










Meu SciELO
 Serviços customizados
Serviços customizadosServiços Personalizados
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Protección Vegetal
versão On-line ISSN 2224-4697
Rev. Protección Veg. v.22 n.2 La Habana maio-ago. 2007
COMPARACIÓN DEL LÍMITE DE DETECCIÓN DE MÉTODOS DE ANÁLISIS DE ÁCIDOS NUCLEICOS PARA EL VIROIDE DEL ENANISMO DEL LÚPULO (HSVd) EN CÍTRICOS
COMPARISON OF DETECCTION LIMIT OF NUCLEIC ACID ANALYSIS METHODS FOR THE HOP STUNT VIROID (HSVd) IN CITRUS
Marianela Soto*, Lien González**, Esther Lilia Peralta*** y Nuria Duran-Vila****
*Dirección de Protección de Plantas, Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, La Habana, Cuba.Correo electrónico: msoto@censa.edu.cu.**Facultad de Biología, Universidad de La Habana, calle 25 e/ H y J, Plaza de la Revolución, Ciudad de La Habana, Cuba.***Instituto de Investigaciones en Fruticultura Tropical (IIFT), Ave 7ma no. 3005 e/ 30 y 32, Playa, C. Habana.****Instituto Valenciano de Investigaciones Agrarias (IVIA), Carretera de Moncada a Náquera, km 4.5, apartado oficial 46113, Moncada, Valencia, España
RESUMEN
Se determinó y comparó el límite de detección de la electroforesis secuencial en geles de poliacrilamida (sPAGE), hibridación de ácidos nucleicos (NASH) y transcripción reversa-reacción en cadena de la polimerasa (RT-PCR) para el diagnóstico del viroide del enanismo del lúpulo (HSVd) en cítricos, basados en el análisis de ácidos nucleicos de plantas de cidro y pepino como amplificadores biológicos. El límite de detección de la sPAGE fue mayor (5x10-3) en plantas de pepino que en plantas de cidro (5x10-2); sin embargo, este fue 50 veces menos sensible que el de la NASH (10-4). El límite de detección de la RT-PCR en pepino fue el mayor de todos los métodos analizados (10-6). Dicho valor se potenció cuando los productos de RT-PCR fueron detectados por NASH en lugar de electroforesis en geles de agarosa. La RT-PCR fue el más sensible de todos los métodos y su sensibilidad se incrementó cuando se combinó con la NASH. Sin embargo, la presencia de contaminaciones asociada a su alta sensibilidad constituye una problemática en el diagnóstico rutinario. Por su parte, la NASH es un método con una sensibilidad adecuada para las condiciones analizadas, su límite de detección fue superior al de la sPAGE, aunque menor que el de la RT-PCR.
Palabras clave: viroide; HSVd; sPAGE; NASH; RT-PCR; diagnóstico molecular.
ABSTRACT
The detection limit of sequence polyacrilamide gel electrophoresis (sPAGE), nucleic acid hybridization (NASH) and reverse transcription-polymerase chain reaction (RT-PCR) was determined for the diagnosis of the hop stunt viroid (HSVd) in citrus hosts, using citron and cucumber plants as biological amplifiers. The detection limit of sPAGE was higher (5x10-3) when cucumber plants were analyzed than that using citron plants (5x10-2); however, it was 50 times less sensitive than that of NASH (10-4) by comparison. The RT-PCR detection limit for cucumber host was the highest (10-6) of all methods analyzed. This value was higher when the RT-PCR products were detected by NASH and no with agarose gel electrophoresis instead. RT-PCR was the most sensitive method and its sensitivity was increased when it was combined with NASH. However, the presence of contaminations associated with the high sensitivity of RT-PCR is a routine diagnosis problem. On the other hand, NASH is a method with a proper sensitivity under the conditions analyzed since the detection limit was higher than that of sPAGE, although slower than that of RT-PCR.
Key words: Viroid; HSVd; sPAGE: NASH; RT-PCR; molecular diagnosis.
INTRODUCCIÓN
Los cítricos constituyen un eslabón fundamental en la economía agrícola de un grupo importante de países, y su gran maquinaria de producción puede verse afectada por un amplio número de enfermedades infecciosas. Dentro de estas se encuentra la cachexia de los cítricos, la cual es ocasionada por determinada variante de secuencia del viroide del enanismo del lúpulo (HSVd) (16,22). Hasta el presente los viroides son las entidades más simples capaces de inducir enfermedades únicamente en plantas superiores. Están compuestos solamente por una molécula de ácido ribonucleico (ARN) de cadena simple, cerrada covalentemente, con un alto grado de complementariedad interna, y no codifican para proteína alguna (3,4,7,8,24). En cítricos se ha descrito la presencia de seis especies de viroides (11,16) aunque sólo dos son capaces de ocasionar enfermedades bien caracterizadas: el HSVd, causante de la cachexia de los cítricos, y el viroide de la exocortis de los cítricos (CEVd), el cual provoca la enfermedad con igual nombre.
Las variantes del HSVd que se presentan en los cítricos se clasifican fundamentalmente en dos tipos, según la presencia de una secuencia motivo que involucra a cinco nucleótidos del dominio variable de la molécula del viroide, la cual define el carácter patogénico o no de las mismas (23). Las variantes patogénicas se refieren en ocasiones como CVd-IIb mientras que las no patogénicas como CVd-IIa (16). Debido a que los viroides no producen proteínas, se impone cierta restricción en los métodos que puedan emplearse para su detección, ya que los métodos inmunológicos, tan ampliamente utilizados para la detección de virus en animales y plantas (5,12,13,33), no pueden ser aplicados en el diagnóstico del HSVd. Igualmente las técnicas de microscopía electrónica son inapropiadas, ya que no se pueden detectar sus partículas características. Por estas razones los procedimientos para su detección han estado confinados a pruebas biológicas, electroforesis en geles de poliacrilamida (PAGE, del inglés sequence polyacrilamide gel electrophoresis), reacción en cadena de la polimerasa combinada con la reverso transcripción (RT-PCR, del inglés reverse transcription-polimerase chain reaction) y la hibridación de ácidos nucleicos (NASH, del inglés nucleic acid hibridization) (2,9).
Para el diagnóstico del HSVd en cítricos se ha utilizado como indicador biológico el mandarino Parson´s Special injertado sobre limonero rugoso (15). Los síntomas de cachexia se manifiestan como depósitos de goma en la línea de unión del injerto, y para su expresión es preciso mantener las plantas a 28-32ºC durante un período de 12-18 meses (19). Existe además otra especie cítrica denominada Clemelín 11-20 (Citrus clementina Hort. Ex Tan. x Citrus sinensis L. Osb), en la cual los síntomas específicos de cachexia ocurren con mayor rapidez e intensidad que en el mandarino Parson´s Special (19). Además de las plantas indicadoras cítricas se han utilizado plantas de pepino (Cucumis sativus L.), las cuales son capaces de replicar al HSVd a niveles detectables, y donde las variantes patogénicas son capaces de inducir un reverdecimiento de las hojas (15).
En Cuba se ha informado la presencia del HSVd en cítricos con un alto porcentaje en plantas de diferentes zonas citrícolas (20,21,32) y durante años el Instituto de Investigaciones en Fruticultura Tropical (IIFT) ha desarrollado un importante trabajo en la caracterización biológica de aislados de este viroide, procedentes de cítricos cubanos (20,21); sin embargo, desde el punto de vista molecular, no se tiene información sobre las variantes de secuencias del HSVd presentes en estos aislados. También se ha trabajado en métodos de detección basados en ensayos biológicos (20,21) y su combinación con el análisis de los ácidos nucleicos mediante electroforesis en geles de poliacrilamida e hibridación de ácidos nucleicos (31). Estos métodos involucran necesariamente un paso previo de inoculación en una especie cítrica que actúa como amplificador biológico (31) a partir del cual se analizan los ácidos nucleicos, lo cual impone como limitante el requisito de al menos tres meses de incubación.
En la actualidad, la citricultura en Cuba ejecuta un proceso de sustitución y diversificación de patrones de cítricos como parte de las medidas de manejo y control del complejo virus de la tristeza de los cítricos-Toxoptera citricida K. Los nuevos patrones, sin embargo, resultan en su mayoría susceptibles a las enfermedades provocadas por viroides (18). En este sentido, la búsqueda de procedimientos rápidos y eficaces, que permitan el análisis rutinario de un gran número de muestras para su aplicación en el sistema de producción de material certificado es una de las líneas de trabajo a seguir.
Teniendo en cuenta estos antecedentes, el objetivo del presente trabajo ha sido determinar y comparar el límite de detección de la sPAGE, NASH y RT-PCR como métodos para el diagnóstico del HSVd en cítricos.
MATERIALES Y MÉTODOS
Material vegetal
Se utilizaron extractos de ácidos nucleicos, purificados según Semancik et al. (26), a partir de plantas de cidro (Citrus medica L.) (10 plantas) y pepino (Cucumis sativus L.) (8 plantas) inoculadas previamente con aislados del HSVd obtenidos en cítricos. Los extractos utilizados pertenecían al banco de colección del laboratorio de viroides del Instituto Valenciano de Investigaciones Agrarias (IVIA), España.
Electroforesis secuencial en geles de poliacrilamida (sPAGE)
La sPAGE se realizó según lo descrito por Semancik et al. (26). Básicamente se ejecutó una primera corrida electroforética en condiciones nativas en un gel de acrilamida al 5% (25). Para ello las muestras se prepararon previamente (20 µL de la extracción de ácidos nucleicos, 20 µL de glicerol al 60% y 10 µL de solución colorante (xilene cianol 0.3%, bromofenol azul 0.3%), y una vez aplicadas se sometieron a una corriente constante de 60 mA a 4ºC hasta que el colorante bromofenol azul migrara aproximadamente 8 cm desde el tope del gel (alrededor de 2.5-3 horas). La corrida electroforética se realizó en tampón TAE 1X pH 7.2 (tris 40 mM, acetato de sodio trihidratado 20 mM, ácido etilendiamino-tetra-acético (EDTA) 1 mM). Una vez detenida la corrida electroforética se tiñó el gel con bromuro de etidio (25), y con este colocado directamente sobre el trasluminador, se cortó de forma horizontal la fracción del gel que contiene a ambos colorantes (llamada zona de viroides).
El fragmento del gel cortado se colocó en el tope de un gel de poliacrilamida desnaturalizante, el cual se preparó previamente. Para la preparación de este gel se mezclaron en un frasco 14.4 g de urea, 3 mL de TAE 3X pH 6.5 (tris 120 mM, acetato de sodio trihidratado 60 mM, EDTA 3 mM); 5 mL acrilamida-N,N´-metilenobisacrilamida 30% y 7 mL de agua destilada, los cuales se disolvieron bajo calor medio (en un agitador magnético con temperatura), y luego se mezclaron con 2.5 mL de TEMED 2% y 0.5 mL de persulfato de amonio 10%.
La corrida electroforética del gel desnaturalizante se realizó a corriente constante (en un rango entre 18-20 mA), a temperatura ambiente por aproximadamente 4 horas. El gel se tiñó con nitrato de plata para visualizar las bandas correspondientes al ARN del viroide (10).
Hibridación de ácidos nucleicos (NASH)
Los extractos de ácidos nucleicos se aplicaron sobre membranas de nylon cargadas positivamente (Boehringer Mannheim), utilizando un equipo en formato de slot-blot conectado al vacío (Schleider & Schuell). Previo a la aplicación en la membrana se desnaturalizaron 20 µL de estos en una solución que contenía 12 µL de SSPE 20X (cloruro de sodio, NaCl 2.9 M; di-hidrógeno fosfato de sodio mono-hidratado, NaH2PO4H2O 0.1 M; EDTA 19.8 mM) y 8 µL de formamida, se mantuvieron a 60ºC durante 15 min. y posteriormente se incubaron en hielo. Las membranas se expusieron a 80°C durante dos horas para fijar los ácidos nucleicos.
La sonda de ácidos nucleicos utilizada consistió en un fragmento de ADN de doble cadena, correspondiente a un monómero del HSVd, al cual se le incorporó la digoxigenina (DIG) como marcador no radiactivo. El método de marcaje empleado consistió en una reacción de PCR según describe Palacio-Bielsa et al. (16), en el cual se utilizó como molde la secuencia completa de una variante del HSVd clonada en un vector plasmídico (30). La diferencia en el contenido de la mezcla de la reacción consistió en que en este caso se añadieron 5mM de dUTP-DIG (Roche). La concentración de ADN se determinó mediante la medida de la densidad óptica a 260 nm de longitud de onda (Ultrospec 2100 pro, Amersham Pharmacia Biotech) (25).
La NASH se realizó según lo describe Palacio-Bielsa et al. (16). Para la prehibridación se añadieron, por cada 100 cm2 de membrana, 10 mL de una solución que contenía 50% formamida; 0.5% sal sódica de dodecilo sulfato (SDS); 6X SSPE (NaCl 0.87 M; NaH2PO4H2O 0.03 M; EDTA 5.94 mM pH 7.4), solución Denhardt 5X (0.1% Ficoll; 0.81% polivinilpirrolidona; 0.1% albúmina de suero bovino) y 10 mg de ADN de esperma de salmón desnaturalizado durante 10 min. a 100ºC. Las membranas se mantuvieron en esta solución a 50°C durante dos horas.
La hibridación se realizó añadiendo, por cada 100 cm2 de membrana, 10 mL de una solución que contenía 50% formamida; 5% SDS; 6X SSPE (0.87 M NaCl; 0.03 M NaH2PO4H2O; 5.94 mM EDTA pH 7.4), 10 mg de ADN de esperma de salmón y 10 mg de sonda de ADN marcada con DIG. Tanto el ADN de esperma de salmón como la sonda se desnaturalizaron por calor previo al ser aplicados a la solución de hibridación. Las membranas se mantuvieron hibridando durante 16 horas a 50°C.
Al concluir el tiempo de hibridación se realizaron dos lavados a temperatura ambiente durante 15 min. en SSC 2X (NaCl 0.29 M; citrato de sodio 0.03 M, pH 7) y 0.1% SDS, y un lavado a 60°C por una hora con SSC 0.1X (NaCl 0.014 M; citrato de sodio 1.5 mM, pH 7) y 0.1% SDS.
La detección de los híbridos marcados se realizó de manera quimioluminiscente, con el empleo de un conjugado anti-DIG-fosfatasa alcalina (fragmentos Fab), siguiendo el procedimiento descrito por el productor (Boehringer Manneheim). Como sustrato de la enzima se utilizó el compuesto quimioluminiscente CSPD (3-(4-metoxispiro{1,2-dioxetano-3,2´-(5´-cloro) triciclo[3.3.2.13,7] decan}-4-il) fenil fosfato (Roche). La visualización de los híbridos se realizó mediante la exposición de láminas X-Omat (Kodak) a 37°C durante 1 hora.
Reverso transcripción y reacción en cadena de la ADN polimerasa (RT-PCR)
La síntesis de la cadena complementaria se realizó mediante el uso del cebador específico del HSVd Vp19 (5´-GCCCCGGGGCTCCTTTCTCAGGTAAG-3´), complementario a las bases 60 a 85 (15). Como paso previo se desnaturalizaron 2.5 mL del extracto de ácidos nucleicos en presencia de este cebador a 85ºC durante 3 min., luego la temperatura se redujo lentamente a 45ºC, a razón de 2ºC por min. Posteriormente se añadió el resto de los componentes de la mezcla (1 mM de dNTPs y 2 U de la enzima transcriptasa inversa del virus de la leucemia murina (M-MLV, Promega) y se mantuvo a 42ºC durante 45 min., seguido de un calentamiento a 90ºC por 3 min. para la inactivación de la enzima (16).
La síntesis de la cadena homóloga y la posterior amplificación del ADN se realizaron con los cebadores Vp19 y Vp 20 (5´-CGCCCGGGGCAACTCTTCTCAG AATCC-3´), este último contiene las bases 78 a 102 de la secuencia del HSVd (15). La reacción de la PCR se realizó en un volumen de 50 mL adicionando 4 mL del producto de la RT a la mezcla de reacción, la cual contenía: MgCl2 1.5 mM, dNTPs 120 mM, 0.5 mM de cada uno de los cebadores y 1 U de Taq ADN polimerasa (Promega). Las condiciones de PCR consistieron en un paso de desnaturalización inicial de 1 min. a 94ºC, seguido de 30 ciclos (desnaturalización a 94ºC, 40s; anillamiento a 60ºC, 40s y extensión a 72ºC, 1 min.), y un paso de extensión final a 72ºC, 5 min.. La síntesis del ADN del tamaño esperado (300pb) se confirmó mediante electroforesis en geles de agarosa al 2% en TBE 0.5X (tris hidroximetil aminometano (Tris-HCl) 0.04M; EDTA 1mM; ácido bórico 0.04M; pH 8), a 100V de voltaje constante. El ADN se visualizó mediante tinción con bromuro de etidio (25).
Los productos de la RT-PCR se visualizaron además mediante NASH. Para ello se desnaturalizaron por calor (100ºC durante 10 min.) 5 µL del producto, se incubaron 5 min. en hielo y se aplicaron sobre membranas de nylon. El procedimiento de la NASH se realizó según se describió anteriormente.
Determinación del límite de detección
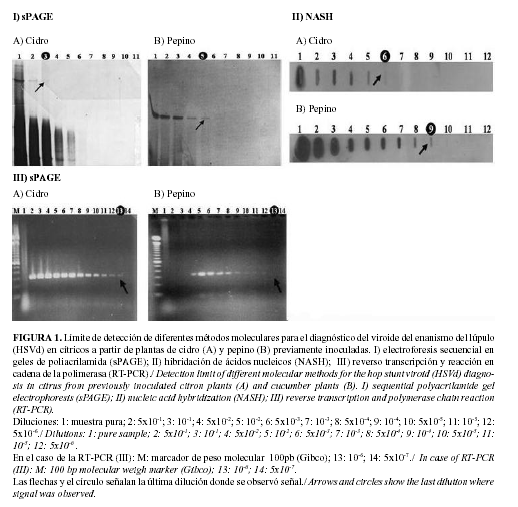
Para los tres métodos de detección del HSVd analizados (sPAGE, NASH y RT-PCR) se determinó el límite de detección a partir de plantas de cidro y pepino previamente inoculadas. Para ello se realizaron diluciones seriadas de los extractos de ácidos nucleicos: muestra pura; 5x10-1; 10-1; 5x10-2; 10-2; 5x10-3; 10-3; 5x10-4; 10-4; 5x10-5; 10-5; 5x10-6; 10-6; 5x10-7. El límite de detección fue hallado como la mayor dilución en la que se evidenció la presencia del viroide, y este fue el criterio para la comparación de dichos métodos.
RESULTADOS Y DISCUSIÓN
Los métodos de análisis de ácidos nucleicos analizados permitieron identificar la presencia del HSVd en el 100% de las plantas de cidro y pepino previamente inoculadas. En los casos de la sPAGE y la NASH se observó que el límite de detección de estas técnicas fue superior cuando el material vegetal de partida se correspondió a plantas de pepino (Fig. 1 I y II). Para estas dos técnicas el uso de la planta herbácea en lugar de la cítrica como amplificador biológico para la posterior detección del HSVd, implicó un aumento de la sensibilidad de las mismas, resultando 10 veces superior en el caso de la sPAGE y 50 para la NASH.
Cuando el análisis se realizó por RT-PCR no se observaron diferencias en el límite de detección a partir de los tipos de materiales vegetales (Fig. 1 III). Cuando se emplearon diluciones bajas del extracto inicial no se observaron señales de hibridación, lo cual puede deberse a la presencia de agentes inhibidores de la reacción de RT-PCR en el extracto de ácidos nucleicos. Este es un fenómeno ampliamente referido por otros autores como uno de los factores determinantes a tener en cuenta durante el desarrollo de esta técnica (14).
Al comparar el límite de detección de cada uno de los métodos se observó que la NASH resultó 50 y 100 veces más sensible que la sPAGE cuando el material vegetal utilizado fue cidro y pepino respectivamente. Mientras, la RT-PCR se comportó con un límite de detección superior a la NASH de 5000 y 100 veces cuando se utilizó cidro y pepino, respectivamente. Cuando los productos de la RT-PCR se visualizaron por NASH en lugar de electroforesis en geles de agarosa, el límite de detección fue de 5x10-7, lo cual resultó superior.
En todos los casos el límite de detección para el HSVd resultó superior cuando se utilizaron plantas de pepino en comparación a las plantas de cidro. Este resultado es consistente con el hecho de que este viroide se replica más fácilmente en la planta herbácea, y por tanto, el ARN se acumula a niveles más elevados (15).
Según lo mostrado anteriormente, de los métodos analizados, la RT-PCR fue el de mayor sensibilidad, la cual se incrementó cuando se combinó con la NASH. No obstante, la problemática de la presencia de contaminaciones, asociada a la alta sensibilidad de la RT-PCR, constituye una limitante para su empleo en el diagnóstico rutinario (1, 17). Otro de los problemas que se presentan durante la realización de la RT-PCR es el efecto inhibitorio de la reacción que producen algunos de los elementos, propios de la planta, remanentes en el extracto de ácidos nucleico. En este sentido, se han realizado un grupo importantes de trabajos con el objetivo de disminuir tal efecto (6, 27, 28, 29).
Por su parte, la NASH es un método con una sensibilidad adecuada, su límite de detección es superior al de sPAGE y es altamente factible para el análisis de un número amplio de muestras (15, 31). Por lo tanto, el uso de la NASH para el diagnóstico rutinario del HSVd en cítricos puede constituir una herramienta importante en el programa de manejo y control de este viroide. El empleo de plantas de pepino como amplificador biológico intermedio potencia la sensibilidad y la utilidad de este método.
REFERENCIAS
1. Bernad L, Duran-Vila N. A novel RT-PCR approach for detection and characterization of citrus viroids. Mol Cell Probes. 2006;20(2):105-113.
2. Cohen O, Batuman O, Stanbekova G, Sano T, Mawassi M, Bar-Joseph M. Construction of a multiprobe for the simultaneous detection of viroids infecting citrus trees. Virus Genes. 2006;33(3):287-292.
3. Ding B, Itaya A. Viroid: a useful model for studying the basic principles of infection and RNA biology. Mol Plant Microbe Interact. 2007;20(1):7-20.
4. Ding B, Itaya A, Zhong X. Viroid trafficking: a small RNA makes a big move. Curr Opin Plant Biol. 2005;8(6):606-612.
5. Esfandiari N, Kohi-Habibi M, Mosahebi G. Occurrence of irases infecting pea in Iran. Commun Agric Appl Biol Sci. 2006;71(3):1281-1287.
6. Fekih Hassen I, Roussel S, Kummert J, Fakhfakh H, Marrakchi M, Jijakli MH. Detection and epidemiological characteristics of peach latent mosaic viroid in Tunisia. Commun Agric Appl Biol Sci. 2005;70(3):115-28.
7. Flores R, Delgado S, Gas ME, Carbonel A, Molina D, Gago S, et al. Viroids: the minimal non-coding RNAs with autonomous replication. FEBS Letters. 2004;567:42-48.
8. Flores R, Hernández C, Martínez de Alba AE, Darós JA, Di Serio F. Viroids and viroid host interactions. Ann Rev Phytopathol. 2005;43:117-139.
9. Hernández C, Di Serio F, Ambrós S, Daros JA, Flores R. An element of the tertiary structure of peach latent mosaic viroid RNA revealed by UV irradiation. J Virol. 2006;80(18):9336-9340.
10.Igloi GL. Silver stain for the detection of nanogram amounts of tRNA following two-dimensional electrophoresis. Anal Biochem. 1983;134:184-188.
11.Ito T, Ieki H, Ozaki K. Characterization of a new citrus viroid species tentatively named citrus viroid OS. Arch Virol. 2001:146(5):975-82.
12.Ma JC, Wang LW, Li XJ, Liao YF, Hu XY, Gong ZJ. Relationship between HBV genotypes and anti-viral therapeutic efficacy of interferon-alpha. Hepatobiliary Pancreat Dis Int. 2007;6(2):166-171.
13.Mostafae S, Mosahebi G, Habibi MK. Potato virus Y isolated from pepper fields in Tehran Province. Commun Agric Appl Biol Sci. 2006;71(3):1335-1340.
14.Nakaune R, Nakano M. : Efficient methods for sample processing and cDNA synthesis by RT-PCR for the detection of grapevine viruses and viroids. J Virol Methods. 2006;134(1-2):244-249.
15.Palacio-Bielsa A, Foissac X, Duran-Vila N. Indexing of citrus viroids by imprint hybridization. Eur J Plant Pathol. 1999;105:897-903.
16.Palacio-Bielsa A, Romero-Durban J, Duran-Vila N. Characterization of citrus HSVd isolates. Arch Virol. 2004;149(3):537-552.
17.Peiman M, Xie C. Development and evaluation of a multiplex RT-PCR for detecting main viruses and a viroid of potato. Acta Virol. 2006;50(2):129-133.
18.Peña I. Pérez JM, Batista L, Velázquez K, Alonso M. Principales enfermedades virales y afines de los cítricos. En: Curso taller sobre producción de material de propagación certificado de cítricos. Editorial Fondo fiduciario Pérez Guerrero, Cuba; 2004. p. 109.
19.Pérez R. Optimización del diagnóstico biológico del viroide de la cachexia de los cítricos. Tesis presentada en opción al grado científico de Doctor en Ciencias Agrícolas, Ministerio de la Agricultura, Instituto de Investigaciones en Fruticultura Tropical (IIFT); 2004.
20.Pérez R. Enfermedades ocasionadas por viroides en la citricultura. En: Curso taller sobre producción de material de propagación certificado de cítricos. Editorial Fondo fiduciario Pérez Guerrero, Cuba; 2004. p. 119.
21.Pérez R, González A, Bello L, Marrero A. Clemelín 11-20: a new indicador plant for citrus cachexia. In: Proc. 7th Intern. Citrus Cong; 1992 marzo 8-13; Acireale, Italia;1992. p. 786-787,
22.Reanwarakorn K, Semancik J. Regulation of pathogenicity in hop stunt viroid-related group II citrus viroids. J Gen Virol. 1988;79:3163-3171.
23.Reanwarakorn K, Semancik J. Discrimination of cachexia disease agents among citrus variants of hop stunt viroid. Ann Appl Biol. 1999;135:481-487.
24.Rocheleau L, Pelchat M. The Subviral RNA Database: a toolbox for viroids, the hepatitis delta virus and satellite RNAs research. BMC Microbiol. 2006;6:6-24.
25.Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning: A laboratory manual. 2da Ed. Cold Spring Harbor Laboratory Press, New York. U.S.A; 1989.
26.Semancik JS, Morris TJ, Weathers LG, Rordorf GF, Kears DR. Physical properties of a minimal infectious RNA (viroid) associated with the exocortis disease. Virology. 1975;63:160-167.
27.Singh RP, Dilworth AD, Singh M, Babcock KM. An alkaline solution simplifies nucleic acid preparation for RT-PCR and infectivity assays of viroids from crude sap and spotted membrane. J Virol Methods. 2006;132(1-2):204-211.
28.Singh RP, Dilworth AD, Singh M, McLaren DL. Evaluation of a simple membrane-based nucleic acid preparation protocol for RT-PCR detection of potato viruses from aphid and plant tissues. J Virol Methods. 2004;121:163-170.
29.Sipahioglu HM, Usta M, Ocak M. Use of dried high-phenolic laden host leaves for virus and viroid preservation and detection by PCR methods. J Virol Methods. 2006;137(1):120-124.
30.Soto Marianela, González Lien, Peralta Esther Lilia, Duran-Vila Nuria. Caracterización molecular de variantes del viroide del enanismo del lúpulo (HSVd) en el aislado cubano PM 26-X. Rev Protección Veg. 2004;19(2):127-130.
31.Velázquez K. Caracterización biológica y molecular de viroides de cítricos de Cuba. Perfeccionamiento de su diagnóstico. Tesis presentada en opción al grado científico de Doctor en Ciencias Agrícolas, Ministerio de la Agricultura, Instituto de Investigaciones en Fruticultura Tropical (IIFT); 2004..
32.Velázquez K, Soto M, Pérez R, Pérez JM, Rodríguez D, Duran-Vila N. Biological and molecular characterization of two isolates of citrus viroids recovered from Cuban plantations. In: Proceedings of the Fifteenth IOCV Conference; 2002 Nov 11-19; Paphos, Chipre; 2002. p. 258-263,
33.Xie L, Wu XD, Huang DZ, Chen HL, He LX, Wang J, et al. Clinical application and analysis of hepatitis C virus NS3 antigen detection by ELISA in human serum. Chin Med J. 2007;120(4):294-299.
(Recibido 6-6-2006; Aceptado 3-4-2007)


{kind=link}