobacco mosaic virus coat protein gene. Science. 1986;232(4751):738-743.
2. Asad S, Haris WAA, Bashir A, Zafar Y, Malik KA, Malik NN, et al. Transgenic tobacco expressing geminiviral RNAs are resistant to the serious viral pathogen causing cotton leaf curl disease. Arch Virol. 2003,148(12):2341-2352.
3. Baulcombe DC. RNA silencing. Curr Biol. 2002;12: R82-R84.
4. Baulcombe DC. Mechanisms of pathogen-derived resistance to viruses in transgenic plants. The Plant Cell. 1996;8:1833-1844.
5. Bejarano ER, Lichtenstein, CP. Expresion of TGMV antisense RNA in transgenic tobacco inhibits replication of BCTV but not ACMV geminiviruses. Plant Molecular Biology. 1994;24:241-248.
6. Bendahmane M, Gronenborn B. Engineering resistance against tomato yellow leaf curl virus (TYLCV) using antisense RNA. Plant Mol Biol. 1997;33(2):351-357.
7. Bilang R, Klöti A, Shcrott M, Potrykus I. PEG- mediated direct gene transfer and electroporation. Plant Molecular Biology Manual. 1994;A1:1-16.
8. Chepallan P, Vanitharani R, Fauquet CM. Short interfering RNA accumulation correlates with host recovery in DNA virus-infected host and gene silencing target specific viral sequences. J Virol. 2004;77:6785-6798.
9. Day A, Bejarano E, Buck K, Burrell M, Lichtenstein C. Expression of an antisense viral gene in transgenic tobacco confers resistence to the DNA virus tomato golden mosaic virus. Proc Natl Acad Sci. 1990;88:6721-6725.
10.Dempsey DMA, Silva HG, Klessig DF. Engineering disease and pest resistance in plants. Trends in Microbiology. 1998;6(2):54-60.
11.Eastham K, Sweet J. Genetically modified organisms (GMOs): The significance of gene flow through pollen transfer. European Environment Agency, Environment issue report No. 28, Copenhagen; 2002. 75 p.
12.Frankenhuyzen KV, Beardmore T. Current status and environmental impact of transgenic forest trees. Can J For Res. 2004;34(6):1163-1180.
13.Frischmuth T, Engel M, Jeske H. Beet curly top virus DI DNA-mediated resistance is linked to its size. Molecular Breeding. 1997;3(3):213-217.
14.Frischmuth T, Stanley J. Recombination between viral DNA and the transgenic coat protein gene of African cassava mosaic geminivirus. Journal of General Virology. 1998;79:1265-1271.
15.Frischmuth T, Stanley J. Strategies for the control of geminivirus diseases. Seminars in Virology. 1993;4:329-337.
16.Gómez O, Piñon M, Martínez Y, Quiñones M, Fonseca D, Laterrot H. Breeding for resistance to begomoviruses in tropic- adapted tomato genotypes. Plant Breeding. 2004;123(3): 275-279.
17.Hiatt W, Kramer M, Sheehy R. The application of antisense RNA to technology of plants. Proc Nat Acad Sci. 1988;85:49-62.
18.Hutvagner G, Zamore PD. RNAi : nature abhors a double-strand. Curr Opin Genet Develop. 2002;12:225-232.
19.Kunik T, Salomon R, Zamir D, Navot N, Zeidan M, Michelson I. Transgenic tomato plants expressing the Tomato Yellow Leaf Curl Virus capsid protein are resistant to the virus. Biotechnology. 1994,12:500-504.
20.Lapidot M, Friedmann M. Breeding for resistance to whitefly-transmitted geminiviruses. Ann Appl Biol. 2002;140:109-127.
21.Maiss E, Koenig R, Lesemann DE. Heterologous encapsidation of viruses in transgenic plants and in mixed infections. In: Third International Symposium on the Biosafety Results of Field Tests of Genetically Modified Plants and Microorganisms; 1994. p. 129-139.
22.Martínez YM, Zabalgogeazcoa I, De Blas C, Sánchez F, Peralta EL, Ponz F, et al. Geminivirus associated with diseased tomatoes in Cuba. J Phytopathology. 1996;144:277-279.
23.Martínez Y, De Blas C, Quiñones M, Castellanos C, Peralta EL, Romero J. Havana tomato virus, a new bipartite geminivirus infecting tomatoes in Cuba. Arch Virol. 1998;143:1757-1772.
24.Martínez Y, Fonseca D, Quiñones M, Palenzuela I. Presence of Tomato Yellow Leaf Curl Virus (TYLCV) infecting squash (Curcubita pepo) in Cuba. Plant Dis. 2004;88:572.
25.Martinez Y, Muñiz Y, Quiñones M. A new begomovirus infecting peppers plants in Cuba. Plant Pathology. 2006;55(6):817.
26.Martínez Y, Quiñones M, Fonseca D, Potter J, Maxwell D. First report of Tomato yellow leaf curl virus associated with beans, Phaseolus vulgaris, in Cuba. Plant Dis. 2002;86:814.
27.Mubin M, Mansoor S, Hussain M, Zafar Y. Silencing of the AV2 gene by antisense RNA protects transgenic plants against a bipartite begomovirus. J Virol. 2007;4:10.
28.Philipon P, Tastemain C. Plantes transgéniques: les graines de la discorde. Paris. Elsevier; 1998.
29.Pooggin M, Hohn T. Fighting geminiviruses by RNAi and vice versa. Plant Molecular Biology. 2004;55(2):149-152.
30.Praveen S, Kushwaha CM, Mishra AK, Singh V, Jain RK, Varma A. Engineering tomato for resistance to tomato leaf curl disease using viral rep gene sequences. Plant Cell Tissue and Organ Culture. 2005;83(3):311-318.
31.Praveen S, Dasgupta A, Varma A. Phylogenetic analysis and homologies of the replicase of Tomato Leaf Curl Geminivirus: Implications for obtaining pathogen-derived resistance. Virus Gene. 2004;28(1):197-201.
32.Praveen S, Mishra AK, Antony G. Viral Suppression in Transgenic Plants Expressing Chimeric Transgene from Tomato Leaf Curl Virus and Cucumber Mosaic Virus. Plant Cell Tissue and Organ Culture. 2006;84(1): 49-55.
33.Praveen S, Mishra AK, Dasgupta A. Antisense suppression of replicase gene expression recovers tomato plants from leaf curl infection. Plant Science. 2005;168:1011-1014.
34.Quiñones M, Fonseca D, Martínez Y, Accotto GP. First Report of Tomato Yellow Leaf Curl Virus (TYLCV)-infecting Peppers Plants in Cuba. Plant Dis. 2002,86:73.
35.Quiñones M. Estrategia de control por Ingeniería genética a geminivirus aislado en Cuba. Master´s Thesis. Univ. Internacional de Andalucía, España; 2002.
36.Quiñones M, Martínez Y, Fonseca D, Gómez O. Virus del Encrespamiento Amarillo de la Hoja del Tomate (TYLCV): Aportes a los estudios de variabilidad y diagnóstico para Cuba y la región de América Latina y el Caribe. En: Actas de la 44 Reunión de la Sociedad de Fitopatología Americana-División Caribe, Ciudad de La Habana, Cuba; 2004.
37.Ramos PL, Guerra O, Peral R, Oramas P, Guevara RG, Rivera-Bustamante R. Taino tomato mottle virus, a new bipartite Geminivirus from Cuba. Plant Dis. 1997;81:1095.
38.Register III JC, Beachy RN. Resistance to TMV in transgenic plants results from interference with an early event in infection. Virology. 1988;166:524-532.
39.Sanford JC, Johnston SA. The concept fo parasite-derived resistance-deriving resistance genes from the parasite´s own genoma. J Theor Biol. 1985;113:395-405.
40.Sasson A. Cultivos transgénicos: hechos y desafíos. Elfos Scientiae. La Habana; 2001. p 11-16.
41.Stanley J, Saunders K, Pinner MS, Wong SM. Novel defective interfering DNAs associated with Ageratum Yellow Vein Geminivirus infection of Ageratum conyzoides. Virology. 1997;239:87-96.
42.Vanderschuren H, Stupak M, Fütterer Gruissem JW, Zhang P. Engineering resistance to geminiviruses-review and perspectives. Plant Biotechnology Journal. 2007;5(2):207-220.
43.Vanitharani R, Chepallan P, Fauquet CM. Geminiviruses and RNA silencing. TRENDS in Plant Pathology. 2005;10(3):144-151.
44.Voinnet O. RNA silencing: small RNAs as ubiquitous regulators of gene expression. Curr Opin Plant Biol. 2002;5:444-451.
45.Wilson TMA. Strategies to protect crop plants against viruses: pathogen-derived resistance blossoms. In: Proceedings of the National Academy of Sciences. 1993;90:3134-3141.
46.Yang Y, Sherwood TA, Patte CP, Hiebert E, Polston JE. Use of Tomato Yellow Leaf Curl Virus (TYLCV) Rep Gene sequences to engineer TYLCV Resistance in Tomato. Phytopathology. 2004,94:490-496.
47.Zrachya A, Kumar P, Ramakrishnan U, Levy Y, Loyter A, Arazi T, et al. Production of siRNA targeted against TYLCV coat protein transcripts leads to silencing of its expression and resistance to the virus. Transgenic Research. 2007;16(3):385-398.
(Recibido 2-6-2006; Aceptado 13-4-2007)


{kind=link}