










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión impresa ISSN 1010-2752versión On-line ISSN 2224-4697
Rev. Protección Veg. v.22 n.3 La Habana sep.-dic. 2007
Meloidogyne mayaguensis RAMMAH Y HIRSCHMANN, PLAGA EMERGENTE PARA LA AGRICULTURA TROPICAL Y SUBTROPICAL
Meloidogyne mayaguensis RAMMAH Y HIRSCHMANN, EMERGENT PEST FOR TROPICAL AND SUBTROPICAL AGRICULTURE
Mayra G. Rodríguez, Lucila Gómez y Belkis Peteira
Dirección de Protección de Plantas, Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, La Habana, Cuba. Correo electrónico: mrguez@censa.edu.cu
RESUMEN
Meloidogyne mayaguensis Rammah y Hirschmann, constituye una importante plaga en países de la zona tropical y subtropical, donde parasita de manera natural más de 30 especies de diferentes plantas de cultivo como guayaba (Psidium guajava), cafeto (Coffea spp.), hortalizas, ornamentales y arvenses. Su distribución ha ocurrido relativamente rápido debido a factores antropogénicos y la mayor preocupación con esta especie es su capacidad para parasitar y reproducirse en plantas con resistencia a Meloidogyne spp. Esta especie presenta alta variabilidad en la morfología de su patrón perineal, por lo que es recomendable, para su identificación, el uso de elementos complementarios como los que ofrecen los perfiles de esterasas. Esta especie puede hallarse en poblaciones concomitantes junto a Meloidogyne incognita, Meloidogyne arenaria y Meloidogyne javanica, haciendo más complejo su diagnóstico y manejo. Entre las regulaciones empleadas para mitigar su efecto negativo se encuentran, la implementación de medidas de cuarentena interna y el uso de Cadusafos y Abamectina, con resultados variables. De igual modo, grupos de investigación en África y América han evaluado extractos de Crotalaria spp. y agentes de control biológico (Arthrobotrys spp.), entre otras tácticas. M. mayaguensis se considera una de las especies más peligrosas del género, su detección, correcta identificación y contención en las áreas donde aparece constituyen uno de los retos más importantes de las autoridades sanitarias en los países donde ha sido encontrada, para los cuales la información oportuna y capacitación efectiva de los actores involucrados en esta esfera representa un elemento de vital importancia para el enfrentamiento a esta plaga emergente.
Palabras clave: Meloidogyne mayaguensis; hospedantes; distribución; diagnóstico; manejo.
ABSTRACT
Meloidogyne mayaguensis Rammah y Hirschmann, is an important pest in tropical and subtropical countries, because it affects in natural way more than 30 different species like guava (Psidium guajava), coffee (Coffea spp.), vegetables, ornamental plants and weeds. Its distribution has been very fast due to anthropogenic factors, and the highest concern about this species is its capacity to infest and reproduce on resistant plants to Meloidogyne spp. This species has a high variability in its perineal pattern morphology; thus the use of complementary elements like the offered by esterase profile is recommended. This specie has been found in mixed population with Meloidogyne incognita, Meloidogyne arenaria and Meloidogyne javanica, making more complex its diagnosis and management. Among the measurements to diminish its negative effect are the implementation of internal quarantine measures and the use of Cadusafos and Abermectine with variable results. In the same way, different research teams from Africa and America have evaluated extracts from Crotalaria spp., biological control agents (Arthrobotrys spp.), among other tactics. M. mayaguensis has been considered one of the most dangerous species of this genus. Its detection, right identification and contention in the areas in which it is found constitute one of the most important challenges for the sanitary authorities in the countries when the species has been reported. The opportune information and the effective training of the staff involved in the phytosanitary area represent a vital element for facing this emergent pest.
Key words: Meloidogyne mayaguensis; hosts; distribution; diagnostic; management.
INTRODUCCIÓN
Los nematodos del género Meloidogyne Goeldi constituyen importantes plagas en los países tropicales y subtropicales. Este género posee más de 90 especies descritas (41); sin embargo, las pérdidas atribuidas a dichos nematodos se relacionan a menudo, solo con algunas especies, entre las que usualmente se encuentran Meloidogyne incognita (Kofoid y White) Chitwood, Meloidogyne arenaria (Neal) Chitwood y Meloidogyne javanica (Treub) Chitwood, ampliamente distribuidas en el planeta. No obstante, en los últimos años otras especies han sido informadas también asociadas a daños, destacándose algunas capaces de romper la resistencia de diversos cultivos y que están exhibiendo una dispersión relativamente rápida.
La especie Meloidogyne mayaguensis Rammah y Hirschmann, fue señalada por Fargette et al. (27) como un grupo biológico distintivo y altamente virulento. Según estos autores, resultaba necesario determinar la distribución global de esta plaga potencialmente dañina e implementar medidas de cuarentena adecuadas.
En Cuba, a inicios de la década de los 80' del pasado siglo se encontraron poblaciones de nematodos formadores de agallas compuestas por varias especies en la provincia Granma, entre las que se distinguieron ejemplares cuyas características morfológicas y patogenicidad los diferenciaban de las especies del género previamente informadas para el cultivo del cafeto en nuestro país y se les denominó nueva "forma" de Meloidogyne (30), sugiriéndose posteriormente, sobre la base de estudios preliminares, que podría tratarse de poblaciones de la especie Meloidogyne mayaguensis (20,60). Estudios realizados en la década de los 90' e inicios del 2000 confirmaron la presencia en la zona oriental de M. mayaguensis (55,60), constituyendo actualmente una de las especies de nematodos objeto de cuarentena para Cuba, ubicada en la categoría A2.
Hasta el año 2000, el conocimiento a escala mundial que se tenía de M. mayaguensis era limitado, tanto desde el punto de vista morfológico como de su comportamiento en general, debido a que la misma había mostrado hasta entonces una distribución restringida a pocos países de la región tropical y solo había sido informada como plaga en cultivos de importancia económica en África (59), sin embargo, el informe de su presencia en la península de la Florida en el 2002 llamó la atención de la autoridades sanitarias de la región (53). En la actualidad la importancia de la especie es reconocida y numerosos grupos de investigadores han desarrollado diversos estudios relativos a su distribución, hospedantes, diagnóstico y posibles medidas de manejo.
El objetivo de la presente revisión es poner a disposición de los especialistas de la rama agrícola en el país, un análisis crítico de la información disponible acerca de esta especie, considerada ya en ciertas zonas como especie invasora, teniendo como premisa el hecho de que la información actualizada, resulta vital para decisores, especialistas y agricultores, en el proceso de diseño y ejecución de planes de manejo para una plaga en particular.
PARTE ESPECIAL
La especie de nematodo formador de agallas M. mayaguensis es considerada en la actualidad como una de las más importantes del género por su habilidad de reproducirse en plantas que presentan resistencia a otras especies del género. Su dispersión hacia diversas áreas en países como Brasil (72) y Venezuela (Cassasa, comunicación personal1) por factores antropogénicos (distribución de plántulas infestadas) y las pérdidas provocadas en esas zonas, constituyen elementos que avalan el planteamiento de que representa una plaga emergente que debe recibir atención por parte de las autoridades fitosanitarias de los países donde está informada.
Especie: Origen, distribución actual
La especie M. mayaguensis fue descrita por Rammah y Hirschmann (56), a partir del estudio de una población proveniente de Puerto Rico, que tenía como hospedante original a Solanum melongena L., previamente descrita como M. arenaria y que se encontraba depositada en el banco de nematodos de la Universidad de Carolina del Norte (57), lugar donde se establecieron las poblaciones estudiadas a través del Proyecto Internacional de Meloidogyne.
Esta especie ha sido informada en los siguientes continentes y países: África: Burkina Faso, Burundi, Congo, Costa de Marfil, Malawi, Senegal y Sudáfrica (27,71,76). América: Brasil, Costa Rica, Cuba, Estados Unidos de América, Guatemala, Martinica, Puerto Rico; Trinidad y Tobago y Venezuela (10,20,47,53,56,59,60,64,71,74) y en Europa: Francia (5).
Según la información disponible, en Brasil su diseminación ha ocurrido muy rápidamente, a través de diversos estados y ha causado pérdidas millonarias a la fruticultura de ese país, por lo que recibe en la actualidad atención por parte de la Empresa de Pesquisas Agropecuarias, que emiten información acerca de los municipios donde está presente la especie y las vías de diseminación y desarrolla la capacitación y sensibilización de los actores sociales involucrados en el diagnóstico y manejo de esta plaga en diferentes estados. En este país ha sido encontrada en plantaciones agrícolas (10) y áreas forestales (46). Un fenómeno similar se presenta en Senegal, donde la diseminación ha sido rápida, constituyendo un problema agronómico en ese país (21).
Lo contrario ocurrió en Cuba, su distribución se restringió a las provincias orientales (61), debido a las medidas de cuarentena establecidas por el Centro Nacional de Sanidad Vegetal (CNSV, La Habana), sin embargo, Molinari et al. (49), informaron la presencia de M. mayaguensis en Ciudad de La Habana, en Lycopersicon esculentum Mill. que crecía en la denominada Zona Franca, aspecto que amerita el desarrollo de un estudio a nivel nacional para determinar su actual distribución en el país, como elemento primario en el diseño y establecimiento de programas de manejo.
Identificación de M. mayaguensis: Variabilidad en la morfología del patrón perineal y uso de técnicas complementarias
El diagnóstico de Meloidogyne hasta nivel de género resulta relativamente fácil, sin embargo la determinación de especies es compleja debido a variaciones intra-específicas (39). En este género, esta actividad se realiza en dos niveles. El primer diagnóstico se ejecuta en el campo, cuando se extraen plantas y se observa la presencia de agallas en sus raíces, lo que indicará la existencia de nematodos de los géneros Meloidogyne o Nacobbus Thorne y Allen (falso nematodo de agallas), mientras que la identificación de segundo nivel se ejecuta en laboratorios. Esta abarca estudios morfológicos, moleculares y fisiológicos.
La identificación de las especies dentro del género Meloidogyne es basada primariamente en los caracteres morfológicos. En este género, las características de los patrones perineales de las hembras adultas han constituido históricamente los elementos fundamentales en la identificación de sus especies. Para algunas poblaciones este carácter es muy estable y especie-específico (ej. M. javanica), pero en otras la identificación es más complicada por la presencia de aberraciones y formas intermedias (65). Esto último se pone de manifiesto en los patrones perineales de las poblaciones de M. mayaguensis en varias partes del mundo donde ha sido identificada la especie.
Los patrones perineales de M. mayaguensis, frecuentemente exhiben caracteres atípicos, unas veces, con elementos intermedios entre M. incognita, M. arenaria y M. javanica (12,29,59), otras, donde los patrones se semejan mucho a M. incognita (6,52), resultando difícil la discriminación entre ambas especies.
La población cubana estudiada exhibe los patrones redondeados dorso - ventralmente. Arco dorsal bajo con estrías cuticulares generalmente continuas. Área ventral redondeada. Zona de la cola mayormente elipsoidal y despejada (59,64), carácter que aparece como distintivo en la descripción que, de esta especie, aparece en la página de la Sociedad de Nematólogos (67). Presenta, usualmente, líneas laterales cortas o llegando a los extremos del patrón, incluso, en ocasiones aparecen como pequeños campos laterales que no sobrepasan el área del períneo. En la población se presentan dos tipos de patrones, uno donde las estrías cuticulares se sitúan concéntricas a la vulva y otro donde estas se disponen concéntricas a la cola (64). Las características de los patrones coinciden con lo referido en la descripción original de la especie M. mayaguensis (56).
Por otra parte, en el estudio de las poblaciones foráneas de M. mayaguensis, provenientes de Puerto Rico y Costa de Marfil, Rodríguez (59) encontró que los patrones perineales presentaban características semejantes a M. incognita, destacándose que en ambas no fue frecuente la presencia de la zona de la cola despejada y elipsoidal, carácter señalado en la descripción original de Rammah y Hirschmann (56). Al respecto, se ha reconocido que esta especie, posee un patrón perineal con características variables que abarcan a poblaciones semejantes a M. incognita (29) y M. arenaria (59).
En este sentido, Torres et al. (69) destacaron que la configuración del patrón perineal, considerada aisladamente, no permite un diagnóstico conclusivo de esta especie, pues en muchos casos, las poblaciones presentes en Brasil, exhiben patrones semejantes a M. incognita o aparecen como atípicos. Patrones semejantes a los de esta última especie fueron observados también en poblaciones de M. mayaguensis en África (29), Brasil (12,69) y Florida (6).
Las poblaciones estudiadas en la Florida, poseen perineales similares, en parte, a los informados en la descripción original de M. mayaguensis. Brito et al. (6) señalaron que el 70% de los especímenes examinados mostraban patrones redondeados u ovoides dorso-ventralmente; sin embargo que el restante 30% presentaban arco dorsal trapezoidal similares a los que se presentan en M. incognita, por lo que estos patrones fueron indistinguibles de esta última especie. Estos autores plantearon que no descartan la posibilidad de que M. mayaguensis estuviera presente en la Florida antes del 2002, momento en que fue informada, y que su diagnóstico por parte del servicio fitosanitario fuera de M. incognita.
Con el objetivo de pod0er discriminar M. incognita y M. mayaguensis, Perichi et al. (52) estudiaron poblaciones de ambas especies e informaron que la posición del poro excretor en las hembras adultas puede ser empleado con este fin. Así el poro de M. mayaguensis se encuentra generalmente ubicado a nivel del bulbo medio, como se constata en los dibujos de la región anterior de la hembra que ofrecen Rammah y Hirschmann (56), mientras que en M. incognita, el poro excretor está ubicado cercano a la base del estilete. No obstante esos hallazgos, diversos autores señalan que resulta complicado distinguir M. mayaguensis de M. incognita utilizando solo caracteres morfológicos (6,59).
La existencia de estos antecedentes y el hecho de que Rodríguez (59) encontrara en su estudio que, en una población pura de M. mayaguensis, hay caracteres como patrones semejantes también a los de M. arenaria y otros con características atípicas (aberrantes), indican la necesidad de establecer una metodología que permita a los especialistas, identificar poblaciones de M. mayaguensis con bajas probabilidades de error.
El estudio de machos y juveniles contribuirá a la identificación de la especie. En la población cubana de M. mayaguensis, estudiada por Rodríguez (59), se encontró que los machos poseen el estilete robusto con cabezas basales grandes, redondeadas e inclinadas hacia atrás. La cutícula se presenta finamente estriada transversalmente y con cuatro líneas laterales longitudinales (las dos externas areoladas). Casquete cefálico sin anillación. El hemizonidio se encontró situado de 3 a 5 anillos antes del poro excretor. Con relación a los juveniles de segundo estadio (J2), se constató la existencia de estilete delicado y hemizonidio anterior al poro excretor. Al igual que los machos, presentan 4 líneas laterales. El recto aparece dilatado, las colas delicadas que afinan gradualmente y poseen término redondeado, presentando en ocasiones, un lóbulo. Término hialino bien definido y fasmídeas distintivas. La longitud total de los J2 estuvo entre 398.8 y 500µm (x=446.6 µm); mientras que la longitud entre la base del estilete y la apertura de la glándula dorsal esofágica estuvo entre 3.3 a 4.1µm (x=3.9 µm).
Los estudios complementarios desarrollados, utilizando la población cubana, permitieron establecer que el número de cromosomas somáticos de 2n = 44-45 (valor modal 2n = 44), apareciendo estos en forma de diadas y dispersos en la zona del núcleo (59, 64), valores que se encuentran contenidos dentro de los intervalos informados para M. mayaguensis (2n = 44-45) (60).
Por su parte, los perfiles enzimáticos, en especial los relacionados con la actividad de las esterasas han sido utilizados en los últimos años para el diagnóstico de especies de Meloidogyne y en especial de M. mayaguensis. El estudio ejecutado por Rodríguez (59) evidenció la existencia de coincidencias entre la población cubana de Meloidogyne y la población foránea proveniente de Puerto Rico de M. mayaguensis en cuanto a número de bandas de actividad esterasa no específica (formas electroforéticas) y su movilidad relativa.
Al respecto, se conoce que los informes de presencia de M. mayaguensis efectuados desde inicios del 2000 hasta el presente, han sido sobre la base de estudios de perfiles de isoenzimas (11), en especial de las esterasas (6,69,71,74), donde las poblaciones de esta especie exhiben un perfil de alfa esterasas denominado M2.
El uso de la Reacción en Cadena de la Polimerasa (PCR) con los cebadores que actúan en la región del Espacio Inter- Génico (EIG) entre los genes 18S y 5S (3) permite separar las poblaciones de M. mayaguensis de otras especies como M. incognita, M. javanica, M. arenaria, Meloidogyne paranaensis Carneiro, Carneiro, Abrahantes, Castangnoni y Almeida y Meloidogyne exigua Goeldi (59).
También se ha empleado otros métodos basados en el uso del ADN para distinguir M. mayaguensis de otras especies tropicales de este género, como son el Polimorfismo de la Longitud de los Fragmentos de Restricción (RFLP) (28) y la Amplificación al azar del DNA Polimórfico (RAPD) (4,43). Con relación a esta última técnica, patrones RAPD de aislados de Meloidogyne spp., generados con el cebador o iniciador OPA-4 hicieron posible distinguir M. paranaenses, M. incognita, Meloidogyne arabicida López y Salazar, M. javanica; M. arenaria, M. exigua y M. mayaguensis (14).
Recientemente, Blok et al. (5), demostraron que la amplificación de la región del ADN mitocondrial, entre los genes COII e IARN y la región repetitiva de 63pb puede ser utilizada también para diferenciar M. mayaguensis de las otras especies tropicales.
Con el empleo de Hospedantes diferenciales se pudo comprobar que la población cubana de M. mayaguensis (59) mostró un comportamiento similar al de poblaciones de M. incognita raza 2, tal y como informaran Rammah y Hirschmann (56). Esta población cubana fue capaz de atacar al tomate genotipo Guadajira (33), el cual posee el gen Mi de resistencia a Meloidogyne spp., por lo que demostró su alta virulencia y capacidad para romper estas barreras (59). Sin embargo, el estudio de poblaciones de esta especie provenientes de Florida mostraron un comportamiento similar al de M. incognita raza 4, lo que según Brito el al. (6) fue también observado en poblaciones africanas de M. mayaguensis y sugieren la posibilidad de existencia de razas en esta especie, aspecto que debe ser estudiado en Cuba, por las implicaciones que posee en el manejo de las poblaciones.
Presencia de poblaciones concomitantes
Las comunidades poli-específicas de nematodos parásitos de plantas son encontradas usualmente en suelos agrícolas y ecosistemas estables (1) y en el caso de los nematodos formadores de agallas es común la presencia de dos o más especies de Meloidogyne en el mismo campo, sistema radical y agalla (59).
M. mayaguensis ha sido encontrada frecuentemente en poblaciones concomitantes, donde convive con M. incognita, M. arenaria y M. javanica (7,59,64) y se conoce que la presencia de esta especie en poblaciones concomitantes complejiza el manejo de los nematodos formadores de agallas.
Detección
La adopción de medidas adecuadas de manejo de un organismo plaga están basadas, en primer lugar, en una certera detección e identificación de la misma (39). En Cuba, la detección y estimación de poblaciones de nematodos formadores de agallas presentes en un suelo se ejecuta a través de bioensayos con el uso de las llamadas Plantas Indicadoras (59), donde se utilizan especies y variedades de la familia Cucurbitaceae.
bajo nuestras condiciones, la variedad de calabaza que se ha empleado históricamente en los bioensayos ha mostrado un comportamiento variable. Rodríguez et al. (64) y Sánchez y Rodríguez (inédito2) informaron que no constituyó buen hospedante de M. javanica, mostrando similar comportamiento frente a poblaciones de Meloidogyne sp., del cafeto (59,63); sin embargo, Fernández et al. (32), informaron que la variedad RG-1 se comportó como muy susceptible a diversas poblaciones de M. incognita (razas 1,2 y 3), M. arenaria (raza 2) y M. javanica.
Estos hallazgos sugieren que, para el trabajo con poblaciones compuestas por diversas especies, la selección de hospedantes para el bioensayo debe realizarse previamente, con el objetivo de determinar si este posee selectividad para las especies y razas presentes, siendo recomendable el uso de la madama (Impatiens balsamina L.) cuando se trate de poblaciones en que M. mayaguensis esté involucrada en un alto porcentaje, de ahí que, debe ejecutarse el estudio de nuevas especies, para determinar su factibilidad y ser empleadas como plantas indicadoras, que permitan realizar los bioensayos teniendo en cuenta las condiciones locales: especies de nematodo predominantes, disponibilidad material de siembra; entre otros.
Los resultados obtenidos por Rodríguez (59) señalaron a la madama como el hospedante favorable para la realización de los bioensayos en aquellos suelos donde se posee evidencia o sospecha de la presencia de M. mayaguensis. Además de esta característica, otras no menos importantes son el hecho de que esta planta produce abundantes semillas y es bastante rústica, lo que la convierte en una opción práctica para estos fines.
Plantas Hospedantes: Implicaciones en el manejo
El efecto que ejercen los factores físicos del entorno (suelo) en el establecimiento y desarrollo de las poblaciones de nematodos resulta importante, sin embargo, es vital la función del hospedante como componente del ecosistema, ya que representa el alimento y es quien, en última instancia, determina el tipo y cuantía de los organismos que se encuentran en la rizosfera y como interactúan ya sea positiva o negativamente con los nematodos.
Se conoce que, una parte de las especies de nematodos formadores de agallas son polífagas. Según Jepson (39) cada una de las cuatros especies "mayores" (M. incognita, M. arenaria, M. javanica y M. hapla Chitwood) parasita un gran número de hospedantes con representantes en casi todas las familias botánicas.
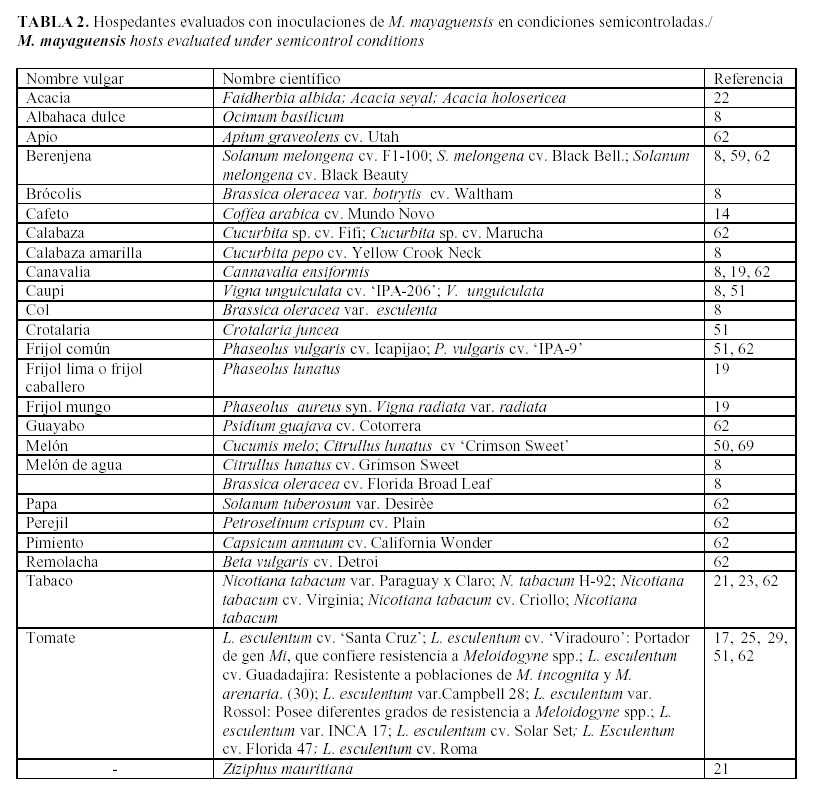
M. mayaguensis, aparece en el panorama mundial como una especie polífaga, que parasita plantas de cultivo, ornamentales y arvenses (Tabla 1), lo que constituye un elemento a tener en cuenta en el diseño e implementación de estrategias de manejo.
La utilización de tácticas como la rotación de cultivos y el uso de variedades o portainjertos resistentes debe ser antecedida por la evaluación de los genotipos a emplear antes de ser llevados a campo.
Por su parte, debido a la importancia de esta especie de Meloidogyne, numerosos grupos de investigadores han evaluado la capacidad hospedante de diversos cultivos en condiciones semicontroladas, resultados que se presentan en la Tabla 2. En ella se puede constatar que especies pertenecientes a familias botánicas como Fabaceae, Cucurbitaceae, Rubiaceae, Brascicaceae, Myrtaceae y Solanaceae son parasitadas.
Al estudiar los resultados obtenidos por los diferentes grupos de investigadores que trabajan con M. mayaguensis se puede constatar la gran variabilidad en la capacidad como hospedantes de diversas plantas para esta especie de Meloidogyne. En este aspecto, poblaciones de Brasil que parasitan guayaba no lo hacen en cafeto (Hidalgo, comunicación personal3).
Debido a que una de las medidas de manejo de nematodos más efectivas, lo constituye la rotación de cultivos, en especial en áreas productoras de hortalizas, papa, ornamentales anuales, frijoles y otros cultivos no perennes, resulta imprescindible evaluar los cultivos frente a la población de nematodos formadores de agallas antes de llevarse al campo, de ahí la necesidad de continuar este tipo de estudio para el caso de la población cubana de M. mayaguensis.
Sintomatología y Daños
Las interacciones de los nematodos parásitos de plantas y sus hospedantes son complejas y dinámicas y pueden abarcar, dependiendo de las especies, la estimulación de la eclosión de los huevos, atracción al hospedante, penetración y migración en los tejidos, el reconocimiento de los tejidos favorables para el establecimiento del sitio de alimentación y la elaboración de células modificadas en el hospedante (37).
El parasitismo de los nematodos en las raíces puede (y tiene) un impacto dramático en la fisiología y crecimiento de las plantas. Los síntomas que provocan son variados y están directamente relacionados con los niveles de nematodos en las raíces (37).
En Cuba, Rodríguez (59), describió la sintomatología provocada en cafeto por poblaciones mixtas de M. mayaguensis (especie en mayor proporción), M. incognita y M. arenaria, señalando que, más del 30% de las plantas afectadas presentaban alto grado de defoliación, muy marcada en las ramas de las zonas baja y media de la copa. Gran parte de las hojas que permanecían en las plantas estaban cloróticas, excepto la parte superior correspondiente a la zona de crecimiento donde estas eran verdes. Las plantas con estos síntomas exhibían exigua producción de cerezas. Las más afectadas podían ser extraídas con relativa facilidad y muchas morían. El cuadro de síntomas aparece generalmente en forma de parches en los campos, tanto en las laderas de elevaciones como en la base de estas.
En las áreas estudiadas, el 57.2% de las plantas evaluadas exhibían agrietamiento en la zona del cuello de la raíz y parte baja del tronco, que alcanzaba entre 20-25cm de altura en el tronco, este puede presentarse como una pequeña herida o en todo el perímetro del tronco en esta zona baja, presentando suberización de la zona afectada dándole una apariencia corchosa. Las raíces secundarias se observaron también agrietadas. En aquellas plantas donde la afectación era severa se producía la pérdida del sistema de raíces secundarias, la raíz se engrosaba y las zonas de las rajaduras se presentaban más oscuras. Cuando estas áreas afectadas se seccionaron longitudinalmente, se constató la presencia de hembras de Meloidogyne, formando hileras a lo largo de las rajaduras.
En numerosas plantas produce una proliferación de raicillas que parten perpendicularmente de la zona de la raíz por debajo del cuello, y gran parte de estas raicillas estaban agalladas y los cecidios, de hasta 5mm de diámetro, aparecían individualmente o formando collares, en muchos casos se presentó agrietamiento del tejido de la superficie en los mismos y en su interior se encontraron hembras en todos los estadios de desarrollo.
Por su parte, en las plantas afectadas por Meloidogyne spp., con predominio de M. mayaguensis, se pudieron observar síntomas radicales similares a los descritos para C. arabica, en las plantas de C. canephora, aunque las rajaduras ocuparon, solo ¼ del perímetro del tronco, de manera general. Se observaron agallas pequeñas en las raíces superficiales que se desarrollaban en los primeros 30 cm de profundidad (59). En dicho estudio, bajo las condiciones en que se desarrolló la evaluación, C. canephora se comportó como tolerante al complejo de Meloidogyne donde M. mayaguensis fue la especie mayoritaria en la población, aunque cuando el nivel de la población aumentaba se producían afectaciones (31,59).
En hortalizas, se observó que en plantas de pimiento parasitadas por una población mixta de M. mayaguensis y M. incognita, en Brasil, presentaron clorosis, disminución del crecimiento, marchitamiento en las horas más calientes del día y una consecuente reducción de la cantidad y calidad de los frutos. Los sistemas radiculares severamente infestados por los nematodos presentaron pobre desarrollo con un gran número de agallas y ausencia de raíces secundarias (18).
En la actualidad, M. mayaguensis constituye una de las especies de nematodos formadores de agallas de importancia para América Latina. Al respecto, Carneiro et al. (13) la incluyen entre las que mayores daños ocasionan al cultivo del cafeto.
Investigadores brasileros, desarrollan desde el año 2003, investigaciones en la Universidad Estadual del Norte Fluminense y han determinado, que en guayaba (P. guajava) aun con la aplicación de materia orgánica como táctica de manejo, las pérdidas han llegado al 30%, teniendo en cuenta que las dos variedades que usan, Paluma y Ogawa, son altamente susceptibles a M. mayaguensis (72), planteándose que podría aniquilar la producción de guayaba (2).
En un estudio ejecutado, comparando la patogenicidad M. mayaguensis con M. incognita, M. arenaria, M. javanica y M. floridensis en tomate, se encontró que M. mayaguensis exhibió la mayor producción de huevos y porcentaje de agallamiento en las raíces, donde el tamaño de las agallas fueron además inusualmente grandes (16), lo que podría estar relacionado con la agresividad de esta especie.
Además de afectar frutales como guayaba, M. mayaguensis fue encontrada en el estado de São Paulo, Brasil, parasitando un portainjerto de pimiento y dos variedades de tomate (resistentes a Meloidogyne), señalando Carneiro et al. (2005), citados por Cirotto et al. (18) que dicho nematodo viene causando pérdidas en estas hortalizas en los municipios de Pirajúi, Santa Cruz do Rio Pardo, Reginópolis y Campos Novos Paulista.
En Guadalupe y en Martinica, M. mayaguensis está sistemáticamente asociado con un grave deterioro de los guayabos, llegando, incluso, a causar la muerte de los árboles jóvenes de 5 a 7 años de haber sido plantados. Esta especie emergente de nematodo por el momento limitada a las zonas tropicales, tiene importantes capacidades parasitarias y representa actualmente una amenaza, ante su eventual introducción y aclimatación en Europa (55).
Prácticas utilizadas y/o estudios de nuevas tácticas para su posible uso en el manejo de las poblaciones de M. mayaguensis.
Medidas de exclusión: En Cuba se ejecutan medidas cuarentenarias para evitar la diseminación de M. mayaguensis desde hace más de 15 años (61), lográndose que la especie tenga en la actualidad una limitada distribución en el país.
En Brasil, investigadores y autoridades sanitarias han determinado la necesidad de establecer medidas de exclusión (70,72) para detener la rápida diseminación del parásito en estados productores de guayaba.
Material de siembra: Se recomienda el uso de plántulas certificadas en el caso de guayabo (72) y cafeto (31), para evitar que el parásito sea introducido a nuevas áreas.
Evaluación de genotipos para su uso en programas de manejo: Han sido desarrollados estudios de capacidad hospedante de diversas especies, líneas y variedades y sus resultados se recogen en la Tabla 2, donde se exponen los hospedantes de M. mayaguensis. Plantas de muy diversas familias botánicas son hospedantes de este nematodo, por lo que se hace difícil el diseño de estrategias de manejo que incluyan la rotación de cultivos.
Sin embargo, se conoce que el maní forrajero (Arachis pintoi L.) no fue capaz de mantener y multiplicar las especies M. incognita y M. mayaguensis, aseverando Queneherve et al. (54) que, esta planta tampoco es hospedante de las principales especies de nematodos agalladores. Según estos autores, el uso de dicha planta tiene como ventajas: la existencia de semillas comercializadas, la propagación por semilla o vegetativa; no es hospedante de varias especies de nematodos incluido Meloidogyne spp., es compatible como planta de cobertura y el aporte de aproximadamente 60 kg. ha-1 al año de nitrógeno al suelo. Por su parte, señalan como inconvenientes: que son plantas hospedantes de nematodos endoparásitos migratorios, como Radopholus similis y Pratylenchus coffeae; de instalación lenta y la necesidad de inocular la bacteria específica asociada.
La introducción y la utilización de A. pintoi como planta de cobertura en plantaciones podría pues efectuarse en sitios en Martinica y las Antillas en otros agrosistemas distintos que aún no se han experimentado, sobre todo en huertas de frutales, como los cítricos y sobre todo, para los guayabos que sufren graves ataques de M. mayaguensis en las Antillas (54).
En Venezuela, Crozzoli et al. (19), evaluaron diversas especies, líneas y variedades de leguminosas y encontraron que P. vulgaris cv. MGM-10-02-35 fue resistente a M. mayaguensis, genotipo que pudiera constituirse en una opción para emplear en suelos infestados con esta especie de nematodo.
El uso de patrones tolerantes está siendo evaluado en el cultivo de guayaba (P. guajava ), uno de los más afectados en el continente, por especialistas de Martinica (44), quienes introdujeron en 1999, desde Costa Rica a Psidium friedrichstalianum (Berg.) Nied. El que no ha mostrado incompatibilidad con las variedades actuales de guayaba. Según estos autores, la técnica para injertar está bajo ensayo para mejorar el porcentaje de éxito y se desarrollan experimentos de campo para determinar la eficacia de esta asociación a largo plazo, el crecimiento del árbol, la productividad y la calidad de la fruta.
Inspecciones a las plantaciones establecidas: Resulta muy conveniente, efectuar muestreos para determinar nuevas áreas infestadas y en el caso de frutales como la guayaba, en Brasil indican a los productores la búsqueda de síntomas como la presencia de hojas con bordes amarillos o rojizos y la presencia de agallas en las raíces (72) como indicativo de la presencia del nematodo, para tomar medidas que van desde la aplicación de materia orgánica y productos químicos, hasta la eliminación de las plantas afectadas.
En Cuba, se ejecuta de forma anual una encuesta para M. mayaguensis (61), y así determinar la presencia del nematodo en nuevas áreas y los niveles poblacionales en las zonas donde está presente, como elemento base para establecer medidas de manejo, sin embargo, resulta necesario perfeccionar el método de detección y diagnóstico para esta especie, toda vez que posee características que hacen posible sea confundida con otras especies de Meloidogyne de amplia distribución en nuestro país.
Tratamiento con productos comerciales en áreas infestadas: Hace una década, en Sudáfrica, en guayaba se comenzó a emplear el nematicida Cadusofos (100g.kg-1 a razón de 10g.m2-1) en la zona de irrigación, siendo duplicado al siguiente año, observándose que al tercer año, los árboles con infestaciones de M. mayaguensis mostraban buenas condiciones (76).
En Brasil, se estudió el efecto de la Abamectina y el Cadusofos en la reducción de las poblaciones de M. mayaguensis en el cultivo de la guayaba, teniendo en cuenta la especial problemática que poseen los productores de esta fruta en dicho país con la dispersión y daños provocados por este nematodo.
Moreira et al. (51) evaluaron la eficiencia de las dosis 2, 4 y 8 mL. planta-1 de Abamectina (diluida en 5 litros de agua, para cada planta) y de Cadusofos, 22.5 g de producto comercial.planta-1(incorporado el suelo), aplicados al ruedo de las plantas, en tres aplicaciones con intervalos de 30 días. A los 60 días desde la primera aplicación, se extrajeron muestras de suelo y raíces, que fueron procesadas por el método de Embudos de Baermann y se contabilizó la población total de juveniles de segundo estadio, para determinar el porcentaje de eficiencia de los tratamientos. En este estudio, todos los tratamientos produjeron reducciones de la población en suelo y raíces, pero las mayores se produjeron con las dosis de Abamectina de 4 y 8 mL. planta-1, (48% en suelo y 66% en raíces y 44% -51% respectivamente). Por su parte, el Cadusofos, produjo reducciones de la población de 71% en suelo y 10% en las raíces.
Se ha informado que investigadores brasileros han evaluado tácticas como la aplicación de abundantes volúmenes de materia orgánica en las plantaciones de guayabo afectadas por M. mayaguensis con resultados favorables y que han obtenido respuesta de resistencia en un material evaluado que debe ser estudiado ahora como portainjerto de P. guajava (72).
Prospección y Evaluación de Agentes de Control Biológico: Dos aislamientos de Arthrobotrys oligospora Fresenius fueron aislados y evaluados en Senegal, como agentes de control biológico, los que atraparon el 87% de los juveniles de segundo estadio de M. mayaguensis en experimentos in vitro. En el estudio ejecutado en macetas, el índice de agallamiento (evaluado según escala de Zeck) fue menor (grado 2) en los tratamientos con los dos aislamientos del hongo, difiriendo significativamente del tratamiento con nematodos sin el agente de control biológico, donde el valor fue de 4, teniendo igual comportamiento el Índice de reproducción del nematodo. Cuando se evaluaron los aislamientos en condiciones de campo, uno de ellos, el denominado ORS 18692 S7 produjo menor índice de agallamiento y una disminución de las poblaciones (25). En este estudio, los autores plantearon que este aislamiento podría contribuir al control de M. mayaguensis dentro de programa MIP, pero señalaron que se necesitaban otros estudios.
A pesar de que se plantea que estos hongos atrapadores no presentan especificidad en su acción sobre estados vermiformes de nematodos, Duponnois et al. (26) informaron que los aislamientos ORS 18690 S2, ORS 18692 S5 y ORS 18692 S7 atraparon juveniles de M. mayaguensis en diferentes proporciones, pero no capturaron juveniles de M. incognita y M. javanica. En su estudio, estos autores hacen referencia a que la acción "atrapadora" de estos hongos está bien estudiada y que tiene relación con la existencia de glicoproteínas compatibles, como las lectinas, en la cutícula de los juveniles, aseverando que la mejor diana de los aislamientos Saharianos de Arthrobotrys fue M. mayaguensis.
No obstante, los resultados satisfactorios de estos ensayos con tomate y tabaco (23), así como el hecho de que los investigadores de ORSTOM- Francia, habían hallado el método de reproducir el hongo en bloques de estiércol y otros desechos (21), está disponible poca información acerca de su uso ulterior en las condiciones de África, contando solo con el informe de que la actividad antagónica de A. oligospora es mayor cuando están presentes hojas de Acacia holoserica (24).
Duponnois et al. (26) sugirieron evaluar inóculo dual de A. oligospora y Pausteria penetrans (Thorne) Sayre y Starr, el cual podría tener un gran potencial para el control de M. mayaguensis en campo, sin embargo, en estudios ejecutados por Trudgill et al. (71), este nematodo se comportó como resistente a la infección por P. penetrans. En la Florida, no se encontraron juveniles de M. mayaguensis infectadas de P. penetrans (7), aspecto que debe continuar siendo objeto de investigaciones, teniendo en cuenta la diversidad de aislamientos que se posee de esta bacteria.
En Brasil se trabajará en la búsqueda de agentes de control biológico para el manejo de M. mayaguensis con la colaboración de especialistas cubanos (2,34).
Modificaciones en la rizosfera: Se conoce que factores relacionados con el ambiente (planta_suelo) poseen influencia en la composición y cantidad de los exudados radiculares y que estos a su vez, ejercen influencia en la eclosión de los huevos de los nematodos. La modificación de los factores que provoquen retardo en la eclosión de los huevos, podrían tener influencia en el manejo de las poblaciones.
En el caso de M. mayaguensis, Siqueira et al. (66) desarrollaron un estudio para determinar el efecto de los lixiviados de caupí (Vigna unguiculata (L.) Walp, cv. Epace 10) inoculado con Glomus etunicatum Becker y Gerdemann y/o Bradyrhizobium sp. sobre la eclosión de los huevos de esta especie y en los de M. incognita y M. javanica. Estos autores encontraron que la eclosión de los huevos en Meloidogyne varía con el tipo de lixiviado y el tiempo de exposición a estos. Para la especie M. mayaguensis, la mayor eclosión ocurre en agua y en los lixiviados de caupí inoculado con una mezcla de estirpes de Bradyrhizobium sp., seguida de los valores obtenidos en los lixiviados de caupì cv. Epace 10 con G. etunicatum + Bradyrhizobium sp.
Estos elementos son importantes si van a utilizar estos organismos en condiciones de campo, resultando necesario en nuestras condiciones, la evaluación del comportamiento de este parámetro en plantas tratadas con EcoMic® u otro producto/organismo con función similar, sabiendo que no constituyen agentes de control biológico y que su efecto sobre las poblaciones de Meloidogyne es variable.
Extractos y/o productos de origen botánico: Según Jourand et al. (40), la habilidad de las especies de Crotalaria de fijar nitrógeno por la simbiosis con organismos como Bradyrhizobium sp. o Methylobacterium sp. , han hecho que se utilicen frecuentemente en los sistemas agrícolas y teniendo en cuenta que algunas leguminosas han mostrado alta resistencia a nematodos formadores de agallas, el uso de Crotalaria sp., aparece como una alternativa muy interesante para reducir las poblaciones en los suelos agrícolas.
Estos autores evaluaron in vitro la producción de parálisis en los segundos estados juveniles de tres importantes especies para la agricultura mundial: M. incognita, M. javanica y M. mayaguensis por los extractos acuosos de raíces y tallos frescos de 15 especies de Crotalaria procedentes de África Occidental. En este estudio se demostró que la acción de las especies de este género varía con la especie de nematodo evaluada. De las especies evaluadas, los extractos de raíces de Crotalaria atrorubens y Crotalaria lathyroides fueron más efectivos sobre M. mayaguensis; mientras que los extractos de tallos de C. grantiana fueron efectivos contra las tres especies de nematodos.
En este estudio se discute la sostenibilidad del uso de extractos y se aconseja el uso de Crotalaria como abono o enmienda verde y/o intercalado en plantaciones, evaluando antes el efecto nematostático de las partes u órganos a emplear, aspecto que debe ser evaluado en Cuba.
Aplicaciones de materiales orgánicos: El efecto reductor de los abonos y otros materiales orgánicos sobre las poblaciones de fitonematodos es bien conocido. En este sentido, Sousa et al. (68) evaluaron cuatro aplicaciones de estiércol vacuno curado (60kg.planta-1 a intervalos de 120 días entre cada aplicación), incorporando dicho material en el área del ruedo de las plantas de guayaba. También el uso de estiércol curado mezclado con 200g de torta de neem (Azadirachta indica A. de Jussieu) (con intervalos de 105 días entre los tratamientos) y un tercer tratamiento con estiércol complementado con follaje de Mentha sp. y Mucuna aterrina Hol. En todos los casos se produjeron reducciones sustanciales de la población de M. mayaguensis en el suelo de guayaba y aumento en los rendimientos.
CONCLUSIONES
M. mayaguensis constituye una importante plaga en cultivos relacionados con la alimentación y con otros generadores de rubros exportables en nuestra región. Las dificultades para su diagnóstico certero y su rápida distribución en algunos territorios sugieren la necesidad de desarrollar estudios conducentes a determinar su distribución y hospedantes naturales, optimizar métodos de diagnóstico y determinar la variabilidad morfológica y patogénica de sus poblaciones, así como la evaluación de variedades y tácticas para el diseño de programas de manejo.
AGRADECIMIENTOS
Los autores desean expresar su agradecimiento a la Dra. Yaima Arocha (CENSA) por su colaboración en el suministro de información para la elaboración de este artículo.
REFERENCIAS
1. Abawi GS, Chen J. Concomitant pathogen and pest interactions. In: Barker K, Pederson G, Windham G, Editors. Plant and Nematode Interactions. Agronomy Monograph No 36. Madison, Wisconsin, USA; 1998. p. 135-158.
2. Anónimo. Cultura da goiaba enfrenta problemas; 2006. (Consultado: 2 Jul 2007). Disponible en: http://www.todafruta.com.br/todafruta/mostra_conteudo.asp?conteudo=11093.
3. Blok Vivian C, Phillips MS, Fargette M. Comparison of sequences from ribosomal DNA Intergenic Region of Meloidogyne mayaguensis and others Major Tropical Root-Knot Nematodes. J Nematol. 1997;29(1):16-22.
4. Blok Vivian C, Phillips MS, McNicol JW, Fargette M. Genetic variation in tropical Meloidogyne spp. as shown by RAPD-PCR. Fundam Appl Nematol. 1997;20:127-133.
5. Blok Vivian C, Wishart Jane, Fargette Mireille, Berthier Karine, Phillips MS. Mitochondrial DNA differences distinguishing Meloidogyne mayaguensis from the major species of tropical root-knot nematodes. Nematology. 2002;4(7):773-781.
6. Brito Janete A, Powers TO, Mullin PG, Inserra RN, Dickson DW. Morphological and Molecular Characterization of Meloidogyne mayaguensis Isolates from Florida. J Nematol. 2004;36(3):232-240.
7. Brito Janete A, Stanley J, Cetintas R, Powers T, Inserra R, McAvoy G, et al. Meloidogyne mayaguensis a new plant nematode species, poses threat for vegetable production in Florida. 1-6; 2004. (Consultado: 3 Jul 2007). Disponible en: http://www.mbao.org.2004. proceeding08.pdf .
8. Brito Janete A, Stanley J, Mendes ML, Cetintas R, Dickson DW. Host status of selected cultivated plants to Meloidogyne mayaguensis in Florida. Nematropica. 2007;37(1):65-71.
9. CAB International. Meloidogyne mayaguensis. In: Crop Protection Compendium, global module, 3rd ed. Wallingford, UK: CAB International; 2001.
10.Carneiro Regina MDG, Alameida Maria RA, Queneherve P. Enzyme phenotypes of Meloidogyne spp. populations. Nematology. 2000;2(6):645-654.
11.Carneiro Regina MDG, Almeida María RA, Carneiro RG. Enzyme phenotypes of brazilian populations of Meloidogyne spp. Fundam Appl Nematol. 1996;19(6):555-560.
12.Carneiro Regina MDG, Moreira WA, Almeida María RA, Gomes ACMM. Primeiro registro de Meloidogyne mayaguensis em goiabeira no Brasil. Nematol Bras. 2001;25:223-228.
13.Carneiro Regina MDG, Barros EVSA, Goncalves W, Pereira AA. Reaction of Coffea arabica genotypes to Meloidogyne spp.; 2006. (Consultado: 2 Jul 2007). Disponible en: http://www.asic-cafe.org/pdf/abstract/PA200_2006.pdf.
14.Carneiro Regina MDG, Tigano Myrian, Randig O, Almeida Maria RA, Sarah JL. Identification and genetic diversity of Meloidogyne spp. (Tylenchida: Meloidogynidae) on coffee from Brazil, Central America and Hawaii. Nematology. 2004;6(2):287-298.
15.Carneiro Regina MDG, Mônaco Amaral, Ana P Do, Peruzzo Marcela, Nakamura Kelly C, Scherer Alexandra. Identificação de Meloidogyne mayaguensis em Goiabeira e em plantas invasoras, em solo argiloso, no Estado do Paraná. Nematol. Bras. 2006;30(3):293-298.
16.Cetintas R, Dickson D, Brito Janete A, Nyczepir A. Pathogenicity of Meloidogyne mayaguensis compared with M. incognita, M. arenaria, M. javanica and M. floridensis on tomato in microplots in Florida. J Nematol. 2005;37:361.
17.Cetintas R, Kaur R, Brito Janete A, Mendes ML, Nyczepir A, Dickson D. Pathogenicity and reproductive potential of Meloidogyne mayaguensis and Meloidogyne floridensis compared with three common Meloidogyne spp. Nematropica. 2007;37(1):21-31.
18.Cirotto PA, Resende OF, Quintanilha AP, Carneiro Regina MDG. Resistência de cultivares de Capsicum annuum a Meloidogyne spp. In: Proccedings of the 27th Congresso Brasileiro de Nematologia. 2007;Universidade Federal de Goiás. Brasil; 2007.
19.Crozzoli R, Perichi G, Perez D, Espinosa M. Response of some legumes to Venezuelan population of root knot nematode Meloidogyne mayaguensis. Nematropica. 2006;36(2):120.
20.Decker H, Rodríguez Maria E. Uber das Auftretendes Wurzelgallennematoden Meloidogyne mayaguensis an Coffea arabica in Kuba. Wiss Z WPU Rostock N-Reihe. 1989;38(3):32-34.
21.Duponnis R, Gueye M, Mustapha A, Sene Veronique. Champignons contre nématodes lutte biologique au Sénégal. ORSTOM Actualites. 1997;54:35-39.
22.Duponnis R, Tabula TK, Cadet P. Étude des interactions entre trois espécés d´Acacia (Faidherbia albida Del., Acacia seyal Del., A. holosericea A Cunn. ex G. Don) et Meloidogyne mayaguensis au Sénégal. Can J Soil Science. 1997;77(3):359-365.
23.Duponnois R, Mateille T, Ba A. Potential effects of sahelian nematophagous fungi against Meloidogyne mayaguensis on tobacco (Nicotiana tabacum L. var. Paraguay x Claro). Ann Du Tabac, Section 2.1997;29:61-70.
24.Duponnois R, Chotte JL, Sall S, Cadet P. The effects of organic amendments on the interactions between a nematophagous fungus Arthrobotrys oligospora and the root-knot nematode Meloidogyne mayaguensis parasitizing tomato plants. Biology and Fertility of Soils. 2001;34:1-6.
25.Duponnois R, Mateille T, Gueye M. Biological characteristics and effects of two strains of Arthrobotrys oligospora from Senegal on Meloidogyne species parasitizing tomato plants. Biocontrol Sci and Technology. 1995;5:517-525.
26.Duponnois R, Mateille T, Sene V, Sawadogo A, Fargette Mireille. Effect of different West Africans species and strains of Arthrobotrys nematophagous fungi on Meloidogyne species. Entomophaga. 1996;41(3/4):475-483.
27.Fargette Mireille, Davies KG, Robinson MP, Trudgill DL. Characterization of resistance breaking Meloidogyne incognita - like populations using lectins, monoclonal antibodies and spores of Pausteria penetrans. Fundam Appl Nematol. 1994;17(6):537-542.
28.Fargette Mireille,Phillips M, Blok Vivian C, Waugh R, Trudgill D. An RFLP study of the relationships between species, populations
and resistance-breaking lines of tropical species of Meloidogyne. Fundam Appl Nematol. 1996;19(2):193- 200.
29.Fargette Mireille, Braaksma R. Use of esterase phenotype in the taxonomy of the genus meloidogyne. 3. A study of some "B" race lines and their taxonomic position. Revue Nématol. 1990;13(4):375-386.
30.Fernández E, Gandarilla Hortensia, Sariol H. Observaciones sobre una nueva forma de Meloidogyne Goeldi, que ataca el cafeto (Coffea arabica L.) en Cuba. En: 1ra Jornada Científica de Sanidad Vegetal; 1981. Cienfuegos. Tomo 3; 1981.
31.Fernández E, Acosta Oneida, García Isabel, Cuadra R, Rodríguez Mayra G, Sánchez Lourdes, et al. Manejo Integrado de los nematodos del género Meloidogyne en el cafeto. En: VIII Forum de Ciencia y Técnica; 1993. INISAV. SNT-Ciencia y SNTAF. Ciudad Habana.
32.Fernández E, Pérez M, Gandarilla Hortensia. Guía para disminuir infestaciones de Meloidogyne spp. mediante el empleo de cultivos no susceptibles. Boletín Técnico (INISAV). 1998;4(4):1-18.
33.Fernández R, Cabrera J, Rodríguez MC, Espárrago G, González JA, Báguena M, et al. "Guadajira" and "Gevora": Open-pollinated, procesing tomato cultivars resistant to Root Knot Nematodes and Fusarium Wilt. HortScience. 1999;34(2):356-357.
34.Franca Cristina A cultura da goiaba agoniza no vale do São Francisco. FCAP/UPE; 2005. (Consultado: 3 Jul 2007). Disponible en: http://www.portaldoagronegocio.com.br/index.php?p=texto&&idT=59.
35.Guimaraes LM, Moura RM, Pedrosa EMR. Parasitismo de Meloidogyne mayaguensis em diferentes espécies botanicas. Nematol Bras. 2003;27:139-145.
36.Hernández A, Fargette Mireille, Moliner V, Ramenason H, Decazy B, Sarah JL. Enzymatic characterization and reproductive fitness on Coffee of Root- Knot Nematode population from Central America. In: Proccedings of the 3th International of Nematology Congress; 1996. Gosier, Guadeloupe; 1996..
37.Hussey RS, Williamson Valerie M. Physiological and molecular aspects of nematode Parasitism. In: Barker K, Pederson G, Windham G, editors. Plant and Nematode Interactions. Agronomy Monograph No. 36. Madison, Wisconsin, USA; 1998. p. 87-108.
38.Inserra R, Brito Janete A. Nematodes of special interest. Triology. 2003;42(1):DPI - FDACS.htm
39.Jepson Susan. Identification of root-knot nematodes (Meloidogyne spp.). CAB. International. Wallingford, UK; 1987.
40.Jourand P, Rapior Sylvie, Fargette Mireille, Mateille T. Nematostatic activity of aqueous extracts of West African Crotalaria species. Nematology. 2004;6(5): 765-771.
41.Karssen Gerrit, Moens M. Root Knot Nematodes. In: Perry RN, Moens M, editors. Plant Nematology. CAB International; 2006. p. 59-90.
42.Kaur R, Brito Janete, Dickson DW, Stanley JD. First Report of Meloidogyne mayaguensis on Angelonia angustifolia. Plant Dis. 2006;90:1113.
43.Kermarrec A, Panoma S, Queneherve P, Mauleon H, Castagnone-Sereno P. Genetic variability of Meloidogyne mayaguensis isolates from the Caribbean. In: Brill, Netherland editor. Fourth International Congress of Nematology. 2002 Jun 8-13; Tenbel, La Galletas, Arona, Tenerife. Canary Islands, Spain; 2002. p. 139.
44.Lavigne C, Pigeat C, Bertin Y, Ducelier D, Jannoyer M. El injerto de la guayaba (Psidium guajava) en el cas (P. friedrichstalianum) protege contra la sensibilidad al nematodo (Meloidogyne mayaguensis). En: Actas del II Simposio Internacional de Fruticultura Tropical. FRUTICULTURA 2007. 17 - 21 de septiembre. Ciudad de La Habana, Cuba; 2007.
45.Lima IM, Dolinski CM, Sousa RM. Dispersão de Meloidogyne mayaguensis em goiabais de São João da Barra (RJ) e relato de novos hospedeiros dentre plantas invasoras e cultivadas. En: Actas del XXIV Congresso Brasileiro de Nematologia. Petrolina, Brasil, 2003. p. 139.
46.Lima IM, Sousa RM, Silva CP, Carneiro Regina MDG. Meloidogyne spp. from preserved areas of Atlantic Forest in the State of Rio de Janeiro, Brazil. Nematol Bras. 2005;29(1):31-38.
47.Lugo Zunilde, Crozzoli R, Molinari S, Greco N, Perichi G, Jiménez-Pérez N. Patrones isoenzimáticos de poblaciones venezolanas de Meloidogyne spp. Fitopatol Venez. 2005;18(2):26-29.
48.Medeiros JE, Moura RM, Pedrosa EMR. (2003): Novas plantas hospedeiras de Meloidogyne mayaguensis. En: Jornada de Ensino, Pesquisa e Extensão da Universidade Federal Rural de Pernambuco. Recife, Brasil; 2003.
49.Molinari S, Lamberti F, Crozzoli R, Sharma SB, Sanchez Lourdes (2005): Isozyme patterns of exotic Meloidogyne spp. populations. Nematol Medit. 2005;33:61-65.
50.Moreira WA, Pereira AVS, Magalhães EE, Lopes DB, Barbosa FR, Moura AOS. Avaliação da eficiência de abamectina e do cadusafos na redução populacional de nematóides-das-galhas, na cultura da goiabeira. En: Actas de la XLIX Reunião Anual da Sociedade Interamericana de Horticultura Tropical e Horticultura Tropical em Regiões Semi-Áridas. Fortaleza, CE, Brasil; 2003. p. 141.
51.Moura RM, Pedrosa EMR, Guimarães LMP. Meloidogyne mayaguensis parasitims on different plant species. Nematol Bras. 2003;27(2):139-145.
52.Perichi G, Crozzoli R, Lugo Zunilde. Diferenciación morfológica de poblaciones venezolanas de Meloidogyne mayaguensis y de Meloidogyne incognita. Nematropica. 2006;36(2):140.
53.Pest Alert. NAPPO Phytosanitary Alert System. Un nematodo de la raiz nuevo para EU; 2002. (Consultado: 28 Jun 2002). Disponible en: http://doacs.state.fl.us/~pi/enpp/nema/m-ayaguensis.html.
54.Queneherve P, Bertin Y, Chabrier C. Arachis pintoi: ¿una planta de cobertura para los bananales? Ventajas e inconvenientes desde un punto de vista nematológico. Infomusa. 2002;11(1):28-30.
55.Queneherve P. (): El estudio de microorganismos parásitos en plantas es fundamental para la protección del medio ambiente; 2006.(Consultado: 23 Jul 2007). Disponible en: http://www.mx.ird.fr/imprimer_article.php3?id_ article=2509.
56.Rammah A, Hirschmann Hedwig. Meloidogyne mayaguensis n. sp. (Meloidogynidae), a Root-Knot Nematode from Puerto Rico. J Nematol. 1988;20(1):58-69.
57.Rammah A. Morphological and taxonomic studies of certain populations of Meloidogyne arenaria and M. javanica. PhD. Thesis. North Carolina State University. In: Dissertation Abstracts International. 1989;49(7):2447.
58.Randig, O.; Carneiro, Regina M.D.G. y Castagnone-Sereno, P. (2004): Identificacao das principais species de Meloidogyne parasitas do cafeeiro no Brasil com Marcadores SCAR-Cafè em Multiplex-PCR. Nematol. Bras. 28(1): 1-10.
59.Rodríguez Mayra G. Identificación y caracterización de Meloidogyne mayaguensis en el cafeto en Cuba. Tesis en opción al Grado científico de Doctor en Ciencias Agrícolas. Universidad Agraria de La Habana. Cuba; 2000.
60.Rodríguez Mayra G, Sánchez Lourdes, Rodríguez I, Enrique R. Detalles acerca de una nueva especie del género Meloidogyne que parasita el cafeto en Cuba. Rev Protección Veg. 1989;4: 88-90.
61.Rodríguez Mayra G, García Isabel, Gandarilla Hortensia. Internal quarantine for Meloidogyne mayaguensis in Cuba. In: Proccedings of the XXXVI Annual Meeting of Organization of Nematologists of Tropical America. Puerto Vallarta, Jalisco. Mexico; 2004. p. 70-71.
62.Rodríguez Mayra G, Sánchez Lourdes, Rowe Janet. Host status of agriculturally important plant families to root-knot nematode Meloidogyne mayaguensis in Cuba. Nematropica. 2003;33(2):125-130.
63.Rodríguez Mayra G, Sánchez Lourdes, Enrique R. Evaluación de hospedantes para el monitoreo de poblaciones de Meloidogyne spp., presentes en el cafeto. Rev Protección Veg. 1999;14(1):51-54.
64.Rodríguez Mayra G, Rodríguez I, Sánchez Lourdes. Especies del género Meloidogyne que parasitan el cafeto en Cuba: Distribución geográfica y sintomatología. Rev Protección Veg. 1995;10:123-128.
65.Santos MS, Abrantes Isabel. Identification of Root-Knot nematodes. In: A Workshop Manual for Research on Verticillium chlamydosporium as a biological control agent for Root-Knot Nematodes. EU Project FAIR5-PL97-3444. IACR-Rothamsted, UK; 1998. p. 64-71.
66.Siqueira KMS, Torres GRC, Pedrosa Elvira MR, Moura RM. Eclosão de Meloidogyne incognita, Meloidogyne javanica e Meloidogyne mayaguensis em Lixiviados de Caupi Associado a Glomus etunicatum e Bradyrhizobium sp. Fitopatol bras. 2004;29(3):259-262.
67.Sociedad de Nematólogos (SON). Meloidogyne mayaguensis Pest Information. Exotic Nematode Plant Pest of Agricultural and Environmental Significance to the United State; 2002. (Consultado: 22 Jul 2002). Disponible en: http://www.son.org/Meloidogyne mayaguensis Pest Information.htm.
68.Sousa RM, Nogueira MS, Lima IM, Melarato M, Dolinski CM. Manejo do nematóide das galhas da goiabeira em Sao Joao da Barra (RJ) e relato de novos hospedeiros. Nematol Bras. 2006,30(2):165-169.
69.Torres GRC, Sales R, Nerivania Vitorina, Rehn C, Pedrosa Elvira MR, Moura RM. Ocorrência de Meloidogyne mayaguensis em Goiabeira no Estado do Ceará. Nematol Bras. 2005;29(1):105-107.
70.Torres GRC, Covello Vitorina N, Sales R, Pedrosa Elvira MR, Moura RM. Meloidogyne mayaguensis em Psidium guajava no Rio Grande do Norte. Fitopatol bras. 200429(5):570.
71.Trudgill D, Bala G, Blok Vivian C, Daudi A, Davies KG, Gowen SR, et al. The importance of tropical nematodes (Meloidogyne spp.) and factors affecting the utility of Pausteria penetrans as a biocontrol agent. Nematology. 2000;2(8):823-845.
72.UENF. Pesquisadores discutem como conter parasita de goiabeiras: Praticamente invisibel, nematóide ataca variedades mais plantadas no Norte Fluminense. Comunicación de: Universidade Estadual do Norte Fluminense Darcy Riveiro; 2007. (Consultado: 22 Jul 2007). Disponible en: http://www.uenf.br.
73.UFRRJ. Infecção natural de Cucurbita moschata por Meloidogyne mayaguensis, no Estado do Rio de Janeiro. En: Resumos do XXVI Congresso Brasileiro de Nematologia; 2007. (Consultado: 6 Jun 2007). Disponible en: http://www.ufrrj.br/institutos/ib/denf/redepublicacao.htm.
74.Villain L, Sarah JL, Hernandez A, Charmenat P, Bertrand B, Anthony F, et al. Biodiversity of Root Knot Nematodes, Meloidogyne spp., on Coffee in Central America. ASIC-CAFÉ; 2006. (Consultado: 2 Jul 2007). Disponible en: http://www.asic-cafe.org/pdf/abstract/PA189_2006.pdf.
75.Willers P. Die aalwurmprobleem by koejawels en moontlike gebruik van kadusafos aalwurmdoder. (Guava Koejawel). ITSC Information Bulletin. 1997;293:10-11.
76.Willers P. First record of Meloidogyne mayaguensis Rammah and Hirschmann, 1988: Heteroderidae en commercial crops in the Mpumalanga Province, South Africa. Subtropica. 1997;18(2):19-20.
1 MSc. Ana María Cassasa. Universidad de Zulia. Venezuela.
2 Sánchez, Lourdes y Mayra G. Rodríguez. (1998): Complejo de especies de Meloidogyne presentes en tabaco y evaluación de plantas indicadoras para su detección. Informe de investigación. CENSA. 6 pp.
3 Dr. Leopoldo Hidalgo. Centro Nacional de Sanidad Agropecuaria (CENSA). Informaciones obtenidas en intercambio con especialistas de la Empresa Brasilera de Pesquisas Agropecuarias, 2007.
(Recibido 3-8-2007; Aceptado 18-9-2007)


{kind=link}
{kind=link}