










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión impresa ISSN 1010-2752versión On-line ISSN 2224-4697
Rev. Protección Veg. v.24 n.3 La Habana sep-dic. 2009
TRABAJO ORIGINAL
COMPORTAMIENTO DE GENOTIPOS DE LA FAMILIA SOLANACEAE FRENTE A Meloidogyne incognita (KOFOID Y WHITE) CHITWOOD1
BEHAVIOUR OF GENOTYPES FROM SOLANACEA FAMILY IN FRONT TO Meloidogyne incognita (KOFOID Y WHITE) CHITWOOD1
Mayra G. Rodríguez*, Lucila Gómez*, Farah M. González**, Yudines Carrillo***, Maite Piñón**, Olimpia Gómez**, A.S. Casanova**, Martha Álvarez****, Belkis Peteira*
*Grupo de Plagas Agrícolas, Dirección de Protección de Plantas, Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, La Habana, Cuba. **Instituto de Investigaciones Hortícolas "Liliana Dimitrova" (IIHLD). Quivicán, La Habana. Cuba. ***Facultad Agronomía, Universidad Agraria de La Habana, Cuba. ****Instituto Nacional de Ciencias Agrícolas (INCA), San José de las Lajas, Cuba
RESUMEN
Los nematodos formadores de agallas (Meloidogyne spp.) constituyen factores limitantes de los rendimientos en la producción de hortalizas en campo abierto y casas de cultivo, donde el tomate (Solanum lycopersicum L. (ex Lycopersicon esculentum Mill)) representa un buen hospedante de estos nematodos. Para manejar las poblaciones de estos organismos, entre otras tácticas se hace uso de genotipos resistentes o se acude al injerto herbáceo para obtener resultados productivos favorables, resultando necesaria la evaluación de los genotipos a emplear frente a poblaciones nativas, pues se sabe que la resistencia puede ser quebrada por factores tales como la virulencia de la población de Meloidogyne. El objetivo de este estudio fue determinar el comportamiento de líneas, híbridos y variedades de tomate ante una población nativa de M. incognita raza 2, como elemento básico del programa de injerto herbáceo que se desarrolla en Cuba para contar con alternativas al bromuro de metilo. Se inocularon 10 plantas por genotipo, con 1,5 huevos/juveniles.g de suelo-1 y se evaluaron a los 60 días, determinándose el Índice de Agallamiento (IA), Factor de Reproducción (FR) e índice de reproducción (IR). Los cultivares evaluados fueron Lycopersicon peruvianum acc. B6-140; Physalis ixocarpa acc. B6236; Lycopersicon pimpinelifolium acc. Mex 121-A; las variedades/híbridos de S. lycopersicum var. cerasiforme, Vyta, Mariela, Amalia, HC 3880 y HA 3019; el híbrido inter-específico Lycopersicon hirsutum x L. esculentum, Solanum torvum y los híbridos de tomate T-1x14, T-1x15, T-2x16 y LTM. Resultó inmune L. hirsutum x L. esculentum y muy resistentes S. torvum y L. peruvianum. Por su parte, P. ixocarpa se comportó como moderadamente resistente y el resto de los materiales evaluados fueron susceptibles a la población cubana de M. incognita raza 2.
Palabras clave: Meloidogyne incognita; resistencia; susceptibilidad; Solanaceae; injerto
ABSTRACT
The root knot nematodes (Meloidogyne spp.) are limiting factors for yields of vegetables in open areas and crops under shelter conditions, where tomato Solanum lycopersicon L. (ex Lycopersicon esculentum Mill.) represents a very good host for these nematodes. For population management of this pest, resistance and graffing are used, among other tactics, to obtain good productive results, but it is necessary to evaluated the resistance of genotypes to native populations, since as it is know resistancemay be broken by factors like virulence of Meloidogyne population. The objective of this study was to determine the behaviour of tomato lines hybrids and varieties to native population of M. incognita race 2, as a basic element for the graffting program being develop in Cuba as an alternative to methyl bromide. Ten plants per genotype were inoculated with 1,5 eggs/juvenils.g of soil-1 and evaluated after 60 days to determined the Root Galling Index (IA), the Reproduction Factor (FR) and the Reproduction Index (IR). The cultivars evaluated were Lycopersicon peruvianum acc. B6-140; Physalis ixocarpa acc. B6236; Lycopersicon pimpinellifolium acc. Mex 121-A; the following varieties/hybrids of Solanum lycopersicum var. ceraciforme, Vyta, Mariela, Amalia, HC 3880 and HA 3019; the interespecific hybrid Lycopersicon hirsutum x L. esculentum, Solanum torvum and the tomato hybrids T-1x14, T-1x15, T-2x16 and LTM. The interespecific hybrid L. hirsutum x L. esculentum was immune, whereas S. torvum and L. peruvianum were very resistant. In other hand, P. ixocarpa behaved as moderately resistant and the rest of genotypes were susceptibles to the Cuban population of M. incognita race 2.
Key words: Meloidogyne incognita; resistance; susceptibility; Solanaceae; grafting
INTRODUCCIÓN
Para Cuba, la producción protegida y semi-protegida de hortalizas constituye uno de los programas priorizados del Ministerio de la Agricultura (MINAG) (Pérez, comunicación personal2), de ahí que el desarrollo de estos sistemas ha recibido un impulso en los últimos años, con la incorporación de decenas de hectáreas y la creación de miles de puestos de trabajo que abarcan la cadena productiva agro-industrial.
En estas instalaciones se producen fundamentalmente cultivos como tomate (mayor proporción), pimiento, pepino y melón (1), donde numerosos factores inciden negativamente en los rendimientos, entre ellos se destacan los nematodos fitoparásitos del género Meloidogyne (2, 3), los que, durante algunos años, constituyeron una de las causas fundamentales del uso de bromuro de metilo en las instalaciones.
Teniendo en cuenta el programa que desarrolla Cuba en la sustitución de este químico dañino a la capa de Ozono, el MINAG viene financiando investigaciones básicas y acciones de innovación tecnológica desde hace algunos años, con el objetivo de dotar a los productores de alternativas conducentes a lograr el manejo de plagas como los nematodos formadores de agallas (Meloidogyne spp.), sin bromuro de metilo. Estos estudios se han desarrollado en vario proyectos, como el ejecutado entre los años 2003 y 2006 por un equipo de investigación de más de 15 especialistas pertenecientes a cinco instituciones (2). En dicho proyecto, se evaluaron alternativas como la biofumigación (4), uso de plantas trampa (5), agentes de control biológico (2) y la resistencia de genotipos de tomate y otras especies frente a Meloidogyne spp., entre otras.
Según Bridge (6), el uso de cultivares resistentes podría ser la forma más útil y barata de controlar nematodos para campesinos a pequeña escala, lo que se puede aplicar también a la agricultura de altos insumos como la producción protegida de hortalizas. Este autor planteó que, el injerto podría ser utilizado para el control de plagas de suelo, cuando no hay cultivares resistentes disponibles en el cultivo que se pretende desarrollar. El uso del injerto herbáceo constituye una de las opciones de amplio uso en la producción protegida de hortalizas, como alternativa al uso de bromuro de metilo (7, 8). Con relación a esto, debe tenerse presente que, la interrelación de los nematodos fitoparasíticos y sus plantas hospedantes es compleja (9) y por tanto se hace imprescindible su evaluación cuidadosa para poder determinar su comportamiento ante una población dada.
El objetivo de este estudio fue evaluar la resistencia de diferentes genotipos de la familia Solanaceae a una población nativa Meloidogyne incognita (Kofoid y White) Chitwood, para su uso como porta-injertos y/o su empleo comercial en los sistemas de cultivos protegidos de hortalizas.
MATERIALES Y MÉTODOS
Para el estudio se seleccionó una población de M. incognita raza 2, proveniente de una casa de cultivo enclavada en el municipio de San José de las Lajas, provincia La Habana en el año 2001 y mantenida en el banco de poblaciones puras del CENSA. La selección de la especie de nematodo estuvo determinada por el hecho de que constituye la de mayor distribución en instalaciones de producción protegida en el país. La especie seleccionada fue previamente identificada a través de las técnicas morfológicas y moleculares (3). Para su multiplicación y mantenimiento se utilizaron cajuelas de zinc galvanizado de 50 x 50cm con una mezcla de suelo y material orgánico (proporción 1:1), esterilizada en autoclave (121°C, 30 min.), donde crecían plantas de berenjena (Solanum melongena L.), madama (Impatiens balsamina L.) y tomate (S. lycopersicum var. Campbell 28).
Los experimentos se realizaron en condiciones semi-controladas, en los aisladores biológicos del CENSA, con el uso de macetas de 1,5L de capacidad (»1,5 kg), donde se colocó una mezcla de suelo y material orgánico, con características similares a las descritas anteriormente.
Las semillas de cada genotipo se colocaron en bandejas de polietileno con alvéolos (cepellones) contentivas de una mezcla de suelo y materia orgánica. A los 21 días de haber germinado, las plántulas se transfirieron a las macetas.
Evaluación de genotipos para ser utilizados como porta-injertos de hortalizas: Los materiales evaluados con el objetivo de conocer su comportamiento frente a M. incognita fueron los siguientes: Lycopersicon peruvianum L. acc. B6-140; Physalis ixocarpa Brot acc. B6236; Lycopersicon pimpinelifolium Dunal (tomate cimarrón) acc. Mex 121-A; S. lycopersicum var. cerasiforme; el híbrido interespecífico Lycopersicon hirsutum Humb. & Bonpl. x L. esculentum y Solanum torvum Sw. (pendejera). Las semillas de estos cultivares fueron suministradas por el Proyecto de Injerto, del Programa Ramal de Hortalizas que lidera el Instituto de Investigaciones Hortícolas «Liliana Dimitrova» (IIHLD).
De igual forma se evaluaron tres cultivares de tomate, que de mostrar resistencia, serían candidatos potenciales para el programa de injerto, por las posibilidades de contar con semillas de procedencia nacional. Los cultivares de tomate fueron: HC 3880; Vyta (suministradas por el IIHLD); Amalia y Mariela, obtenidas por el Instituto Nacional de Ciencias Agrícolas (INCA).
Evaluación de híbridos para la producción en casas de cultivo: Se realizó la evaluación de cuatro híbridos de tomate, provenientes del programa de mejoramiento del IIHLD para ser empleados en casas de cultivo por sus potencialidades en cuanto a resistencia a patógenos. Los híbridos aparecen registrados con los siguientes códigos. T-1x14, T-1x15, T-2x16 y LTM. Adicionalmente, se evaluó el cultivar de tomate HA 3019 (Firma Hazera), que se utiliza generalmente en el país en la producción protegida, que no había sido evaluada en las condiciones de Cuba frente a M. incognita.
En ambos ensayos, las plantas se inocularon una semana después del trasplante. El inóculo se preparó siguiendo la metodología de Hussey y Barker (10) y se introdujo en el suelo a través de cuatro orificios practicados sobre el sistema radical cercano al tallo en cada maceta, por el vertimiento de una suspensión de 1,5 huevos _ J2.g de suelo-1 de M. incognita.
Las macetas se dispusieron siguiendo un diseño completamente aleatorizado, donde los genotipos inoculados constituyeron los tratamientos, con 10 repeticiones cada uno, utilizando como control susceptible igual cantidad de plantas de Cucurbita moschata var. RG5. Las plantas se mantuvieron con riego en días alternos y se ejecutaron evaluaciones semanales del estado sanitario de los experimentos, de modo que si aparecían plagas (hongos, artrópodos, otros) tomar las medidas pertinentes.
A los 60 días de la inoculación, se determinó el Índice de Agallamiento (IA) mediante la escala de Taylor y Sasser, (11) y se extrajeron de las raíces los huevos y J2, a través del método de Hussey y Barker (10), para determinar la población final de nematodos. La cuantificación de la suspensión resultante se ejecutó a través del conteo directo de los J2- huevos en un microscopio estereoscopio Zeiss® con 160 aumentos, lo que determinó el Factor de reproducción (FR) (11).
El FR, que establece la cantidad de veces que se reprodujo la población inicial, se calculó a través de la formula: FR = Pf / Pi; donde Pf constituye la población final extraída de las raíces después del experimento y Pi la cantidad de huevos-larvas inoculadas a las plantas.
La categorización de los hospedantes se hizo a través de la metodología de Sánchez et al. (inédito3) y los detalles se recogen en la Tabla 1. Para ello se determinó el Índice de Reproducción a través de la siguiente formula: Índice de Reproducción (IR)= (Pvar. X 100)/P testigo, donde, P. var = número de huevos-J2 producido por la variedad vegetal que se evalúa y P testigo: número de huevos-J2 producido por la variedad vegetal que se empleó como testigo susceptible.
Con los datos de IA, FR e IR de los genotipos que se evaluaron para ser utilizados como porta-injertos se realizó un análisis de conglomerado utilizando la distancia de Manhattan y el Método de Ward, del paquete estadístico InfoStat (12).
RESULTADOS Y DISCUSIÓN
Evaluación de genotipos para ser utilizados como porta-injertos de hortalizas:
Se evidenció que Lycopersicon hirsutum x L. esculentum se comportó como inmune, con IA e IR=0 (Tabla 2), mientras que S. torvum y L. peruvianum se comportaron como muy resistentes. P. ixocarpa permitió un nivel de reproducción del nematodo M. incognita que lo ubica como moderadamente resistente, mientras que el resto de los genotipos evaluados se comportaron como muy susceptibles a la población cubana de M. incognita raza 2.
Con relación al primer genotipo (L. hirsutum x L. esculentum), se plantea que es una de las más diversas y ricas fuentes de genes de resistencia a plagas y enfermedades que atacan al tomate cultivado (13; 14). En este sentido, Arie et al. (14) señalaron que por ejemplo, de las 16 plagas y 32 patógenos que afectan el tomate, L. hirsutum presenta resistencia a 14 y 9 respectivamente. En su estudio, utilizando recientes avances de la biología molecular y de los estudios de interacción planta-patógeno, Zang et al. (13) lograron construir un mapa de vínculo molecular basado en el cruce de L. esculentum × L. hirsutum e identificaron 29 marcadores RGA (genes análogos de resistencia), lo que podría ofrecer avances en los estudios con esta especie en un futuro cercano. Este genotipo debe continuar en el programa de estudio para la tecnología de injertos de hortalizas, siendo necesaria su evaluación ante poblaciones cubanas de las especies de Meloidogyne que han sido encontradas en los polos de cultivos protegidos del país.
Con relación a S. torvun, su reacción en este estudio coincide con lo mostrado por este genotipo en investigaciones desarrolladas en el ámbito internacional. Respecto a la resistencia de esta especie, Daunay y Dalmasso (1985) citados por Gómez (3), informaron que S. torvum se comportó como resistente a M. incognita y M. arenaria, comportándose como pobre hospedante de M. javanica.
Con relación a las investigaciones que deben ser ejecutadas para lograr éxito en el programa de injerto herbáceo, se sabe que resulta necesario desarrollar evaluaciones relacionadas con la resistencia del porta-injerto a los nematodos, como las efectuadas en este estudio, así como la evaluación de la compatibilidad de estos materiales con el genotipo injertado, estudios que se ejecutan en la actualidad por un equipo de investigación del IIHLD y CENSA, liderado por la primera institución (15).
Al respecto, Rahman et al. (16), condujeron un estudio con el objetivo de determinar la resistencia de patrones de especies de Solanum a nematodos formadores de agallas para su uso como porta-injertos de variedades cultivadas de berenjena, evaluándose también la compatibilidad entre patrón y variedad. Los patrones S. torvum y Solanum sisymbriifolium mostraron reacción resistente frente a M. incognita. De igual forma, estos autores evaluaron catorce genotipos frente a M. incognita y entre ellas, solo seis mostraron resistencia. Cuando tres de esos materiales (Sufala, Singnath y Kazla), fueron injertados a su vez sobre S. torvum y S. sisymbriifolium, se obtuvo alto porcentaje de éxito (95%) en el caso de S. torvum con Sufala y el menor (85%) en S. sisymbriifolium con Singnath. Las plantas injertadas mostraron resistencia en condiciones semi-controladas y en campo, así como mayores rendimientos que las plantas no injertadas. La combinación S. torvum con Sufala tuvo los mayores rendimientos.
Morra et al. (17), en experimentos en casas de cultivo, donde se empleó berenjena (Solanum melongena L.) cultivar Galine, injertada sobre los patrones S. torvum o S. integrifolium o creciendo sobre sus propias raíces, encontraron que los rendimientos fueron mayores en las plantas injertadas sobre S. torvum. Las plantas fructificaron durante un periodo de tiempo mayor y fueron menos infestadas por M. incognita. Por dichos resultados, S. torvum ha sido utilizada exitosamente como porta-injerto para berenjena a escala internacional y los resultados de este
estudio, avalan su inclusión en el programa de estudios para lograr la eficacia del injerto herbáceo en Cuba.
Con relación a la resistencia de L. peruvianum, Jacquet et al. (18) plantearon en una excelente revisión que, la resistencia a Meloidogyne fue originalmente observada en algunas accesiones de plantas de tomate silvestre de la especie L. peruvianum por Bailey en 1941 y que subsecuentemente se demostró que se debía a un gen simple dominante que llamaron Mi. Además expresaron estos autores que, en los estudios posteriores llevados cabo por Barham y Winstead en la década de los años 50, se demostró que este gen era efectivo o controlaba a tres especies mayores.
Con relación a la transferencia de la resistencia a partir de L. peruvianum hacia L. esculentum, existe alguna controversia con respecto a los autores. Por una parte, Jacquet et al. (18) señalan que el gen Mi fue transferido de L. peruvianun PI12856 a L. esculentum por "rescate de embriones" (rescue embryo) por Smith en 1944 y por otra parte, Cook y Evans (19) aseveran que dicha resistencia a L. esculentum fue transferida desde L. peruvianum por Frazier y Dennett en 1949. En el elemento que si coincide la literatura es que, todas las variedades actuales que exhiben resistencia a Meloidogyne, son derivadas de cruzamientos en los cuales, el gen Mi fue incorporado desde L. peruvianum (20).
En Cuba, en un estudio previo, Cuadra et al. (21) informaron que L. peruvianum se comportó como altamente resistente ante una población cubana de M. incognita raza 2, donde no se produjo agallamiento (IA= 0). Sin embargo, en nuestro trabajo, la accesión de L. peruvianum evaluada mostró agallamiento y hubo reproducción del nematodo, aunque baja. Esto puede deberse a dos factores, en primer lugar a las diferencias en los métodos de inoculación empleados en ambos estudios, donde el uso de raíces agalladas (21) vs. inóculo de J2/huevos en nuestro estudio, puede inducir diferencias. En segundo lugar, el uso de diferentes poblaciones de M. incognita.
El hecho de que la población empleada en este estudio sea capaz de parasitar y tener algún nivel de reproducción en una planta como L. peruvianum (gen Mi), indica que se trata de una población que exhibe lo que algunos autores denominan virulencia parcial (22), ya que según Roberts (23), la virulencia en Meloidogyne por simple definición, se refiere a poblaciones que se reproducen significativamente en plantas hospedantes que previenen o suprimen (portadoras del gen Mi) la reproducción de poblaciones avirulentas.
Con relación a la efectividad del gen Mi para suprimir la reproducción de Meloidogyne spp., se plantea que, aunque es altamente eficiente, en muchos casos, el uso intensivo de genotipos portadores de este gen sin otras medidas de manejo, como no sea el empleo de la resistencia, ha mostrado desaciertos, lo que, unido a la variabilidad patogénica de Meloidogyne spp., ha provocado preocupación acerca de la durabilidad de dicha resistencia.
A pesar de las diferencias encontradas entre los dos estudios ejecutados en Cuba, lo cierto es que L. peruvianum podría constituirse, a partir de los resultados de ambas investigaciones, en una opción favorable a ser incluido en el programa de injerto de hortalizas, teniendo siempre en cuenta que el uso de genotipos resistentes, ya sea como porta-injertos o para su explotación comercial, deben ser parte de programas de Manejo Integrado de Plagas (MIP), donde la utilización de diversas tácticas, entre ellas la rotación de cultivos, permitan mantener las poblaciones bajas y se evite que aparezcan poblaciones virulentas.
Con relación al comportamiento de P. ixocarpa frente a nematodos, resulta muy escasa la información en las fuentes consultadas, encontrándose que Peña-Lomeli et al. (24), no señala a los nematodos formadores de agallas como plaga de este cultivo, lo que indica que probablemente no sea buen hospedante para Meloidogyne.
Por su parte, L. pimpinellifolium se comportó como muy susceptible, resultado que coincide con lo informado por Cuadra et al. (21) acerca de que las accesiones evaluadas resultaron susceptibles. Según Arie et al. (14), esta especie ha sido fuente de genes de resistencia para L. esculentum a los patógenos fungosos y bacterianos. Sin embargo, en su revisión del tema no mencionan que posea resistencia a Meloidogyne. Teniendo en cuenta nuestros resultados, consideramos que este genotipo no debe ser empleado en el programa de injerto herbáceo para el manejo de M. incognita.
De igual manera sucede con los cultivares Vyta, Amalia, Mariela y el híbrido HC 3880, los que se comportaron como muy susceptibles. Con relación al último genotipo, los resultados que se muestran coinciden con los obtenidos anteriormente (21).
El análisis de conglomerado, divide a los genotipos en dos grupos con más de 95% de disimilitud (Fig. 1), en uno donde se ubican los genotipos susceptibles y en el otro donde lo hacen los que exhiben diferentes grados de resistencia y los genotipos L. pimpinelifolium y S. lycopersicum var. ceraciforme, los que, aun cuando resultaron susceptibles, exhiben menores valores de IR, FR y número total de juveniles y huevos por sistema radical que las variedades nacionales de tomate (Vyta, Amalia y Mariela) y el híbrido HC 3880.
El análisis realizado corroboró las categorías asignadas a los genotipos con el uso de esta metodología, la que resulta útil en la evaluación de especies de plantas frente a nematodos formadores de agallas.
Evaluación de cultivares para ser utilizados en la producción en casas de cultivo:
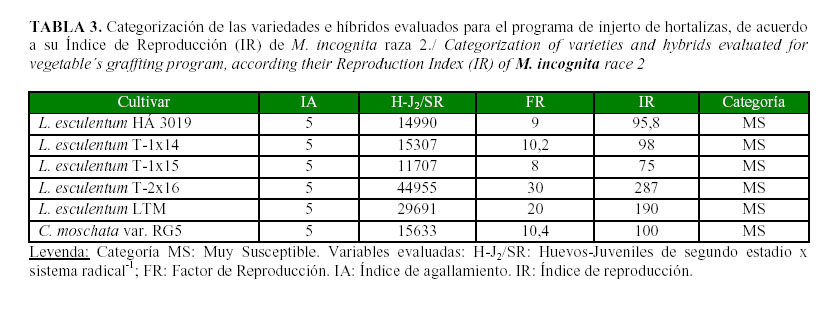
Los resultados de la evaluación de los híbridos para ser utilizados en la producción, se presentan en la Tabla 3, pudiéndose constatar que todos se comportaron como muy susceptibles a M. incognita. Ellos permitieron libremente la reproducción del nematodo, cuya población creció en las raíces entre 8 y 30 veces, con relación al inóculo inicial, con números de huevos y J2 superiores a los 10 000 por sistema radical. Esto sugiere que la plantación/siembra de estas variedades e híbridos en suelos con un nivel de 1,5 huevos-juveniles de M. incognita raza 2 por gramo de suelo provocará una elevación sustancial de la misma en el suelo, lo que comprometería el desarrollo óptimo de esa cosecha y las siguientes.
Los genotipos T-1x14, T-1x15; T-2x16 y LTM constituyen nuevos híbridos que están siendo estudiados en el programa de mejoramiento genético liderado por el IIHLD, debido a que poseen genes de resistencia a begomovirus (25), sin embargo, no se habían enfrentado a poblaciones de Meloidogyne spp.
De forma general, con relación a los cultivares de tomate, la reacción frente a Meloidogyne spp. es diversa, según consta en la amplia información a la que se puede acceder, y las reacciones van desde la susceptibilidad de cultivares como Campbell-28 a la resistencia de Rossol (21, 26).
Resulta importante resaltar el hecho de que el híbrido HA3019, uno de los más empleados en nuestro país (Tabla 3), permita una alta reproducción de M. incognita, especie más distribuida en estos sistemas de producción (3), lo que sugiere que su uso en las instalaciones, sin ejecutar acciones adecuadas de manejo de nematodos, contribuirá sustancialmente a la elevación de las poblaciones de esta plaga lo que conllevará al deterioro de la plantación y a que no se completen los ciclos de seis meses que se esperan con el uso de esta tecnología. Teniendo en cuenta estos resultados y que dicho híbrido continuará siendo utilizado por los productores en la producción protegida de hortalizas, resulta entonces imprescindible el uso de tácticas que permitan mantener las poblaciones en los niveles más bajos que sea posible y entre estas, las prácticas culturales como la biofumigación, tendrán un papel importante.
Es necesario sugerir que la variedad Rossol, la que ha manifestado resistencia en las condiciones de Cuba (21, 26) sea considerada por el equipo de trabajo del IIHLD para su inclusión en el programa de estudio.
La importancia y aplicabilidad de los resultados obtenidos, dan la medida de la necesidad de continuar la evaluación del germoplasma disponible para el programa de injerto que se desarrolla en Cuba como parte de las alternativas que se evalúan al uso del bromuro de metilo.
AGRADECIMIENTOS
Los autores desean expresar su agradecimiento a los técnicos Roberto Enrique y Lidia López del CENSA, por su valiosa colaboración en la realización de este estudio. A la Dra. Ileana Miranda por llevar a cabo los análisis estadísticos y al Ing. A. Martell (MINAG San José de las Lajas), por el suministro de semillas del híbrido HA 3019.
REFERENCIAS.
1. Casanova AS, Gómez O, Pupo FR, Hernández M, Chailloux M, Depestre T, et al. Manual para la Producción Protegida de Hortalizas. MINAG-Viceministerio de Cultivos Varios-IIHLD, La Habana, Cuba. 2007. 110p.
2. Rodríguez mg, Gómez L, Cuadra R, Díaz-Viruliche L., Fernández E, Casanova A, et al. Nematodos formadores de agallas en Sistemas de Cultivos Protegidos: Diagnóstico y Manejo. Informe Final de Proyecto. Programa ramal de Hortalizas - MINAG. (Inédito, copias depositadas en Laboratorio de Nematología CENSA y Programa Ramal Hortalizas-MINAG, Cuba). 2006. 171p.
3. Gómez L. Diagnóstico de nematodos agalleros y prácticas agronómicas para el manejo de Meloidogyne incognita en la Producción Protegida de Hortalizas. Tesis para optar por el Grado Científico de Doctor en Ciencias Agrícolas. Universidad Agraria de La Habana- Centro Nacional de Sanidad Agropecuaria. 2007. 100 p .
4. Gómez L, Rodríguez MG, Díaz-Viruliche L, González E, Wagner F. Evaluación de materiales orgánicos para la biofumigación en instalaciones de cultivos protegidos para el manejo de Meloidogyne incognita. Rev Protección Veg. 2005;21(3):178-185.
5. Cuadra R, Cruz X, Fajardo JL, Ortega J, Fernández E, Rodríguez MG, et al. Los cultivos de ciclo corto como planta extractora de nematodos de las agallas en un sistema de manejo integrado. Ponencia Forum Municipal de Ciencia y Técnica, Boyeros. Ciudad de La Habana. (INIFAT- Documento inédito). 2005. 15 p.
6. Bridge J. nematode management in sustainable and subsistence agriculture. Ann Rev Phytopathol. 1996;34:201-225.
7. Miguel A. Contribución de la técnica del injerto en hortalizas para la eliminación del bromuro de metilo. En: V Seminario Científico Internacional de Sanidad Vegetal. 24-28 Mayo. Ciudad de La Habana, Cuba. 2004. CD Memorias ISBN 959-26-137-6.
8. Rodríguez mg, Gómez L, Díaz-Viruliche L. Alternativas para la sustitución del Bromuro de Metilo en el manejo de nematodos formadores de agallas (Meloidogyne spp.). (Revisión de literatura 1995-2006). Documento contenido en CD "Primer Curso Internacional de Introducción a la Nematología Agrícola". Marzo, 2006. Editado por Instituto Nacional de Investigaciones Agrícolas (INIA), Maracay. Venezuela.
9. Hussey RS, Williamson VM. Physiological and molecular aspects of nematode Parasitism. En: Barker K, Pederson G, Windham G, editors. Plant and Nematode Interactions. Agronomy Monograph No 36. Madison, Wisconsin, USA. 1998:87-108.
10.Hussey RS, Barker KB. A comparison of methods of colleting inocula of Meloidogyne spp. including a new technique. Plant Dis Report. 1973;57:1025-1028.
11.Taylor AL, Sasser JB. Biology, identification and control of root-knot nematodes (Meloidogyne species). Dept. Pl. Pathol. N.C. State Univ., Raleigh; 1978. 111p.
12.Di Riezo JA, Balzamini M, Casanoves F, González L, Tablada M, Robledo CW. Grupo Infostat. Universidad Nacional de Córdoba, Argentina. Infostat Version 2009.
13.Zhang LP, Khan A, Niño-Liu D, Foolad MR. A molecular linkage map of tomato displaying chromosomal locations of resistance gene analogs based on a Lycopersicon esculentum × Lycopersicon hirsutum cross. Genome. 2002;45:133-146.
14.Arie T, Takahashi H, Kodama M, Teraoka T. Tomato as a model plant for plant-pathogen interactions. Plant Biotechnology. 2007;24:135-147.
15.González FM, Hernández A, Casanova AS, Depestre T, Gómez L, Rodríguez MG. El injerto herbáceo; alternativa para el manejo de plagas de suelo. Rev Protección Veg. 2008;23(2):69-74.
16.Rahman MA, Rashid MA, Salam MA, Masud MA, Masum AS, Hussain MM. Performance of Some Grafted Eggplant Genotypes on Wild Solanum Root Stocks against Root-Knot Nematode. J Biological Sciences. 2002;2(7):446-448.
17.Morra L, Troisi A, Bilotto M, D'Amore R. Grafted aubergines, agronomic effects and crop protection. Colture Protette. 2000;29(1):33-38.
18.Jacquet M, Bongiovanni M, Martinez M, Verschave P, Wajnberg E, Castagnone-Sereno P. Variation in resistance to the root-knot nematode Meloidogyne incognita in tomato genotypes bearing the Mi gene. Plant Pathology. 2005;54:93-99.
19.Cook R, Evans K. Resistance and tolerance. En: Brown RH, Kerry B. (Eds.) Principles and practice of nematode control in crops. Academic Press; 1987;79-232.
20.Williamson VM. Root Knot Nematode resistance genes in tomato and their potential for future use. Ann Rev Phytopathol. 1998;36:277_293.
21.Cuadra R, Cruz X, Ortega J, Shagarodsky T, González M. Respuesta de Lycopersicon spp. frente al ataque del nematodo de las agallas (Meloidogyne incognita). Rev Protección Veg. 2005;20(2):114-121.
22.Tzortzakakis EA, Trudgill DL, Phillips MS. Evidence for a dosage effect of the Mi gene on partially virulent isolates of Meloidogyne javanica. J Nematology. 1998;30:76-80.
23.Roberts PA. Conceptual and practical aspects of variability in root-knot nematodes related to host plant resistance. Ann Rev Phytopathol. 1995;33:199-221.
24.Peña-Lomeli A, Molina-Galán JD, Cervantes-Santana T, Márquez-Sánchez F, Sahagún- Castellanos J, Ortíz-Cereceres J. Heterosis invernal en tomate de cáscara (Physalis ixocarpa Brot). Rev Chapingo. 2001;4(1):31-39.
25.Gómez O, Rodríguez G. Impacto del cultivar en la producción protegida de hortalizas. Conferencia contenida en CD del Curso "Manejo Integrado de Plagas en la Producción Protegida de Hortalizas", 14 al 28 de septiembre 2007. Centro Nacional de Sanidad Agropecuaria, Cuba. 26p.
26.Fernández E, Pérez M, Gandarilla H, Vázquez R, Fernández M, Paneque M, et al. Guía para disminuir infestaciones de Meloidogyne spp., mediante el empleo de cultivos no susceptibles. Boletín Técnico, Sanidad Vegetal. 1998;4(4):1-18.
1Estudio desarrollado con financiamiento de los proyectos "Nematodos formadores de agallas en sistemas de cultivos protegidos: Diagnóstico y manejo" (Programa Ramal: Hortalizas) y "Perfeccionamiento de la Agricultura con base Agroecológica en San José de las Lajas" (Programa Nacional de Ciencia y Técnica: Seguridad Alimentaria) ambos del Ministerio de la Agricultura (MINAG), República de Cuba.
2 Dra. María del Carmen Pérez, Ministra de la Agricultura en funciones. Conferencia ofrecida por la Ministra al Pleno de la Academia de Ciencias de Cuba. Instituto de Ciencia Animal, La Habana. mayo de 2007.
3Sánchez L, MG. Rodríguez, I Rodríguez, R Enrique. (1992): Metodología para evaluar la resistencia en cultivos agrícolas a nematodos de agalla (Meloidogyne spp.). (basada en los elementos referidos por Triantaphyllou, 1975; Hadisoeganda y Sasser, 1982 y otros autores, incorporando la categoría de plantas inmunes). Laboratorio de Nematología. CENSA. 12pp (inédito)
(Recibido 29-5-2008; Aceptado 30-9-2009)


{kind=link}
{kind=link}
{kind=link}