










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión On-line ISSN 2224-4697
Rev. Protección Veg. v.24 n.3 La Habana sep-dic. 2009
VCP1 PROTEASE DETECTION IN Pochonia chlamydosporia var. CATENULATA STRAIN IMI SD 187
DETECCCIÓN DE PROTEASA VCP1 EN LA CEPA IMI SD 187 DE Pochonia chlamydosporia var. CATENULATA
B. Peteira-Delgado*, L. Hidalgo-Díaz**
* Grupo de Fitopatología, **Grupo Plagas Agrícolas, Dirección de Protección de Plantas, Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, La Habana, Cuba. Correo electrónico: bpeteira@censa.edu.cu
ABSTRACT
The mode of action of Pochonia as a biological control agent is the penetration by means of the hyphae into the target nematode eggs. The egg shell of nematodes has an outer vitelline membrane composed mainly by proteins. Thus, hyphal penetration is the result of a physic pressure and the specific hydrolytic activity of some enzymes such as proteases and chitinases. The most important protease studied in Pochonia chlamydosporia var. chlamydosporia is VCP1. The aim of this work was to detect and characterize the protease VCP1 in a strain of P. chlamydosporia var. catenulata, a potential biological control agent for root-knot nematodes. The specific activity of this protease was tested on different culture media supplemented with protein inductors using N-succinyl-Ala-Ala-Phe-p-nitroanilide as substrate. PCR and RFLP analyses with specific primers and probes were also done. Isolates from the chlamydosporia variety were used as positive controls. The medium supplemented with chitin was the only one capable of inducing some VCP1 activity but in a low level. The VCP1-encoding gene was not detected in strain IMI SD 187 by RFLP even when the stringency conditions were reduced to the minimum possible. The low level of enzymatic activity detected by biochemical techniques does not support the presence of protease VCP1 in this strain. The low enzymatic activity reached points out to the existence of a VCP1 isoform or another different protease. These hypotheses should be proved widening the range of catenulata variety strains and taking the chlamydosporia variety (strain 10) as a reference in a comparative study with conclusive results. The results of this enzymatic characterization related to the egg infection process of host nematodes are the first achieved in a catenulata variety strain in the world.
Key words: nematode parasites; RFLP; enzymes; protease
RESUMEN
El modo de acción de Pochonia como agente de control biológico es a través de la penetración de las hifas en los huevos de sus nematodos diana. La cubierta de los huevos de los nematodos posee una capa externa vitelina, compuesta en su mayoría por proteínas. Por tanto, la penetración es el resultado de una presión física y de la actividad hidrolítica específica de algunas enzimas tales como proteasas y quitinasas. La proteasa más importante del género Pochonia ha sido estudiada en la variedad chlamydosporia y es conocida como VCP1. El objetivo de este trabajo fue la detección y el estudio del comportamiento de la proteasa VCP1 en una cepa promisoria de Pochonia chlamydosporia var. catenulata. La actividad enzimática de esta proteasa se estudió en diferentes medios suplementados con inductores, empleando como sustrato N-succinil-Ala-Ala-Phe-p-nitroanilida. También se desarrollaron análisis PCR y RFLP con cebadores y sondas específicos. En todos los experimentos, los controles positivos utilizados fueron aislamientos de la variedad chlamydosporia. El medio suplementado con quitina fue el único capaz de inducir alguna actividad de la VCP1 pero en un nivel bajo. El gen que codifica para la enzima VCP1 no fue detectado por RFLP en la cepa IMI SD 187, aún cuando las condiciones de la hibridación de la sonda fueron reducidas al mínimo posible. El bajo nivel alcanzado de la actividad de esta enzima apunta hacia la existencia de una isoforma de la VCP1 u otra proteasa totalmente diferente. Esta hipótesis debe ser comprobada aumentando el rango de cepas de la variedad catenulata, teniendo como referencia la cepa 10 de la variedad chlamydosporia en un estudio comparativo con resultados conclusivos en este aspecto. Los resultados de esta caracterización enzimática relacionada con el proceso de infección de los huevos del nematodo hospedante son los primeros que se desarrollan en una cepa de la variedad catenulata, en el mundo.
Palabras clave: nematodos parásitos; RFLP; enzimas; proteasas
INTRODUCTION
Pochonia chlamydosporia is an opportunistic fungal parasite of eggs of sedentary nematodes including Meloidogyne spp. (1). Due to differences in the conidial disposition on phialides there is some controversy on whether Pochonia chlamydosporia var. chlamydos-poria and P. chlamydosporia var. catenulata are separated species or different varieties. Conidia are disposed as chains in the catenulata variety or as false heads in the chlamydosporia variety. Gams (2) considered that such differences had a limited importance and proposed to consider chlamydosporia and catenulata as varieties of the same species, Verticillium chlamydosporium. This judgement was sustained by Gams and Zare (3) when they renamed this species in the Pochonia genus.
The design of primers based on a nucleotidic sequence of b-tubulin gene intron from P. chlamydosporia has allowed the in situ detection of the chlamydosporia variety in tomato plants infected with root-knot nematodes with high specificity (4). At present, there are specific primers for the detection of the catenulata variety in soil samples by conventional PCR, as well as by real time PCR (5,6). Moreover, it has been possible to differentiate both varieties of the fungus and the biotype A of catenulata variety by RFLP analyses on the Internal transcribed spacer (ITS) amplified products (1,7).
The mode of action of Pochonia as a biological control agent is the penetration of the nematode egg shell by the hyphae (8). The nematode egg shell has an outer vitelline membrane composed mainly by proteins. Thus, fungal penetration is the result of a physic pressure and specific hydrolytic activity of some enzymes such as proteases and chitinases (9). For these reasons, the study of extracellular enzymes and other metabolites excreted by the fungus is an attractive issue for many researchers (10, 11, 12) due to their possible role as virulence factors or as a criterium for strain selection. A serine protease similar to subtilisin was purified and partially characterized from P. chlamydosporia var. chlamydosporia, and this protein is known as VCP1 (11).
The VCP1 variability among isolates of P. chlamydosporia can be due to differences in the subtilisins and they can explain the different niches occupied by this facultative parasite in the soil, its host range as well as the virulence factor (11, 13). Recently, these differences were shown with the sequencing of the VCP1-encoding gene (12). Up to date, all the studies have been carried out with isolates from the chlamydosporia variety, using the strain 10 from Rothamsted collection as a reference.
Other studies at DNA or protein level contributing to the intrinsic characterization of both varieties of the fungus, their behaviour during the infection process and the interaction between both, may provide important insights that may lead to a better distancing and taxonomic classification.
The objective of this work was to detect and characterize the protease VCP1 in a strain of Pochonia chlamydosporia var. catenulata growing in different media supplemented with protein inductors and using RFLP technique.
MATERIALS AND METHODS
Strains and induction media
The strains of P. chlamydosporia var. chlamydos-poria 10, 309, 399, from different geographical areas (Table 1) were kindly donated by the Rothamsted collection, at Nematode Interactions Unit, Rothamsted Research, Harpenden, UK. Strain IMI SD 187 of P. chlamydosporia var. catenulata and the strain 16 of P. chlamydosporia var. chlamydosporia were obtained from the Cuban collection, at the National Center for Animal and Plant Health, Havana, Cuba. The strains were maintained on corn meal agar (CMA) and subcultured on the same medium at 28ºC in the dark for two weeks.
Protein induction in liquid media
For protein induction in a basal liquid medium (14), two compounds "chemically related" to the egg shell structure of Meloidogyne eggs were investigated. These compounds are known to induce enzymes with hydrolytic activity (11). The treatments were as follows: i) basal liquid medium (as control); ii) basal liquid medium supplemented with 0,2% gelatine; iii) basal liquid medium supplemented with 1% chitin practical grade (Sigma) (sifted through a 30- mesh- pore screen) (11). For each treatment, 20 mL of medium was dispensed in sterile Corning 50 mL tubes, and inoculated with four 5 mm-diameter disks of fungal mycelium taken from the edge of a colony of a pure culture for each strain. The tubes were incubated at 28ºC, in the dark with continuous agitation in an orbital shaker at 120 rpm. Three replications per fungal strain were assayed.
The supernatants of the cultures were collected 7 days after fungal inoculation and filtered through filter paper Whatman Nº 1, concentrated by freeze-drying, and finally dissolved in 1 mL of sterile distilled water and then, maintained at -20ºC until they were used in the enzymatic analyses. The fungal biomass collected from each replication was dried in an oven at 80ºC for two days to determine the dry weight, except for those coming from the medium supplemented with chitin, using an analytic scale Sartorius, with an accuracy of 0,0001, and the protocol described in the US Pharmacopeia (15).
Quantification of proteins and enzymatic analyses
The concentration of total proteins was determined following the protocol described by Bradford (16), using the BIORAD kit and ELISA plates as support for the assay. The optical densities were recorded at 495 nm in a Multi Scan MRX (Dynex technologies Ltd., Billinghurst, West Sussex, UK). Serum bovine albumin (BSA) at concentrations ranging from 42 to 10 µg mL-1, was used to prepare the standard curve. The VCP1 activity was determined using N-succinyl-Ala-Ala-Phe-p-nitroanilide as substrate according to Morton et al. (12). In the assay, 2 µL of the enzymatic extract were mixed by agitation in an eppendorf tube with 100 µL of 2 mM substrate and 98 µL of buffer solution (0,1 M Tris HCl, pH 7,9). The absorbance was immediately read at 410 nm for 3 min, in a spectrophotometer for UV and visible light, CARY 50 using the program Kinetics. An activity unit was defined as the quantity of enzyme that releases 1 µmol of p-nitroanilide . min-1 . mL-1 (12). Three replications were done for all determination. Data were compared by Duncan's Test of Multiple Ranges using the SAS program (17); the fungal strain was considered as the independent variable and the indicators analysed in the time and in the particular medium.
Detection of VCP1 protease in strain IMI SD 187 by RFLP.
The genomic DNA of strains IMI SD 187 and 399 (used as reference for VCP1 gene) were obtained according to the extraction method described by Morton et al. (12).
For the RFLP, the genomic DNA was digested by the restriction enzymes BamHI, EcoRI, HindIII and SacI (Roche, Lewes, UK), using the buffer solution specified for each restriction enzyme and incubating the reaction mixture in a water bath at 37°C overnight. The digestion products were separated by electrophoresis on 1% agarose gel in 0,5X TAE at 70 volts. The gel was stained with Etidium Bromide (EtBr) (0,5 mg mL-1) to visualize the digestion products and transferred to a positively charged nylon membrane (Roche, UK) by capillarity overnight. The protocols for gel treatment and transfer were those described by "The DIG system user's for filter hybridisation" (Roche, UK). The gel was checked under UV light and the DNA fixed to the membrane with UV light, in the UV Stratalinker 1800 (Stratagene, UK). The probe used was a fragment of the VCP1 gene from the reference strain amplified by PCR with a couple of specific primers VCP1SF (5'-ATG CAA CTG TCT GTT CTT CTC-3') and VCP1R1 (5'- TAG TCA CAG CTT GTT CTC GCC-3') (12).
The amplified fragment was purified from agarose gel using the "High pure PCR product purification kit" (Roche, UK) and the product was diluted to 50 µL final volume. The labelling was developed using the "PCR DIG samples synthesis kit" (Roche, UK) and it was tested on 1,5% agarose gels. The recommendations followed for both, prehybridisation and hybridisation were described by "DIG high labeling and prevails detection starter kit II" and the user´s manual above referred (Roche, UK). The prehybridisation was carried out for 2 h at 42°C, while the hybridisation was developed overnight at the same temperature. Once hybridisation concluded, washes were as follows: 2 times with 2X SSC-0,1% SDS solution at room temperature for 5 min each time; and 2 times with 0,1X SSC-0,1% SDS solution at 68ºC for 15 min each time. The colour development was carried out according to the specifications described previously for Luminigen as substrate (Roche, UK). The membrane was exposed for 24 h for the final detection in the film.
RESULTS AND DISCUSSION
Protein induction in liquid medium
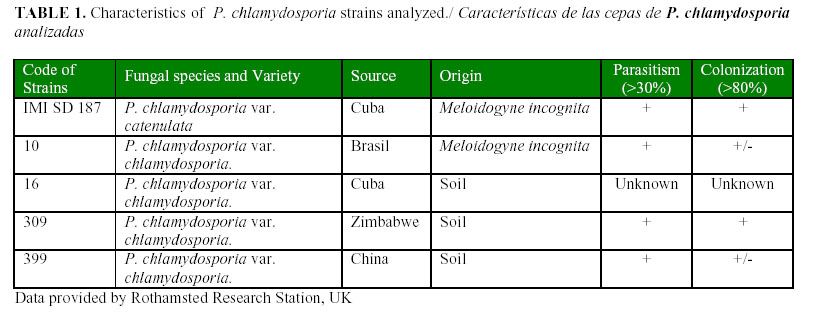
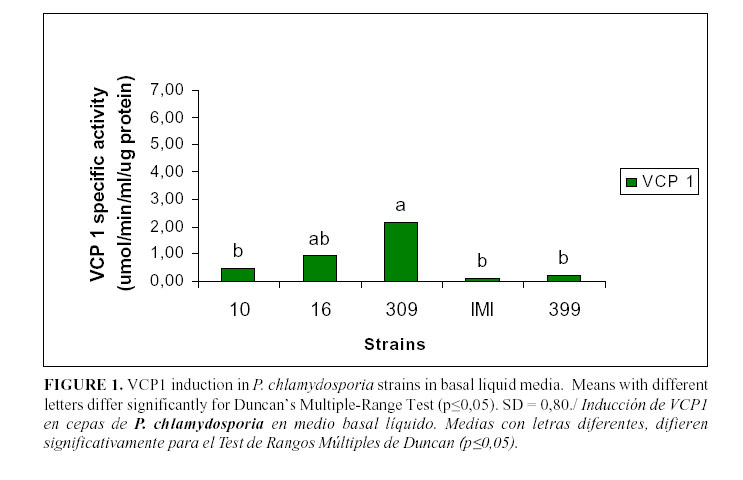
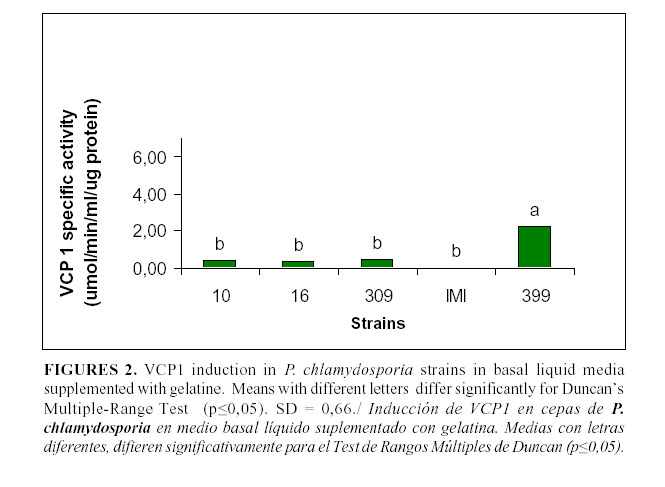
The levels of VCP1 production are shown in Figs. 1, 2 and 3. In the basal medium (Fig. 1), the highest values were reached by the strain 309 although they were not statistically different from those achieved by the strain 16; they were different from those of the remaining strains. In the medium supplemented with gelatine (Fig. 2), only the strain 399 showed a significant enzyme activity, whereas there were no statistical differences among the other strains. However, in the medium with chitin (Fig. 3), all the strains showed a high enzymatic activity induction, even strain IMI SD 187. In this medium, the strains 399 and 16 reached higher values (P<0,05), although the latter strain did not differ from the strains 10 or IMI SD 187.
Although the induction levels of proteaseVCP1 were higher in the medium with chitin, this medium is not recommended for the characterization and selection of isolates because it offers limited information about the enzymes. These results are similar to those of Segers et al., (11) working with the chlamydosporia variety strains and using the strain 10 as a reference. Therefore, the basic medium was a good alternative to study these enzymes and the potential of the isolates for their production. It has been previously stated that these enzymes are also inducible under conditions of scarce or limited nutrition (9), when the fungus has to face adverse conditions (e.g. lack of a carbon source) and then it synthesizes enzymes that are able to degrade other sources and, therefore, it changes from the saprophytic to its parasitic stage.
The basal medium is very poor in nutrients and this fact could have a positive effect on the enzymes related to the infection process induction.
According to the structure and chemical composition of the external layer of nematode eggs, Pochonia should have a selection of enzymes to degrade each layer, which appears as barriers, to arrive finally to the egg inner content and to infect it (11, 12). This process is complex and depends on molecular recognition, immobilization, penetration, colonization and digestion of the nematode egg.
In addition to the differences of the catenulata variety strain with regard to those of the chlamydosporia variety from the morphological point of view, variability within the strains of this last variety is also shown to occur because of the different responses resulting from this variety strain studied with different inductors (12, 18).
Detection of VCP1 protease in strain IMI SD 187 by RFLP.
The VCP1-encoding gene was not detected by RFLP in strain IMI SD 187, even when the stringency conditions were reduced to the minimum possible (data are not shown). Such negative results could suggest that the protease VCP1 gene in this strain has some difference from that in the strains of P. chlamydosporia var. chlamydosporia (11, 12).
At the protein level, Hirsch (personal communication, 2004) had experienced with VCP1 induction in impoverished Czapek medium for a group of strains, including the strain IMI SD 187. The low levels of VCP1 expression, even when studies of repression and induction with glucose and nitrates were carried out for this particular strain, drew this author's attention.
The family of the hydroxyalanine-proteases similar to subtilisin shares similar characteristic such as molecular mass, maximum activity in alkaline conditions, behaviour in front of inhibitors, but intra-specific differences have been found for the protease VCP1. These are supported by studies in the aminoacidic sequence that may have effects on the substrate used, the kinetics of the enzyme and its surface properties. Such differences could justify the use of different substrates to detect VCP1 activity in isolates from root-knot nematodes and those from cysts nematode (12).
In the present study, it was not possible to detect VCP1 gene in strain IMI SD 187 of P. chlamydosporia var. catenulata by RFLP/Hybridisation with a specific probe. This strain has also shown a very low protease activity. In the specific case of the strain IMI SD 187 and under these experimental conditions, it is convenient to talk about a VCP1-like protein.
The fact of a negative result at DNA level points towards more variations in the sequence of this protease in strain IMI SD 187. These variations did not allow the hybridisation with the specific probe. Such variations at protein level could produce changes in the recognition sites of the enzyme or steric impediment, resulting in an extremely low activity. The single presence, for example, of a glycine in strains coming from root-knot nematodes, instead of an alanine in those from cyst nematodes, can change the affinity for amino acids in the substrate union site named S3. Such differences among these enzymes caused high effects on their specificity (9, 12).
The low enzymatic activity points out to the existence of a VCP1 isoform or another different protease. But in both cases, a low affinity for the substrate tested was found. Recently, Atkins et al. (19), found 71 single nucleotide polymorphisms (SNPs) and 19 amino acid differences for the IMI SD 187 VCP1 sequence in comparison with P. chlamydosporia var. chlamydosporia isolate 10 (AJ427460), contrasting with the 15 SNPs and 4 amino acid differences seen when six P. chlamydosporia var. chlamydosporia VCP1 sequences were compared (12). Therefore, at least for this strain, this enzyme should not be considered as the virulence factor.
Segers (11) outlines that it is possible that homologous genes with products that differ in charge (as the isozymes detected in Pochonia) differ in their adsorption onto egg nematode shells, with consequences for pathogenicity. These assertions agree with the present results found in catenulata variety.
Keeping in mind the role attributed to VCP1 as a virulence factor in nematode egg infection by Pochonia (11), the level of the enzyme production could be recommended as a possible biochemical marker related to the strain virulence. This decision would be contradictory with the behaviour observed in IMI SD 187 strain in studies under field conditions. This strain effectively decreases nematode populations once applied to the soil, it is a good parasite, colonizer and survivor, with a high adaptation ability (5, 20, 21).
Lastly and not less important, if they are additionally considered the morphological distinguishing characteristics for the two Pochonia varieties (previously pointed out by Gams) and the differences at DNA level detected by the use of specific primers for each varieties (designed by Arora et al., (22); Atkins et al., (7)), the distinctive characteristics found in IMI SD 187 strain and the possible presence of a virulence factor different from VCP1; the distance between the two Pochonia varieties will become wider, which could even mean changes in an effective taxonomic classification. The results of this enzymatic characterization related to the egg infection process of host nematodes are the first achieved in a catenulata variety strain in the world. These hypotheses should be proved widening the range of catenulata variety strains and taking the chlamydosporia variety (strain 10) as a reference in a comparative study with more conclusive results in this regard.
ACKNOWLEDGMENTS
Rothamsted Research receives grant-aided support from the UK Biotechnology and Biological Sciences Research Council. B. Peteira was supported by funds provided by Rothamsted International. L. Hidalgo-Díaz was supported by EU Project (MiCoSPA, ICA 4-CT-2002-10044. Nematode populations were held in quarantine at Rothamsted Research in accordance with the Department for Environment Food and Rural Affairs Licence Nos. PHL 174C/4813 (04/2004).
REFERENCES
1. Hidalgo L, Brian K. Integration of biological control with other methods of nematode management. In: Ciancio A, Mukerji KG, editors. Integrated mana-gement and biocontrol of vegetable and grain crop nematodes. Serie Kluwer, Springer; 2007. p.28-43.
2. Gams W. A contribution to the knowledge of nematophagous species of Verticillium. Netherl J Plant Pathol. 1988;94:123-148.
3. Gams W, Zare R. A revision of Verticillium sect. Prostrata. III. Generic classification. Nova Hedwigia. 2001;73(3-4):329-337.
4. Hirsch PR, Mauchline TH, Mendum TA, Kerry BR. Detection of the nematophagous fungus Verticillium chlamydosporium in nematode-infested plant roots using PCR. Mycol Res. 2000;104:435-439.
5. Atkins SD, Hidalgo-Diaz L, Kalisz H, Mauchline TH, Hirsch PR, Kerry BR. Development of a new management strategy for the control of root-knot nematodes (Meloidogyne spp.) in organic vegetable production. Pest Manage Sci. 2002;59:183-189.
6. Peteira B, Puertas A, Hidalgo-Díaz L, Hirsch P, Atkins S. Real-time PCR to monitor and assess the efficacy of two types of inoculum of the nematophagous fungus Pochonia chlamydosporia var. catenulata against root-knot nematode populations in the field. Biotecnología Aplicada. 2005;22(4):261-266.
7. Atkins SD, Hidalgo-Diaz L, Clark IM, Morton CO, Montes de Oca N, Gray PA, Kerry BR. Approaches for monitoring the release of Pochonia chlamydosporia var. catenulata, a biological control agent of root-knot nematodes. Mycol Res. 2003;107(2):206-212.
8. López-Llorca LV. Ecología de los hongos nematófagos en el suelo y la rizosfera: modo de acción y aplicaciones prácticas. Conferencia magistral impartida en VI Seminario Internacional de Sanidad Vegetal. Sección Control biológico de nematodos parásitos de plantas. Palacio de Convenciones, La Habana, Cuba. 22-26 de septiembre de 2008.
9. Morton CO, Hirsch PR, Kerry BR. Infection of plant-parasitic nematodes by nematophagous fungi-a review of the application of molecular biology to understand infection process and to improve biological control. Nematol. 2004;6(2): 161-170.
10.López-Llorca LV, Janson HB, Macia Vicente JG, Salinas J. Nematophagous fungi as root endophytes. In: Schultz B, Boyle C, Sieber T, editors. Microbiol root endophytes. Serie Kluwer, Springer; 2006. p.191-206.
11.Segers R, Butt TM, Kerry BR, Beckett A, Feberdy JF. The role of the proteinase VCP1 produced by the nematophagous Verticillium chlamydosporium in the infection process of nematode eggs. Mycol Res. 1996;100:421-428.
12.Morton CO, Hirsch PR, Peberdy JP, Kerry BR. Cloning of and genetic variation in protease VCP1 from the nematophagous fungus Pochonia chlamydosporia. Mycol Res. 2003;107(1):38-46.
13.Kerry BR. Interaction on root and the efficacy of Pochonia chlamydosporia as a biological control agent. Conferencia magistral impartida en VI Seminario Internacional de Sanidad Vegetal. Sección Control biológico de nematodos parásitos de plantas. Palacio de Convenciones, La Habana, Cuba. 22-26 de septiembre del 2008.
14.Tikhonov VE, Lopez-Llorca LV, Salinas J, Jansson HB. Purification and characterization of chitinases from the nematophagous fungi Verticillium chlamydosporium and V. suchlasporium. Fungal Genet Biol. 2002;35:67-78.
15.USP. United States Pharmacopeia, Edition 28. (US Pharmacopeia Eds.) 2005.
16.Bradford M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein _ dye binding. Anal Biochem. 1976;31:426-428.
17.SAS INST. Statistical analysis system, Version 8.02, SAS Institute. Cary, NC, USA. 2001.
18.Mauchline TH, Kerry BR, Hirsch PR. The biocontrol fungus Pochonia chlamydosporia shows nematode host preference at the intraspecific level. Mycol Res. 2004;108(2):161-169.
19.Atkins SD, Peteira B, Clark IM, Kerry BR, Hirsch PR. Use of real-time quantitative PCR to investigate root and gall colonisation by co-inoculated isolates of the nematophagous fungus Pochonia chlamydosporia. Ann Appl Biol. 2009; In press.
20.Kerry B, Hidalgo L. Application of Pochonia chlamydosporia in the integrated control of root knot nematodes on organically grown vegetables crops in Cuba. IOBC/wprs Bull. 2004;27(1):123-127.
21.Montes de Oca N., Arévalo J, Acosta N, Hidalgo L. Herramientas para el control de la calidad de la cepa IMI SD 187 de Pochonia chlamydosporia var. catenulata (Kamyscho ex Barron y Onions) Zare y W. Gams. Rev Protección Veg. 2005;20(2): 86-92.
22.Arora DK, Hirsch PR, Kerry BR. PCR-based molecular discrimination of Verticillium chlamydosporium isolates. Mycol Res. 1996;100:801-809.
(Recibido 22-6-2009; Aceptado 25-6-2009)


{kind=link}
{kind=link}
{kind=link}
{kind=link}