










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión On-line ISSN 2224-4697
Rev. Protección Veg. v.24 n.2 La Habana Mayo-ago. 2009
TRABAJO ORIGINAL
ACTIVIDAD ANTAGÓNICA FRENTE A Pyricularia grisea (Sacc.) Y FITOESTIMULACIÓN EN EL CULTIVO DEL ARROZ DE CEPAS AUTÓCTONAS DE Pseudomonas putida (Trev.)
ANTAGONISTIC ACTIVITY AGAINST Pyricularia grisea (Sacc.) AND GROWTH STIMULATION IN RICE OF Pseudomonas putida (Trev.) NATIVE STRAINS
Narovis Rives*, Yanelis Acebo**, M. Almaguer**, J.C. García** y Annia Hernández**
*Instituto de Investigaciones del Arroz. Km 16½, Autopista Novia del Mediodía, Bauta, La Habana, Cuba. Correo electrónico: narovis@fbio.uh.cu; **Facultad de Biología. Universidad de La Habana. Calle 25, # 455, Plaza de la Revolución, Ciudad de La Habana, Cuba. Correo electrónico: annia@fbio.uh.cu
RESUMEN
La aplicación de inoculantes de origen bacteriano en cultivos de importancia económica, constituye una alternativa ecológica que permite la obtención de mayores rendimientos y la conservación del medio ambiente. La obtención y caracterización de cepas autóctonas, potencialmente eficientes en la promoción del crecimiento vegetal y el control biológico de patógenos constituye un paso esencial para el desarrollo de inoculantes microbianos. Esta investigación se desarrolló con el objetivo de caracterizar cepas de Pseudomonas putida previamente aisladas del cultivo del arroz variedad J-104 en cuanto a su actividad promotora del crecimiento vegetal y su efecto antagónico frente a Pyricularia grisea. Se realizaron experimentos in vitro para detectar la producción de compuestos indólicos, producción de sideróforos, actividad antagónica contra P. grisea y el efecto estimulador del crecimiento vegetal de las cepas, en plantas de arroz de 12 días de edad sembradas en tubos de ensayo con medio Murashige y Skoog (MS). Los resultados mostraron la capacidad de producir metabolitos del tipo ácido 3-indolacético (45,8 - 56 µg.mL-1) y sideróforos (7,3-39,4 µM) por las cepas en estudio, las que manifestaron efecto antagónico frente a P. grisea (82-86% de inhibición del crecimiento fúngico) y como promotor del crecimiento vegetal, esto último, a través de su efecto sobre algunos parámetros del crecimiento evaluados (altura de la planta, largo de la raíz, masa fresca aérea y de raíz), destacándose un comportamiento integral en las cepas de P. putida AJ9, AJ14, AJ20 y AJ18. Desde el punto de vista práctico estos resultados indican las potencialidades de las cepas estudiadas para su utilización como inoculantes microbianos en beneficio del cultivo del arroz.
Palabras clave: arroz; Pseudomonas putida; Pyricularia grisea; promoción del crecimiento; actividad antagónica
ABSTRACT
The application of bacterial inoculants in crops economicaly important crops has been an ecological alternative to improve crop yield and to preserve the environment. The obtaining and characterization of native strains associated with several crops and potentially efficient to promote plant growth and to biocontrol the phytopathogens are key steps to the development of microbial inoculants. This research was carried out to characterize strains of Pseudomonas putida, isolated from rice (Oryza sativa L. var. J-104) as phytostimulators and antagonistic agents against Pyricularia grisea. In vitro experiments were carried out to detect the production of indolic compounds and siderophores, antagonistic activity against P. grisea and to assess the phytostimulating effect of the strains in 12-day-old rice plants grown in Murashige and Skoog medium. The results showed the ability of the strains to produce indolacetic acid-like metabolites (45,8 to 56 µg·mL-1) and siderophores (7,3 to 39,4µM) . The strains showed antagonistic effect against P. grisea (82-86% of fungal growth inhibition) and were also capable of promoting plant growth (by increase in plant height, root length, root fresh weight and aerial part fresh weight), standing out the strains AJ9, AJ14, AJ20 and AJ18 for the best performance. From the practical point of view, these results show the potential of these strains to be used as inoculants in rice crop.
Key words: rice; Pseudomonas putida; Pyricularia grisea; plant growth promotion; antagonistic activity
(Recibido 8-10-2008; Aceptado 23-1-2009)
INTRODUCCIÓN
El cultivo del arroz (Oryza sativa L.) se enfrenta a graves problemas, entre ellos se destacan: la tasa descendente de crecimiento de las cosechas, la escasez de mano de obra y la contaminación del medio ambiente (1). Uno de los factores que inciden en la obtención de bajos rendimientos en este cultivo lo constituye la aparición de enfermedades, una de ellas; la piriculariosis o añublo del arroz con una alta frecuencia de aparición en la mayoría de los agroecosistemas. Si las condiciones ambientales, el grado de susceptibilidad de la variedad, así como la tecnología de cultivo empleada son favorables para que se desarrolle su agente causal (Pyricularia grisea Sacc.), esta puede provocar hasta el 100% de reducción de los rendimientos agrícolas (2). Es por ello, que la búsqueda de alternativas ecológicas que permitan aumentar los rendimientos de este importante cultivo, sin provocar afectaciones en el medio ambiente, constituye una prioridad para los investigadores, especialistas y técnicos arroceros a nivel mundial.
La aplicación de inoculantes de origen bacteriano, ha constituido una alternativa ecológica que permite la obtención de mayores rendimientos y la conservación del medio ambiente. En este sentido, la obtención y caracterización de cepas autóctonas asociadas a estos cultivos, potencialmente eficientes en la promoción del crecimiento vegetal y control biológico de fitopatógenos, constituye un paso esencial para el desarrollo de estos productos biológicos (3).
Las especies del género Pseudomonas se encuentran entre las más estudiadas porque tienen la capacidad de producir reguladores del crecimiento vegetal y otros metabolitos con efecto antagónico y represivo del crecimiento de patógenos en la rizosfera (4). También este género constituye un excelente ejemplo de la combinación de múltiples mecanismos para ejercer un efectivo control biológico (5).
Por lo anteriormente expuesto, este trabajo se desarrolló con el objetivo de caracterizar 10 cepas de Pseudomonas putida (Trev.) previamente aisladas del cultivo del arroz variedad J-104 en cuanto a la actividad promotora del crecimiento vegetal en el cultivo del arroz y su efecto antagónico frente a P. grisea.
MATERIALES Y MÉTODOS
Cultivos microbianos: En este estudio se utilizaron 10 cepas de Pseudomonas putida previamente aisladas de la rizosfera de plantas de arroz de la variedad J-104 sembradas en el área experimental del Instituto de Investigaciones del Arroz (IIArroz, La Habana) sobre suelo Gley Vértico Crómico-Nodular Ferruginoso. Las mismas se identificaron mediante el sistema API 20 NE (6) y se conservaron en medio Luria Bertani (LB) con glicerol al 40% a una temperatura de -20ºC.
Se utilizó la cepa autóctona de P. grisea AAM-246 que pertenece a la Colección de Cultivos Fúngicos de la Facultad de Biología con el número 356. Este cultivo se encontraba conservado en medio Papa Dextrosa Agar (PDA) (Biocen) a 28oC.
Producción de ácido 3-indolacético (AIA)
En tubos de ensayos que contenían 5mL de Caldo Triptona Soya suplementado con triptófano (0,1g.L-1), se inocularon con 500 mL de una suspensión bacteriana de cada aislado, ajustada a una concentración celular de 108 cel.mL-1, comparado con el tubo 0,5 de la escala de MacFarland. Los tubos se incubaron por 48 horas a 30°C, posteriormente el contenido se centrifugó a 5000 rpm por 15 minutos (Centrifuga 5702, Eppendorf).
Para la detección cualitativa, se prepararon suspensiones bacterianas de cada aislado en solución salina fisiológica, ajustando la concentración celular a 108cel.mL-1. Se tomaron 500 mL de cada suspensión, se inocularon en tubos de ensayo con 5 mL de Caldo Triptona Soya suplementado con triptófano (0.1g·L-1) y se incubaron a 24 horas en agitación (150 rpm) a 28ºC. Posteriormente, los cultivos se centrifugaron (Centrifugue 5702, Eppendorf) a 5000 rpm durante 15 minutos, se tomaron 10 mL de los cultivos libres de células y se concentraron en un rotoevaporador hasta su secado total (Büchi B-490). Luego se añadieron 100 µL de etanol absoluto a cada tratamiento. Las muestras (10 µL) correspondientes a las cepas y un patrón de AIA sintético se aplicaron en placas comerciales para cromatografía en Sílica Gel 60 F 254 (MERCK). Se utilizó isopropanol-amonio-agua (80-5-15) como sistema de solventes y el reactivo de Salkowski como revelador.
Para la determinación cuantitativa de AIA, a los cultivos libres de células se les añadió el reactivo de Salkowski (600mL de H2SO4 18M, 4,5g de FeCl3 anhidro y se completó hasta 1L con agua destilada), en una relación 1:1. Los cultivos se incubaron a temperatura ambiente y oscuridad durante 30 minutos, luego se leyó la absorbancia a 530 nm en espectrofotómetro (Genesys 20, Thermospectronic). Paralelamente se elaboró una curva patrón con AIA sintético en concentraciones de 5 µg.mL-1 a 100 µg.mL-1. El experimento se repitió tres veces con cinco réplicas por cepa.
Producción de sideróforos
En el ensayo para la detección de sideróforos se empleó el medio King B sólido, mediante la metodología descrita por Schwyn y Neilands (7). Se utilizó la solución reveladora CAS (Cromo Azurol S) que indica la producción de este metabolito mediante un cambio de coloración en el medio de cultivo de azul a naranja. El experimento se repitió tres veces con cinco réplicas por cepa.
Para la cuantificación de este metabolito, se inocularon tubos con 5mL de caldo Casamino Ácido (Ácido Casamínico 5g·L-1, K2HPO4 1g·L-1, MgSO4 • 7H2O 1.55g·L-1, H20d 1L) con 500 µL de una suspensión bacteriana en solución salina fisiológica de cada aislado, ajustada a una concentración celular de 108cel.mL-1. Los cultivos se colocaron en zaranda orbital durante 48 horas a 150 rpm y 37oC. Se determinó la absorbancia a 595 nm para determinar la concentración celular y a 405 nm en el sobrenadante para cuantificar los metabolitos del tipo sideróforos en espectrofotómetro (Genesys 20, Thermospectronic).
Determinación de la actividad antagónica de las cepas seleccionadas frente a Pyricularia grisea
Se realizaron dos bioensayos in vitro para determinar el efecto antagónico de las cepas en estudio frente P. grisea. En el primer experimento in vitro se siguió la metodología descrita por Bashan et al. (8). Para ello el hongo se sembró previamente en medio de cultivo PDA mediante siembra a profundidad (9) y se incubó a 30ºC durante 7 días. Se realizaron suspensiones bacterianas en solución salina fisiológica estéril, ajustando la concentración celular a 108 cel.mL-1, según la escala de McFarland. Se incubaron por 48 horas a 28ºC en agitación a 125 rpm, (zaranda orbital C1 de New Brunswick Scientific). A las 48 horas, se extrajeron 100 µL de estos cultivos y se sembraron por diseminación mediante espátula de Drigalsky (9) en placas Petri de 9 cm de diámetro con medio King B. Las placas Petri previamente sembradas con el hongo se perforaron utilizando un obturador de 5 mm de diámetro. Los discos se colocaron en el centro de las placas sembradas con la bacteria, a razón de un disco para cada placa y cinco réplicas por tratamiento, luego se incubaron a 30ºC durante 7 días. La actividad antagónica de las bacterias se determinó a través de mediciones del diámetro de crecimiento del hongo en presencia del antagonista y un control donde no se aplicó este último. Con estos datos se determinó el porcentaje de inhibición de las cepas sobre P. grisea, según metodología descrita por Hernández (6). El expe
rimento se repitió tres veces a razón de tres réplicas por tratamiento.
En el segundo experimento in vitro se siguió la metodología descrita anteriormente, pero las bacterias se sembraron con hisopos estériles en forma circular, utilizando una plantilla de un diámetro de 55 mm, en el medio King B. Los discos del hongo se colocaron de igual forma en el centro de las placas Petri de 9 cm de diámetro. El porcentaje de inhibición se determinó mediante la metodología descrita por Hernández (6), utilizando como control, el crecimiento del hongo en ausencia de la bacteria. El experimento se repitió tres veces a razón de tres réplicas por tratamiento.
Determinación del efecto de las cepas de P. putida sobre la estimulación del crecimiento vegetal en el cultivo del arroz
Se realizaron suspensiones bacterianas en solución salina fisiológica estéril, ajustando la concentración celular a 108 cel.mL-1, según la escala de McFarland, tomándose cada una como un tratamiento. Se estableció como tratamiento control de lasolución salina fisiológica estéril sin inocular. Se desinfectaron 100 semillas de arroz variedad J-104 utilizando hipoclorito de calcio al 5% durante 20 minutos, según metodología descrita por Hernández et al. (10) y se transfirieron individualmente a tubos de cultivos de 20 x 3 cm que contenían 15 mL de medio Murashige y Skoog (MS) (11).
Posteriormente se adicionaron 100 µL de caldos de cultivos libres de células, procedentes de las suspensiones bacterianas (500 mL de cada suspensión, se inocularon en tubos de ensayo con 5 mL de medio King B líquido, se incubaron durante 24 horas en agitación a 150 rpm a 28ºC y posteriormente se centrifugaron (Centrifugue 5702, Eppendorf) a 5000 rpm durante 15 minutos) y el control a los tubos de medio MS y se incubaron a temperatura ambiente en régimen de fotoperíodo (14 horas de luz y 10 horas oscuridad). A los 12 días, las plántulas se sacaron de los tubos con mucho cuidado, se lavaron con agua destilada y se secaron con papel de filtro (Whatman). Posteriormente se procedió a separar la parte aérea de las plantas, de la raíz, para determinar los diferentes parámetros de crecimiento (altura de las plantas, largo de la raíz, masa fresca aérea y de la raíz) con la ayuda de un pie de rey (mm) (Würth, Alemania) y una balanza analítica (Sartorius, USA). Para la determinación del peso seco, el material vegetal se colocó en un horno (Memmert, Bélgica) a 70ºC durante tres días. Se establecieron 10 réplicas por tratamiento y el experimento se repitió tres veces.
Análisis biométrico
De forma preliminar, a todas las variables se les realizó la prueba de normalidad de Shapiro y Wilk (12) y homogeneidad de varianza, contenidas en el paquete estadístico Statistica 6.1. A partir de estos resultados, se decidió realizar los análisis paramétricos y no paramétricos según correspondiera.
A los datos que presentaron distribución normal, se les realizó la prueba de Student-Newman-Keuls (SNK) paramétrica. A los datos que no presentaron una distribución normal, se les realizó la prueba de Kruskal-Wallis completado por la prueba de comparación de Mann-Whitney y la correspondiente corrección de Bonferroni (13) o la prueba SNK no paramétrica. Todas las pruebas estadísticas realizadas están contenidas en el paquete estadístico Statistica 6.1
RESULTADOS Y DISCUSIÓN
Producción de ácido 3-indolacético (AIA)
Dentro de los mecanismos por los cuales las rizobacterias pueden manifestar su acción se destacan la producción de compuestos indólicos, a los cuales se les atribuye el incremento en el desarrollo y rendimiento de diversas especies de plantas.
Al realizar el análisis cromatográfico se observaron manchas de color rojizas en las placas de cromatografía que se correspondieron con el patrón de AIA sintético, lo que corrobora la producción de este metabolito por las cepas seleccionadas y la eficacia de la técnica colorimétrica utilizada (Fig. 1).
De manera general, las cepas en estudio demostraron poseer altos niveles en la producción de auxinas, obteniéndose valores de AIA entre 45,8 y 56 µg.mL-1 (Fig. 2). Los mayores niveles en la producción de esta auxina se alcanzaron con la cepa de P. putida AJ16, aunque sin diferir significativamente de las restantes cepas.
Los niveles de producción de esta hormona obtenidos en el presente trabajo son elevados si se comparan con los encontrados por Hernández (6) y Trujillo (14), con cepas de Pseudomonas fluorescens (5,90-18,3 µg.mL-1) aisladas de la rizosfera del cultivo del maíz.
Asimismo, estos resultados son superiores a los obtenidos por Gravel et al. (15) en una investigación sobre el posible papel del AIA en la estimulación del crecimiento en plantas de tomate inoculadas con una cepa de Pseudomonas putida estudiada, en la que obtuvieron una producción de 23,4 µg.mL-1 de este metabolito.
Es importante señalar, que la técnica utilizada para la cuantificación de sustancias estimuladoras del crecimiento del tipo auxinas, puede presentar interferencias por la presencia de otros indoles. No obstante, es empleada por muchos autores para estos fines (6,14,16,17).
Producción de sideróforos
Mediante el método cualitativo de detección de sideróforos se demuestra que todas las cepas tienen la capacidad de producir este metabolito, lo que se denota por la presencia de halos de color naranja en las placas de medio de cultivo con el indicador Cromo Azurol S (Fig. 3).
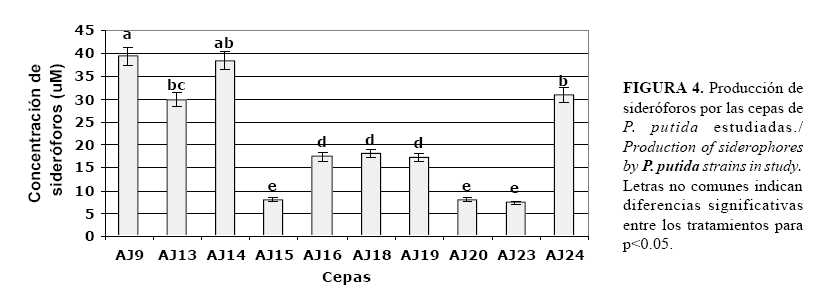
La producción de sideróforos por cepas de rizobacterias pertenecientes a diferentes grupos microbianos y su función en el control biológico de enfermedades de plantas ha sido informada por varios autores (6, 18, 19). En este trabajo se demuestra que todas las cepas de P. putida utilizadas en el estudio, tienen la capacidad de producir sideróforos bajo condiciones limitantes de hierro (Fig. 4).
Los mayores niveles de producción de sideróforos se obtienen en las cepas AJ9, AJ13, AJ14 y AJ24, destacándose las cepas AJ9 y AJ14 con las mayores producciones. Se ha observado que las rizobacterias
Letras no comunes indican diferencias significativas entre los tratamientos para p<0.05. con la capacidad de sintetizar estos compuestos son altamente efectivas en el control biológico de fitopatógenos (19). Al formarse el complejo hierro-sideróforo, disminuye la disponibilidad de este en el suelo, ya que los receptores proteicos que reconocen el complejo son específicos de cada especie bacteriana, estableciéndose una competencia por el hierro frecuentemente fatal para el patógeno, que no posee mecanismos propios tan efectivos para la adquisición de hierro. Este mecanismo, sin embargo no perjudica a las plantas colonizadas, por el contrario, se ha planteado la existencia de mecanismos de incorporación del hierro solubilizado por los sideróforos (20).
Experimentos en condiciones de invernadero demostraron que la cepa B10 de P. putida tiene la capacidad de suprimir la pudrición provocada por Fusarium, sin embargo este efecto se pierde al adicionarle hierro al suelo (que reprimió la producción de sideróforos), lo que demuestra el papel determinante que desempeñan los sideróforos en el control biológico de patógenos (21). Buysens et al. (18) también demostraron la efectividad de los sideróforos en el control de enfermedades causadas por algunas especies de Fusarium y Pythium.
La selección de cepas eficientes en la producción de metabolitos que actúan como estimuladores del crecimiento vegetal y como agentes de control biológico de patógenos es un aspecto de gran interés dentro de la biotecnología agrícola; aspecto que podría estar relacionado con los efectos beneficiosos obtenidos en las plantas de interés económico al inocular rizobacterias del género Pseudomonas (6, 16, 22, 23).
Determinación de la actividad antagónica de las cepas seleccionadas frente a Pyricularia grisea
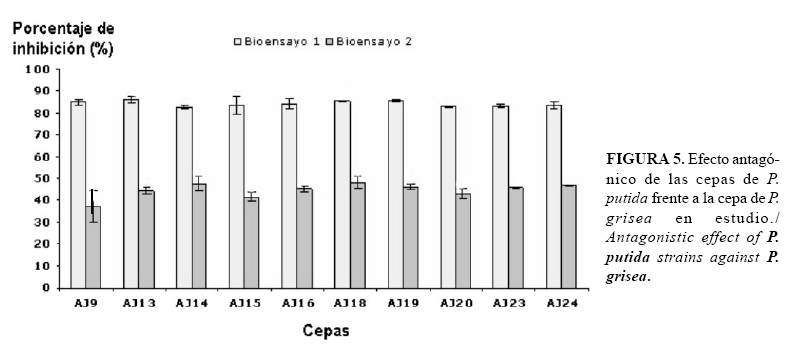
Las cepas de rizobacterias estudiadas mostraron efecto antagónico frente a P. grisea (Fig. 5), lo que corrobora el efecto de las bacterias asociadas a las plantas en el control biológico de patógenos que afectan a cultivos de importancia económica, tales como el arroz (24, 25).
Al establecerse la interacción entre los antagonistas bacterianos y la cepa de P. grisea se observaron altos porcentajes de inhibición en el hongo, lográndose en el primer bioensayo entre el 82 y el 86% de inhibición del crecimiento y en el segundo del 37 al 48%. El análisis estadístico de los valores no mostró diferencias significativas entre el comportamiento de las cepas (Fig. 6). Sin embargo, se debe destacar que en el primer bioensayo se obtiene mayores porcentajes de inhibición que en el segundo, lo que podría estar relacionado con la eficiencia del protocolo utilizado, dado fundamentalmente por la forma en que se establece la interacción entre el antagonista y el patógeno.
En los bioensayos cualitativos influye, entre otros factores, la concentración del inóculo del antagonista, el grosor del agar, la concentración y dureza del medio de cultivo así como las condiciones de incubación (26). En este estudio, a pesar de utilizarse el mismo medio de cultivo y condiciones experimentales en ambos bioensayos, las diferencias obtenidas podrían estar relacionadas con la concentración del inóculo, dado por el método de siembra utilizado y la diferencia en cuanto a la exposición del hongo al efecto del antagonista. Otros aspectos que podrían haber influido en los resultados son: la difusión de los metabolitos en el medio, la complejidad del agar empleado, así como el tamaño de las moléculas.
El efecto antagónico que se pone de manifiesto entre los antagonistas microbianos y el hongo, podría estar relacionado con la producción de metabolitos de naturaleza antibiótica. Investigaciones relacionadas han señalado la capacidad que tienen las cepas de Pseudomonas de suprimir patógenos presentes en la rizosfera a través de la producción de 2,4 dimetyl-phloroglucinol y pirrolnitrina (27). Asimismo, algunos autores han demostrado la presencia de alcaloides del grupo de las quinolinonas, que exhiben actividad antibiótica con efecto antifúngico, en rizobacterias (27).
Es importante destacar que en este caso, también podrían haber influido los sideróforos, si se tiene en cuenta que el medio de cultivo utilizado para el desarrollo de los bioensayos posee bajo contenido de hierro, lo que favorece la producción de este tipo de metabolito.
Diversos autores han encontrado resultados favorables al aplicar rizobacterias como agentes de control biológico en otros sistemas planta-patógeno. Por ejemplo, Hernández et al. (24) al aplicar las cepas de P. fluorescens WCS 417, J-143 y B. cepacia 0057 al cultivo del frijol, lograron disminuir la cantidad de lesiones provocadas por Colletotrichum lindemuthianum (Sacc. & Magn.) Pujol et al. (28), aplicaron la cepa de Pseudomonas fluorescens EPS62e para controlar los altos índices de afectación provocados por Erwinia amylovora (Burrill) en plantas de pera, obteniendo resultados satisfactorios. Asimismo se ha señalado que en el cultivo del maíz en los tratamientos donde las semillas fueron tratadas con cepas de P. fluorescens y Burkholderia cepacia, se observó reducción de los altos índices de infección provocados por Fusarium verticillioides (Sacc.) Nirenb. (6). Sin embargo, se conoce poco de la factibilidad de aplicar rizobacterias para ejercer el control biológico de P. grisea y no existen antecendentes que demuestren el efecto antagónico de rizobacterias del género Pseudomonas frente a este patógeno.
De modo general, los resultados in vitro evidencian las potencialidades de las cepas en estudio en la producción de metabolitos que desempeñan una importante función en el control biológico de patógenos, sin embargo, es importante realizar estudios de interacción planta-bacteria para conocer los verdaderos efectos beneficiosos que se ponen de manifiesto en la interacción con los cultivos.
Determinación del efecto de las cepas de P. putida sobre la estimulación del crecimiento vegetal en el cultivo del arroz
En las Figuras 7, 8, 9 y 10 se muestra el efecto promotor del crecimiento de las cepas de P. putida sobre plantas de arroz de 12 días de edad, demostrándose la aparición de cambios fenotípicos en la planta, fundamentalmente en el sistema radical, lo que se denota por un aumento en el número de raíces adventicias y laterales.
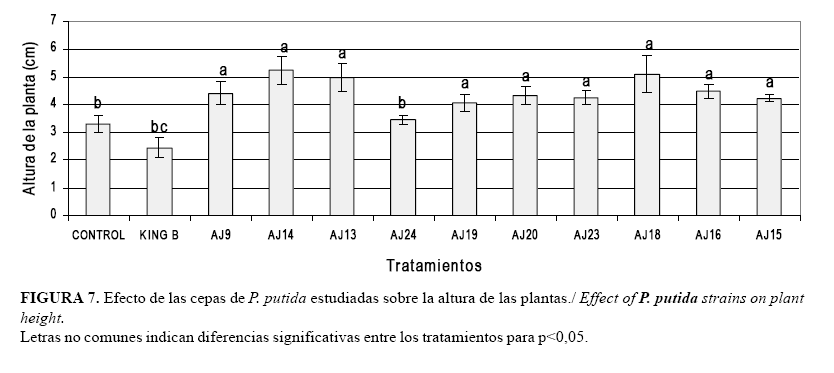
La evaluación de la altura de las plantas de arroz inoculadas con las cepas estudiadas produjo diferencias significativas entre los tratamientos (Fig. 7). Los mayores valores de las mediciones de este parámetro se obtuvieron en los tratamientos donde se aplicaron las cepas AJ9, AJ14; AJ13, AJ19, AJ20, AJ23, AJ18, AJ15, AJ16 que no presentaron diferencias significativas entre ellos, aunque sí con el resto de los tratamientos. A los controles correspondieron los valores más bajos sin mostrar diferencias significativas con el tratamiento donde se inoculó la cepa AJ24.
En cuanto a la masa fresca aérea de las plantas de arroz (Fig. 8), se observaron diferencias significativas entre los tratamientos. Se destacan con el mejor comportamiento las cepas AJ9, AJ14, AJ19, AJ20 y AJ18.
De modo general, los resultados indican que existe cierta relación entre la altura de las plantas y la masa fresca aérea, lo que ha sido informado por otros autores. En este sentido, Rives et al. (3) demostraron la existencia de una relación directa entre estos parámetros, mediante la inoculación de semillas de arroz con cepas de Azospirillum y Herbaspirillum con actividad promotora del crecimiento vegetal. Un comportamiento similar fue informado por Pazos (29), al inocular plantas de arroz con cepas nativas de Azospirillum brasilense.
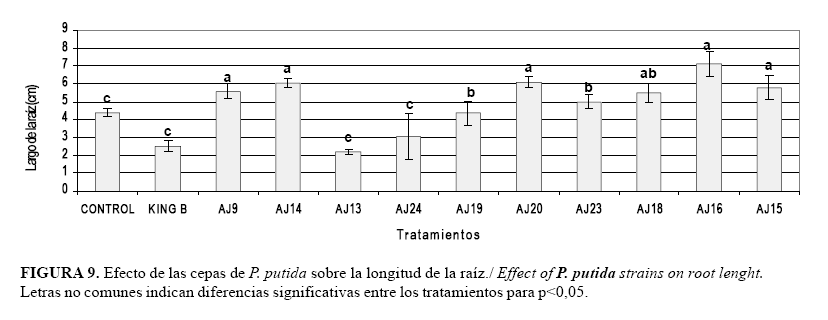
En este trabajo se demostró que la inoculación bacteriana favorece el desarrollo del sistema radical. El efecto de los diferentes tratamientos sobre la longitud de la raíz principal de las plantas de arroz se muestra en la Figura 9, evidenciándose diferencias significativas entre los tratamientos. Se destacan los tratamientos inoculados con las cepas AJ9, AJ14, AJ20, AJ18; AJ16 y AJ15 que presentaron los mayores valores y diferencias significativas con el resto de los tratamientos.
Asimismo, se observa un efecto beneficioso de la inoculación bacteriana sobre la masa fresca de las raíces de las plantas de arroz de 12 días de edad (Fig. 10). Se destacan las cepas AJ18 y AJ15 por los mayores incrementos.
Efectos similares en cuanto a incrementos en la masa fresca aérea y de raíz al inocular gramíneas con rizobacterias han sido informados por diversos autores. Hernández et al. (24) y Egamberdiyeva (30), demostraron que la inoculación de plantas de maíz con rizobacterias provoca incrementos en el largo de la raíz, contribuyendo de esta forma a una mayor absorción de nutrientes del suelo, ya que al aumentar el sistema radical se produce un incremento posterior en la adquisición de sustancias nutritivas y agua por la planta. Un comportamiento similar fue informado por Rives et al. (3), al inocular plantas de arroz con cepas autóctonas de Azospirillum brasilense.
Estos resultados podrían estar relacionados con la producción de sustancias reguladoras del crecimiento por las cepas en estudio. En este sentido, se demostró que las cepas utilizadas en el bioensayo producen auxinas del tipo AIA, la que pudiera estar involucrada con los resultados obtenidos ya que la misma promueve el crecimiento y desarrollo de las plantas y se ha demostrado que la influencia de este metabolito en el desarrollo y las funciones de la raíz en plantas inoculadas, es probablemente uno de los factores de mayores beneficios para los cultivos, pudiendo desempeñar una función vital en los efectos obtenidos en plantas colonizadas (31).
En este estudio se demostró que las rizobacterias de P. putida estudiadas estimulan el crecimiento de las plantas, con mejor comportamiento en la interacción planta-bacteria las cepas de P. putida AJ9, AJ14, AJ20 y AJ18.
Desde el punto de vista práctico, estos resultados tienen un gran valor, ya que se obtuvo un conjunto de cepas nativas de P. putida potencialmente eficientes en la promoción del crecimiento vegetal y el control biológico de P. grisea, que constituyen reservas para la producción de inoculantes microbianos en beneficio del cultivo del arroz, una vez que se realicen estos ensayos a escala de macetas y campo para confirmar que sus efectos son estables y repetibles en el tiempo.
REFERENCIAS
1. FAO. (2007). FAO production yearbook. In: FAO (Food and Agriculture Organization of the United Nations). FAO production yearbook. Roma, Italia. Disponible en: http://www.fao.org. (Consultado: 13 Feb 2007).
2. Álvarez AE, Zamora ZN, Jiménez EM. Comportamiento de variedades de arroz frente a Pyricularia grisea (Sacc.) en la provincia Granma. Rev Protección Veg. 2001;16(1):40-43.
3. Rives N, Hernández A, Acebo Y, Heydrich M. Caracterización de algunos géneros bacterianos asociados al cultivo del arroz variedad J-104. Revista Cubana del Arroz. 2006;2(3):7-13.
4. Acebo Y, Hernández A. Las técnicas moleculares para la identificación de bacterias de importancia agrícola, su contribución en la conservación del medio ambiente. Contribución a la Educación y la Protección del Medio Ambiente. (2004), Vol. 4. ISCTN. Soporte magnético: 34B-41B.
5. Hass D, Keel C. Regulation of antibiotic production in root-colonizing Pseudomonas spp. and relevance for biological control of plant disease. Ann Review of Phytopathology. 2003;41:117-153.
6. Hernández A. Obtención de un bioestimulador del crecimiento vegetal a partir de rizobacterias asociadas al cultivo del maíz (Zea mays L). Tesis para optar al grado de Doctor en Ciencias Biológicas. Instituto Nacional de Ciencias Agrícolas. 2002.
7. Schwyn B, Neilands JB. Universal Chemical Assay for the Detection and Determination of Siderophores. Analytical Biochemistry. 1987;160:47-56.
8. Bashan Y, Holguín G, Ferrera R. Interacciones entre plantas y microorganismos benéficos. Terra. 1996;14(2):159-192.
9. Harrigan WF, Mac Conce M. Métodos de Laboratorio en Microbiología. Editorial Academia. León. España. 1968; 1ra Edición. 426p.
10.Hernández A, Heydrich M, Acebo Y, Velázquez MG, Hernández AN. Antagonistic activity of Cuban native rhizobacteria against Fusarium verticillioides (Sacc.) Nirenb. in maize (Zea mays L.). Appl Soil Ecol. 2008;36:184-186.
11.Murashige T, Skoog F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plantarum. 1962;15:473-497.
12.Shapiro SS, Wilk MB. An analysis of Variance Test for Normality (Complete Samples). Biometrika. 1965;52:591-611.
13.Dayton CM, Schafer ND. Extended Tables of t and chi square for Bonferroni Test with unegual Error Allocation. JASA Asoc. 1973;68:78-83.
14.Trujillo ID. Caracterización de rizobacterias asociadas al cultivo del maíz (Zea mays L.) con actividad promotora del crecimiento vegetal. Tesis en opción al Titulo Académico de Maestro en Microbiología, Facultad de Biología. Universidad de La Habana; 2005.
15.Gravel V, Antoun H, Russell J, Weddell T. Growth stimulation and fruit yield improvement of greenhouse tomato plants by inoculation with Pseudomonas putida or Trichoderma atroviride: Possible role of indole acetic acid (IAA). Soil Biology & Biochemistry. 2007;39:1968-1977.
16.Velásquez M, Hernández A, Heydrich M, Hernández AN. Estudio de la interacción maíz-Burkholderia cepacia. Rev Latinoamericana de Microbiología. 1999;41:17-23.
17.Rojas M, Montoute M, Heydrich M. Potencialidades de cepas de Gluconacetobacter diazotrophicus aisladas de diferentes ecosistemas para la estimulación del crecimiento vegetal. En: Congreso Científico del INCA (7-10 nov 2006, La Habana). Memorias CD-ROM. Instituto Nacional de Ciencias Agrícolas
18.Buysens S, Heungens K, Poppe J, Hofte M. Involvement of pyochelin and pyoverdin in suppresion of Pythium induced damping off of tomato by Pseudomonas aeruginosa 7NSK2. Appl Environ Microbiol. 1996;62:865-871.
19.Ramamoorthy V, Viswanathan R, Raguchander T, Prakasam V, Samiyappan R. Induction of systemic resistance by plant growth promoting rhizobacteria in crop plants against pests and diseases. Crop Protection. 2001;20(11):1-11.
20.De Chial M, Ghysels B, Beatson SA, Geoffroy V, Meyer JM, Pattery T, et al. Identification of type II and type III pyoverdine receptors from Pseudomonas aeruginosa. Microbiology. 2003;149:821-831.
21.Joshi F, Archana G, Desai A. "Siderophore Cross-Utilization amongst Rhizospheric Bacteria and the Role of Their Differential Affinities for Fe3+ on Growth Stimulation under Iron-Limited Conditions. Current Microbiology. 2006;53:141-147.
22.Quintana E, Boado I. Biopreparado efectivo en la producción de cuatro cultivos de organopónico. En. Programas y Resúmenes XII Seminario Científico. Instituto Nacional de Ciencias Agrícolas, La Habana. 2000.
23.Sánchez L, Ortiz S, Hernández A. Evaluation of two culture media for producing siderophore from Burkholderia cepacia through stirred-tank fermentation. Rev Protección Veg. 2004;19:97-101.
24.Hernández A, Rives N, Hernández AN, Heydrich M. Characterization of the production of metabolites such as AIA siderophores and salycilic acid in rhizobacteria associated to maize crop. Rev Colombiana de Biotec. 2004;6(1):6-13.
25.Validov S, Mavrodi O, De La Fuente L, Boronin A, Weller D, Thomashow L, et al. Antagonistic activity among 2,4-diacetylphloroglucinol-producing fluorescent Pseudomonas spp. FEMS Microbiol Lett. 2005;242: 249-256.
26.González S, Rojas TI. Actividad antifúngica in vitro de dos productos residuales de la caña de azúcar. Contribución a la Educación y la Protección Ambiental. Cátedra de Medio Ambiente, 1999; Editorial Academia. V Taller. p. 91-93.
27.Surk-Sik M, Mo Kang P, Sup Yoon K, Se-Joon Y, Bin Park B. Synthesis of plant growth promoting and fungicidal 4 quinolinone metabolites of
Pseudomonas cepacia. Bull Korean Chem Soc. 1995;16:1128-1130.
28.Pujol M, Badosa E, Cabrefiga J, Montesinos E. Development of a strain-specific quantitative method for monitoring Pseudomonas fluorescens EPS62e, a novel biological control agent of fire blight. FEMS Microbiol Lett. 2005;249:343352.
29.Pazos, M. Aislamiento e identificación de cepas nativas, pertenecientes al género Azospirillum mediante técnicas moleculares. Tesis en opción al Grado Científico de Maestro en Ecología Microbiana. Facultad de Biología. Universidad de La Habana. 2000; 50p.
30.Egamberdiyeva D. The effect of plant growth promoting bacteria on growth and nutrient uptake of maize in two different soils. Appl Soil Ecol. 2007;36:184-189.
31.Patten CL, Glick R. Role of Pseudomonas putida indolacetic acid in development of the host plant root system. Appl Environ Microbiol. 2002;68;3795-3801.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}