










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión impresa ISSN 1010-2752versión On-line ISSN 2224-4697
Rev. Protección Veg. v.25 n.2 La Habana Mayo.-ago. 2010
Artículo reseña
POTYVIRUS: CARACTERÍSTICAS GENERALES, SITUACIÓN DE SU DIAGNÓSTICO Y DETERMINACIÓN DE SU PRESENCIA EN EL CULTIVO DEL PIMIENTO EN CUBA
POTYVIRUSES: GENERAL CHARACTERISTICS, SITUATION OF THE DIAGNOSIS AND DETERMINATION OF ITS PRESENCE IN PEPPER CROPS IN CUBA
Acela Díaz*, Madelaine Quiñones**, Franklyn Arana***, Marianela Soto**, Annia Hernández*
*Departamento de Microbiología y Virología. Facultad de Biología, Universidad de La Habana, Calle 25 esquina J, CP 10347, Ciudad de La Habana, Cuba. Correo electrónico: acela@fbio.uh.cu; **Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, La Habana, Cuba; ***Centro Universitario de Las Tunas. Las Tunas, Cuba
RESUMEN
El género Potyvirus es responsable de cuantiosas pérdidas económicas en el cultivo del pimiento (Capsicum annuum L.) en el mundo entero. Los miembros de este género pueden ser transmitidos a través de las semillas, de forma mecánica y por más de 25 especies de áfidos de forma no persistente, lo cual constituye un desafío para el desarrollo de estrategias efectivas de control. La problemática del diagnóstico de dichas entidades virales se ha abordado con diferentes enfoques; los cuales han transitado desde el análisis de síntomas en plantas indicadoras, la microscopía electrónica, el empleo de métodos serológicos e inmunoenzimáticos, hasta las metodologías moleculares de nueva generación. Dado el impacto que conllevan las pérdidas ocasionadas por estos virus en la mayoría de los cultivos de interés económico y el incremento a nivel mundial de estas infecciones, el presente artículo refiere aspectos sobre las principales características de este género viral, además de abordar resultados preliminares de la determinación de su presencia en el cultivo del pimiento en Cuba.
Palabras clave: potyvirus; diagnóstico; pimiento
ABSTRACT
The genus Potyvirus is responsible for important economic losses in the pepper crop (Capsicum annuum L.). Members of this genus can be transmitted by seeds and by more than 25 species of aphids in a non-persistent manner, what is a challenge for the development of effective control strategies. The diagnosis of these viral entities has been approached by different techniques such as the analysis of symptoms in specific plants, electronic microscopy, serological and inmunoenzymatic techniques, and even by using molecular methodologies of new generations. Due to the impact of the damages caused by potyviruses on most of the crops of economic interest and the increase of these infections around the world, the present paper refers to aspects about the main characteristics of this viral genus, in addition to preliminary results on the determination of its presence in the pepper crop in Cuba.
Key words: potyvirus; diagnosis; pepper
INTRODUCCIÓN
Desde el punto de vista histórico, los potyvirus fueron uno de los seis grupos de virus de plantas inicialmente reconocidos por Brandes y Wetter, en el año 1959, basándose en las propiedades morfológicas de las partículas virales. Desde entonces, el número de sus miembros, así como el conocimiento de sus propiedades características no ha cesado de aumentar (1).
Actualmente, tras la aceptación por el Comité Internacional de Taxonomía de Virus de un nuevo criterio taxonómico de clasificación, los potyvirus típicos se han constituido en el género Potyvirus, agrupado dentro de la familia Potyviridae. Son el grupo de virus de plantas más numeroso, constituyendo un 30% del total, con unos 180 miembros descritos y un continuo incremento (2).
Estos virus son transmitidos por semillas, de forma mecánica y por más de 25 especies de áfidos de forma no persistente, lo que constituye una característica distintiva del grupo (1,2,3). Asimismo, es un género responsable de graves pérdidas económicas en cultivos de interés agrícola, así como en pastos y plantas ornamentales (1).
Las enfermedades causadas por potyvirus no pueden ser controladas una vez que la planta es infectada, al igual que ocurre en otras enfermedades virales. Por otro lado, la transmisión no persistente por áfidos representa un desafío para las estrategias de prevención y control de las enfermedades virales en cultivos de importancia económica (2,4). Su modo de alimentación con inserciones cortas, de pocos segundos y su capacidad de inocular el virus en plantas sanas también a través de inserciones cortas, hace que el uso de insecticidas, de repelentes y de otros controles químicos pueda, no solo no evitar la difusión de la infección sino aumentarla, al favorecer un mayor desplazamiento planta a planta de los áfidos. Sin embargo, un conjunto de medidas de manejo integrado de las enfermedades que afectan a los cultivos, pudieran evitar la introducción de virus a plantaciones sanas y jardines o evitar su diseminación (5).
Por estas razones, el conocimiento de la presencia de estos virus en los diferentes cultivos, y el desarrollo de técnicas de diagnóstico rápidas y sensibles es de vital importancia en la actualidad en un país como Cuba, donde la agricultura es una prioridad. Este trabajo tiene como objetivos exponer las principales características de los potyvirus tratando algunos temas relevantes, como son los síntomas asociados a la presencia de potyvirus, las formas de transmisión de estas entidades y los principales métodos utilizados para su diagnóstico, así como abordar los resultados obtenidos en la determinación de la presencia de estas entidades en el cultivo del pimiento en Cuba.
PARTE ESPECIAL
Características generales de los potyvirus
Los potyvirus constituyen un grupo de virus de plantas caracterizados por presentar partículas filamentosas y flexuosas desnudas cuyo tamaño varía entre 680-900 nm de largo por 11-15 nm de an-cho. En algunos de sus miembros la longitud y flexuosidad de la partícula viral se ve afectada por la presencia de metales divalentes como el magnesio (2,6). Presentan como genoma una simple cadena de ARN de polaridad positiva de 9700 nucleótidos, con una proteína VPg unida covalentemente a su ex-tremo 5´ y una cola poli A en el extremo 3´ (el genoma de los Bymovirus está dividido en dos segmentos). Dicho genoma se expresa como una poliproteína de aproximadamente 330 kDa que posteriormente será procesada proteolíticamente para dar lugar a las proteínas funcionales (7,8).
Todos los miembros de la familia Potyviridae for-man cuerpos de inclusión cilíndricos y alargados de naturaleza cristalina en el citoplasma de las células de las plantas infectadas. Estas inclusiones de naturaleza proteica, presentan una estructura tridimensional característica de molinillo con aspas (1), que ha constituido un importante criterio taxonómico para su clasificación y agrupamiento.
La familia Potyviridae está integrada por seis géneros: Potyvirus, Ipomovirus, Macluravirus, Rymovirus, Tritimovirus y Bymovirus, de los cuales Potyvirus es el más conocido (9). Este es el género más amplio de todos los virus de plantas (91 especies descritas y 88 especies tentativas). Algunos de sus miembros son de importancia agrícola y causan grandes pérdidas económicas en el cultivo del pimiento como por ejemplo: el virus Y de la papa (PVY), el virus del grabado del tabaco (TEV), el virus del moteado del pimiento (PepMoV), el virus del moteado de las venas del pimiento (PVMV), el virus del moteado de las venas del chile (ChiVMV) y el virus del mosaico amarillo del pimiento (PepYMV), este último aislado recientemente en Brasil (1,10,11).
Organización genómica y función de los genes
Dentro de la familia Potyviridae, la mayoría de los estudios se han realizado en el género Potyvirus, demostrándose que todos los potyvirus, de un solo com-ponente como genoma (ya que el género Bymovirus posee dos moléculas de ARN como genoma) presentan similar estructura y organización genómica, la que difiere solo en algunos detalles (12).
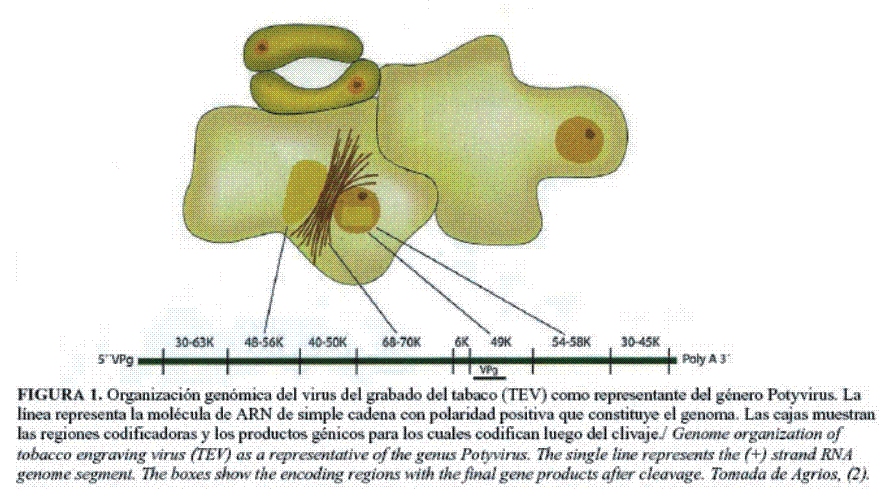
El virus del grabado del tabaco (TEV) constituye el miembro representativo de los potyvirus como género (3). Su estructura génica está compuesta por: una proteína Vpg de aproximadamente 24 kDa; una región 5´ no codificadora de alrededor de 144 nucleótidos rica en adenina y uracilo; un único marco de lectura abierto de 9161 nucleótidos que va a codificar para una gran poliproteína formada por 3000 aminoácidos (340 kDa) y una región en el extremo 3´ no traducida de aproximadamente 190 bases que termina en una cola de poli A (entre 20 y 160 adeninas) (Fig. 1). Este genoma codifica para una poliproteína que posteriormente será escindida por una proteasa viral, dando origen a nueve o diez productos génicos virales (13,14).
La cápsida de TEV está formada por aproximadamente 200 moléculas de una proteína de 30 kDa que es procesada a partir del extremo carboxilo terminal de la poliproteína (15). La proteína de la cápsida está relacionada con la transmisión exitosa de los potyvirus mediada por áfidos. Además, se conoce que la proteína Vpg que está unida covalentemente al extremo 5´ del genoma es procesada a partir de una región intermedia de la poliproteína (13).
En el citoplasma de las células infectadas por potyvirus se pueden encontrar cuerpos de inclusión con una forma característica de remolino constituidos por una proteína codificada por el virus y que ha sido denominada como CI. Algunos estudios in vitro señalan que CI podría estar relacionada con el movimiento célula-célula (16). En algunos casos, se pueden encontrar cuerpos amorfos de inclusión en el citoplasma de las células hospedantes que están constituidos por una proteína conocida como HC-Pro, involucrada en la transmisión mediada por áfidos y que posee un sitio de unión para el zinc cuya función no ha sido determinada (15).
En las células infectadas por TEV u otros miembros del género se forman, además, inclusiones nucleares de dos proteínas: NIa de 48 kDa y NIb de 58 kDa, las que son codificadas por todos los potyvirus. NIa tiene función proteasa y NIb tiene secuencias similares a las polimerasas de otros virus y es una ARN polimerasa ARN dependiente (15,17,18).
Replicación viral
El genoma de los potyvirus se expresa como una poliproteína que luego es procesada en alrededor de 10 productos génicos, de los cuales la mayoría están involucrados de una forma u otra en la replicación viral y constituyen el complejo de replicación viral: CI, 6K2, NIa-VPg y NIb. Como se mostró en el mapa del genoma (Fig. 1), estas proteínas forman un bloque y son análogas en términos de orden de los genes y de secuencia de los motivos a las proteínas 2C, 3A, 3B, 3C y 3D de los poliovirus, respectivamente. Por esta razón se cree que existen similitudes entre ambos grupos de virus en cuanto a las funciones y a las estrategias replicativas (18).
La proteína CI de los potyvirus posee motivos aminoácidos que son indicativos de su actividad helicasa. En el virus de la viruela del ciruelo (PPV) se ha podido demostrar que esta proteína es capaz de unirse al ARN y tener actividad ATPasa y helicasa. Una mutación en alguno de los siete motivos aminoácidos conservados de la misma puede traer consigo la abolición de su actividad helicasa y ATPasa, pero no así de su capacidad de unirse al ARN. Esto se ha constatado en estudios realizados con Escherichia coli Escherich que expresa la proteína CI de PPV (16).
La proteína 6K2 de TEV es una proteína de asociación con las membranas celulares y mediante es-tudios de mutagénesis dirigida se ha demostrado que está involucrada en la replicación viral (19) y que la asociación con la membrana se realiza a través de un dominio aminoácido hidrofóbico central (18). Utilizando proteínas de fusión fluorescentes que contienen a esta proteína de 6 kDa, Schaad et al. (20), mostraron que ella se asocia con una estructura vesicular alargada que se deriva del retículo endoplasmático. En células infectadas por TEV aparece el retículo colapsado en discretas estructuras agregadas y el ARN viral formando complejos replicativos asociados al retículo como si fueran membranas.
Una vez que el producto génico NIa es procesado en dos regiones, la región N-terminal da lugar a la proteína VPg y la región C-terminal da lugar a una proteasa (17). A su vez, la secuencia aminoacídica de la proteína NIb, presenta motivos que sugieren que se trata de una ARN polimerasa ARN dependiente. En experimentos de mutagénesis dirigida se ha demostrado que al mutar la región N-terminal de la proteína, queda abolida la capacidad replicativa de un clon infectivo de TEV en protoplastos.
Numerosos estudios han revelado una serie de interacciones entre varias de estas proteínas, de modo tal que forman un complejo replicativo. Se ha corroborado, utilizando mutantes condicionales y su-presores, que en TEV, NIb interactúa con la región C-terminal de NIa y en el virus del moteado de las venas del tabaco (TVMV) se ha identificado una interacción similar, pero en este caso sí se involucra además, la región N-terminal de la proteína NIa. Algunos autores plantean que la interacción entre NIa y NIb es esencial para la replicación del genoma de TEV. Las proteínas P1 y P3 a su vez interactúan entre sí y con CI, VPg, NIa y NIb, pero el significado de esta interacción aún no ha sido dilucidado. Es conocido además, que la proteína VPg está involucrada en la iniciación de la síntesis del ARN de la misma forma en que ocurre en los picornavirus (21,22,23).
Transmisión
Los potyvirus pueden diseminarse en la naturaleza mediante transmisión mecánica, a través de las semillas y por áfidos, mediante un proceso que se ha definido como no circulativo y no persistente (2).
La transmisión mecánica ocurre de diferentes for-mas: cuando entran en contacto las hojas de una planta enferma con las hojas de una planta sana, a través de las herramientas de trabajo en el campo, de las manos o las ropas de los trabajadores, así como de animales que se mueven de una zona enferma a otra sana llevando consigo las partículas virales y facilitando la expansión de la virosis en el espacio. Este tipo de transmisión ha sido fundamental en los estudios relacionados con la etiología de las enfermedades infecciosas en las plantas, ya que de forma general este es el tipo de transmisión que más se utiliza en los trabajos experimentales. Un sin número de virus como el virus X de la papa, el virus del mosaico del tabaco o el virus del mosaico del pepino, responsables de cuantiosas pérdidas en la agricultura, se transmiten de esta forma (2).
Más de 100 tipos de virus, son transmitidos además a través de las semillas, aunque en general, solo una porción de las semillas provenientes de las plantas infectadas van a transmitir la enfermedad (24,25,26). La frecuencia de transmisión depende en gran medida de la relación que se establece entre el virus y la planta hospedante y del estadio de desarrollo de la planta madre en el momento de la infección. En algunos casos, aproximadamente la mitad de las semillas transportan el virus y en otros el total de las semillas lo transmiten.
Por último, aunque no por ello menos importante, es necesario referirse a la transmisión por áfidos (23,27). Los áfidos, constituyen el grupo de insectos vectores de mayor importancia en la transmisión de los virus de plantas. Son insectos gregarios que forman colonias en las plantas infectadas. De forma general, diversas especies de áfidos son capaces de transmitir un mismo virus y a su vez una misma especie del vector puede transmitir una amplia gama de virus de plantas. En otros casos, la relación que se establece entre el virus y el vector es altamente específica.
Los potyvirus son transmitidos por los áfidos de forma no persistente, lo que implica que el insecto adquiere el virus después de permanecer solo unos segundos (alrededor de 30 segundos) en la planta enferma y posteriormente son capaces de inocularlo en la planta sana transfiriéndole la infección. Se utiliza también la clasificación de "no circulativo" porque el virus en ningún momento atraviesa las barreras del canal digestivo del vector, ni pasa a la hemolinfa o a las glándulas (2).
En Cuba, existen 83 especies de áfidos y se conocen 689 especies de plantas que son atacadas por estos, las que comprenden 419 géneros y 118 familias que incluyen un sexto de las especies cubanas de plantas superiores representando un tercio del número total de géneros y dos tercios del número de familias conocidas de Cuba (28). Entre las especies de áfidos capaces de transmitir diferentes especies de potyvirus se encuentran: Acyrthosiphon pisum (Harris), transmisor de PVY (virus Y de la papa) (29), Aphis craccivora Koch transmisor de TEV (virus del grabado del tabaco), PVMV (virus del moteado de la vena del pimentón), PVY, CVMV (virus del moteado de las venas del ají) (25), Aphis gossypii (Glover) transmisor de TEV, PVMV, PVY, CVMV, Myzus persicae (Sulzer) transmisor de PVY, TEV, PMV (30).
Síntomas asociados a la presencia de potyvirus
Los síntomas producidos como resultado de la infección con potyvirus pueden ser muy diversos y varían en dependencia de la especie afectada, el aislamiento presente y las condiciones ambientales (2). De manera general, en condiciones de campo, estos virus causan en las plantas infectadas un mosaico con moteado y arrugado de las hojas apicales y un bandeado oscuro de las venas de las hojas totalmente expandidas.
En todos los casos se produce una reducción en el desarrollo vegetativo que es más fuerte cuanto más severo es el ataque. La severidad de los síntomas depende de la edad de la planta, siendo más susceptibles las plantas jóvenes e intensificándose los mismos con el frío. El tamaño de la planta se reduce más cuanto más precoz ha sido la infección (1,2,15). Los síntomas aparecen entre los 7 y 21 días después de la infección y pueden producirse en todos los estados de crecimiento de la planta.
Se ha demostrado que el pimiento sufre graves daños cuando es infectado por el PVY, especialmente en los climas tropicales. Los síntomas se inician con un aclarado de las nervaduras de las hojas apicales, que pueden evolucionar pasando a tonos pardos, necrosándose el tejido. En estos casos, en ocasiones, aparece necrosis del pecíolo con caída de las hojas, quedando la planta defoliada, con necrosis apicales e incluso necrosis externas e internas del tallo. Las plantas pueden rebrotar, apareciendo las hojas con mosaicos en manchas de color verde oscuro-verde claro situados encima de las nervaduras (bandeado de venas), de modo que los limbos dejan de ser planos (2).
En pimiento también se pueden observar necrosis sobre las flores y los frutos. Algunas variedades presentan manchas pardas necróticas irregulares hundidas en el pericarpio y manchas necróticas en los pedúnculos. Otros síntomas que pueden aparecer son: enanismo, mosaico severo, deformación de hojas y frutos, que pueden presentarse arrugados, con manchas cloróticas y manchas necróticas, reducción en tamaño del fruto y aborto floral.
Sin embargo, en experimentos realizados por Aramburu et al. (23), pudo constatarse que algunas razas de pimiento, al ser inoculadas mecánicamente con aislados de PVY (cepas ordinarias o necróticas) no expresan síntomas visibles de infección, aunque al ser analizadas mediante la utiliza-ción de un ELISA-DAS las mismas reflejaron resultados positivos. También se ha demostrado que materiales tales como Solanum nigrum L., Solanum dulcamara L., Cirsium sp., Portulaca oleracea L. y Senecio vulgaris L., no presentan síntomas, pero actúan como reservorios naturales del virus en el área mediterránea. Esto indica que en ocasiones se puede encontrar una ausencia total de síntomas, aunque las plantas se encuentren infectadas por el virus; lo que podría estar dado porque las condiciones no sean las adecuadas para el desarrollo de los mismos o porque la planta infectada sea tolerante a la enfermedad. Este hecho es bastante frecuente en los virus vegetales (5,23), observándose fundamentalmente en plantas silvestres, muchas de las cuales constituyen fuentes naturales de resistencia viral.
En años recientes, se ha podido comprobar que diferentes regiones del genoma de los potyvirus desempeñan un importante papel en la aparición de síntomas en las plantas. Johansen et al. (3), mediante la construcción de híbridos recombinantes de dos cepas del virus del mosaico de la semilla del guisante (PSbMV), demostraron que el segmento del genoma que codifica para los genes NIa y NIb tiene una gran influencia en la aparición y severidad de los síntomas en Pisum sativum. Los análisis mutacionales del genoma del TVMV mostraron que la región que codifica para las proteínas P1 y HC-Pro, y particularmente el extremo 5’de HC-Pro, están involucradas en la expresión de síntomas en el cultivo del tabaco (15).
Diagnóstico de las enfermedades causadas por potyvirus.
La problemática del diagnóstico se ha abordado con diferentes enfoques, y todos han estado condi-cionados inevitablemente por el desarrollo y avance de la tecnología. El diagnóstico, específicamente de virus vegetales, ha transitado desde el análisis de síntomas en plantas indicadoras, la microscopía electrónica, el empleo de métodos serológicos e inmunoenzimáticos donde se destacan los ELISA y sus variantes, hasta las metodologías moleculares de nueva generación, como, la Reacción en Cadena de la Polimerasa (RCP) y la Hibridación de Ácidos Nucleicos (HAN) (5,26,31,32,33). Las técnicas inmunoquímicas y las técnicas moleculares son las principales vías para la identificación y diagnóstico de los potyvirus (28,29,34,35,36,37).
-Técnicas inmunoquímicas
Entre las técnicas inmunoquímicas que más se han utilizado para la identificación de los potyvirus se destacan las diferentes variantes de ELISA. Esta técnica inmunoenzimática forma parte de aquellas reacciones serológicas que utilizan conjugados para poder visualizar la reacción antígeno-anticuerpo y se basa en el uso de anticuerpos marcados con una enzima (generalmente la peroxidasa o la fosfatasa), de forma que los conjugados resultantes tengan actividad tanto inmunológica como enzimática (31,38). Al estar uno de los componentes (antígeno ó anticuerpo) insolubilizados sobre la placa, la reacción antígenoanticuerpo quedará inmovilizada y por tanto, podrá fácilmente ser revelada mediante la adición del sustrato, que al actuar sobre la enzima, producirá un color observable a simple vista o cuantificable mediante un equipo especializado. Es una técnica altamente sensible y de gran especificidad, que permite realizar en un corto espacio de tiempo estudios sobre grandes poblaciones, de manera sencilla y económica.
El ELISA presenta además, una buena reproducibilidad y facilidad en la interpretación de los resultados. Se han logrado grandes avances en esta técnica, por lo que es posible tanto la determinación de antígenos como de anticuerpos, con solo variar el tipo de ELISA. Por ejemplo, para la determinación de antígenos, la modalidad más frecuente es el ELISA Sandwicho directo (Fig. 2), sin embargo, para la determinación de antígeno se utilizan además, otras variantes, como el ELISA indirecto y el ELISA competitivo (38).
Fox et al. (38) determinaron que la utilización del DAS-ELISA para la detección de potyvirus presentes en tubérculos de papa almacenados a temperatura ambiente es factible hasta las 10 semanas post cosecha y considerablemente más económico que otros métodos. De igual modo, Aramburu et al. (23) realizaron trabajos de caracterización de aislamientos de PVY provenientes de plantas de tomate donde determinaron la presencia de partículas virales en plantas aparentemente sanas y en otros casos en plantas que presentaban infecciones mixtas. Otros autores como Mota et al. (39) y Huang et al. (40), han empleado otras variantes de ELISA (PTA-ELISA, y ELISA indirecto, respectivamente) para la detección de diferentes especies de potyvirus presentes en cultivos de interés agrícola.
-Diagnóstico molecular de los potyvirus
En la actualidad las técnicas moleculares desempeñan el papel protagónico en el diagnóstico de los potyvirus. Estas técnicas son superiores a las inmunoquímicas en especificidad y sensibilidad, además de ofrecer mayores posibilidades de detección y brindar una información más completa sobre los patógenos en cuestión (41). Muchos han sido los métodos empleados hasta el momento, demostrándose su factibilidad y permitiendo aislar e identificar a más de 30 virus que afectan a cultivos de importancia económica. Entre ellas la Hibridación de Ácidos Nucleicos (HAN) y la Reacción en Cadena de la Polimerasa (PCR), han sido las de mayor aplicación para lograr estos propósitos (6, 42,43).
La HAN se basa en el principio de la complementariedad de las bases que permiten la unión del ácido nucleico genómico del patógeno previamente fijado a un soporte sólido (normalmente membranas de nitrocelulosa o nylon), con moléculas complementarias que reciben el nombre de sonda (ya sea ADNc clonado o no, cebadores sintéticos o ARNc obtenidos por transcripción in vitro). En particular, el objetivo de la HAN es la detección específica de una secuencia de un ADN presente en una muestra biológica, con la ayuda de una sonda marcada, que se sintetiza in vitro empleando nucleótidos, marcados radioactivamente con 32P, o no radioactivamente con biotina o digoxigenina (32). La sonda que no se une es eliminada y los híbridos son detectados por métodos quimioluminiscente, colorimétricos o radiográficos (40,44).
La HAN sobre membrana incluye los siguientes pasos: 1) preparación de la muestra; 2) aplicación e inmovilización de la muestra; 3) prehibridacion; 4) hibridación con la secuencia complementaria marcada (sonda); 5) eliminación del exceso de la sonda con lavados y 6) detección del producto de hibridación (32,33).
Los límites de detección de virus purificados por hibridación con sondas radioactivas están en el ran-go de 1-100pg, superior a ELISA, mientras que con tejido infectado esta última parece tener un límite de detección mayor (33).
La hibridación radioactiva, permite la detección cualitativa y cuantitativa de bajas concentraciones de virus, aun usando extractos crudos de material vegetal que pueden ser en forma de savia extraída de la planta o en forma de impresiones del material vegetal (generalmente hojas) en la membrana de nitrocelulosa o nylon. Esta variante denominada “squash blot”, ha sido ensayada con buenos resultados en la detección del virus en el vector (41).
En la actualidad la hibridación con sondas no radioactivas desempeñan un papel fundamental en el diagnóstico cualitativo y cuantitativo de los virus en las plantas y el vector. Esta metodología requiere el desarrollo de procedimientos simples y rápidos para procesar el material vegetal de forma que permita la detección de los patógenos presentes. A pesar de las innumerables ventajas que posee esta técnica, tiene aparejado aspectos no ventajosos como las exigencias de bioseguridad en los casos en que se emplee el marcaje radioactivo y la inestabilidad de los isótopos, que tienen un tiempo de vida limitado.
Por otro lado, PCR se basa en el principio de la complementariedad de las bases de los ácidos nucleicos y la capacidad de síntesis del ADN por parte de una polimerasa. La PCR consiste en la síntesis in vitro de ácidos nucleicos, por lo que se puede amplificar específicamente un segmento determinado de ADN empleando dos cebadores que lo flanquean, para lo que se necesitan ciclos sucesivos de desnaturalización térmica del ADN, hibridación de los cebadores a las secuencias complementarias y extensión de los cebadores anillados mediante la enzima ADN polimerasa termoestable. Los productos de extensión son complementarios a los cebadores, como resultado en cada ciclo se duplica la cantidad de ADN sintetizada en el ciclo anterior, de forma que ocurre una amplificación exponencial del fragmento (43).
En el caso particular de los agentes virales cuyo material genético está constituido por ARN se utiliza una variante de PCR denominada Reacción en Cadena de la Polimerasa con Trascripción Inversa (RTRCP), la que requiere de una reacción preliminar de síntesis de una cadena de ADN complementaria al genoma viral, lo que se realiza con la utilización de una enzima reverso transcriptasa comercial. Los procesos de extracción y purificación de ARN se pueden considerar como los pasos más críticos e inestimados durante la detección por RT-PCR de los virus de genoma ARN. Dos etapas importantes a tener en cuenta durante el proceso de aislamiento de ARN, son el tratamiento y manipulación de la muestra previo a la extracción del ácido nucleico y el al-macenamiento del ARN una vez extraído. En la actualidad, existen diferentes tecnologías que permi-ten el aislamiento del ARN con un elevado grado de pureza. Estos métodos de extracción se pueden agrupar en dos grandes grupos: los métodos que se basan en la extracción en medio líquido y los métodos que extraen a través de sílica en columnas (34,44). El ARN es una molécula más fácil de degradar que el ADN de ahí que no es conveniente la cuantificación del mismo por espectrofotometría y algunos autores recomiendan verificar el proceso de extracción de ARN a través del proceso de detección por RT-PCR (9, 43,45).
A pesar de las ventajas de las metodologías de RT-PCR sobre los ensayos tradicionales, los prime-ros son muy vulnerables a los resultados falsos positivos y negativos. Esto hace necesario seguir estrictas prácticas de trabajo y utilizar múltiples controles negativos así como controles positivos. Los protocolos deben ser validados y se debe participar en ensayos de laboratorio que demuestren que el desempeño de estos ensayos es confiable.
Al hacer un análisis de la bibliografía consultada y los resultados obtenidos en Cuba con la aplicación de estas metodologías en el diagnóstico y caracterización de virus vegetales, se evidencia la factibilidad de las técnicas inmunoquímicas y moleculares para estos propósitos.
Potyvirus en el cultivo del pimiento
A nivel mundial, los cultivares de pimiento se han visto severamente afectados por las epifitias de origen viral; donde se destacan fundamentalmente: el virus Y de la papa (PVY), el virus del mosaico del pepino (CMV), el virus del estriado del tabaco (TSV), el virus del bronceado del tomate (TSWV), el virus del mosaico del tabaco (TMV), el virus del mosaico del tomate (ToMV), el virus del grabado del tabaco (TEV) y el virus del moteado del pimiento (Pep MoV) (10,11,18,46).
-Determinación de la presencia de potyvirus asociados al cultivo del pimiento en Cuba.
En Cuba, a pesar de los informes realizados por otros autores que señalan a los potyvirus como causantes de pérdidas importantes en el cultivo de pimiento (47,48), no se han realizado trabajos de prospección de dichas enfermedades en las principales áreas productoras. González et al. (47) informan la presencia de potyvirus en plantaciones de pimiento en Guira de Melena, provincia La Habana, mediante la utilización de técnicas de microscopía óptica y electrónica a través de las cuales detectaron la presencia de inclusiones citoplasmáticas fibrosas típicas del género Potyvirus. Además corroboraron la coincidencia de la sintomatología con la descrita en la literatura mediante la utilización de plantas indicadoras, en este caso Nicotiana glutinosa L.. Por otra parte, Depestre (48) señaló la presencia de entidades de este género viral en áreas de la región occidental, central y oriental, sobre la base de la presencia de síntomas similares a los descritos para estas entidades en el cultivo.
En este sentido se realizaron prospecciones en los principales macizos de producción de pimiento y se colectó un total de 212 muestras que mostraron síntomas de moteado, mosaicos leves y arrugamiento de las hojas, así como plantas que presentaron enanismo. Estas se evaluaron mediante la técnica ELISADAS (49) con anticuerpos policlonales genéricos a potyvirus cedidos gentilmente por el Dr. Roggero (Instituto de Fitovirologia Aplicada de Turín, Italia) y disponibles en el laboratorio de Virología Vegetal del Centro Nacional de Sanidad Agropecuaria (CENSA). La evaluación de las muestras permitió determinar un 66% de plantas infectadas por potyvirus en el total de plantas colectadas con síntomas característicos de la presencia de enfermedades virales.
Paralelamente, se trabajó en la puesta a punto de un ensayo de RT-PCR para lo cual se utilizaron los cebadores informados en la literatura Poly T y Poty 4 (50) los cuales amplificaron un fragmento de aproximadamente 2 Kb conteniendo el gen de la replicasa viral (Nib), gen de la proteína de la cápsida (cp) y un fragmento de la región 3’no traducida (UTR 3’). Resultados similares han informado otros autores, lo que demuestra la utilidad de estos cebadores y las condiciones óptimas de la reacción para la detección genérica de estos patógenos virales (5).
De igual forma, nuestros resultados argumentan la utilidad de las condiciones empleadas con vistas a disponer de una metodología de diagnóstico molecular factible de aplicar en estudios de variabilidad de estas entidades y que permita además la evaluación con gran rapidez y sensibilidad de plantas provenientes de programas de mejoramiento genético de cultivos de interés económico, donde la concentración en la cual se encuentran estos entes virales es muy baja.
Por otra parte es importante señalar que estos resultados aportan elementos a tener en cuenta en el manejo de estas entidades en el país y constituyen los primeros de este tipo para Cuba.
CONCLUSIONES
En la actualidad los potyvirus continúan siendo uno de los géneros de virus de plantas responsables de cuantiosas pérdidas económicas en cultivos de interés agrícola, además de afectar pastos y plantas ornamentales, debido fundamentalmente a las características y procesamiento de su genoma viral donde las recombinaciones y mutaciones juegan un papel determinante en el desarrollo y establecimiento de nuevas especies.
Por otra parte, el número de áfidos vectores transmisoras de estas enfermedades y los fenómenos ambientales asociados al cambio climático están condicionando la emergencia y reemergencia de las especies de este género que pudieran llegar a provocar epifitias de graves consecuencias y poner en riesgo la seguridad alimentaria.
En este sentido el conocimiento de la variabilidad de las especies circulantes en cada área o localidad y la posibilidad de disponer de métodos de diagnóstico altamente confiables, sensibles y específicos, constituyen elementos de gran valor para manejar consecuentemente los cultivos y contribuir al control de estas enfermedades.
REFERENCIAS
1. Hull R. Mattews´ Plant Virology. Cuarta Edición; 2002.
2. Agrios GN. Plant Diseases caused by Viruses. Plant Pathology. Fifth Edition. Elsevier Academic Press; 2005. p.724-820.
3. Johansen IE, Dougherty WG, Keller KE, Wang D, Hampton RO. Multiple viral determinants affect seed transmission of pea seedborne mosaic virus in Pisum sativum. J Gen Virol. 1996;77:3149-3154.
4. Spence NJ, Phiri NA, Hughes SL, Mwaniki A, Simons S, Oduor G, et al. Economic impact of Turnip mosaic virus. Cauliflower mosaic virus and Beet mosaic virus in three Kenyan vegetables. Plant Pathol. 2006;56:317-323.
5. Martínez Y, Fonseca D, Peralta EL. Conocimiento y diagnóstico del virus del bronceado del tomate (tswv), una necesidad de la agricultura cubana. Rev Protección Veg. 2005;20(1):11-19.
6. Ogawa T, Tomitaka Y, Nakagawa A, Ohshima K. Genetic structure of a population of Potato virus Y inducing potato tuber necrotic ringspot disease in Japan; comparison with North American and European populations. Virus Res. 2008;131:199-212.
7. López-Moya JJ, García JA. Potyviruses. Encyclopedia of Virology. Segunda Edición. Academic Press, San Diego; 1999.
8. Shukla DD, Ward CW, Brunt AA. The Potyviridae. CAB International, Wallingford, UK; 1994.
9. Susaimuthu J, Tzanetakis JI, Gergerich RC, Martin RR. A member of a new genus in the Potyviridae infects Rubus. Virus Res. 2008;131:145-151.
10. Rolland M, Glais L, Kerlan C, Jacquot E. A multiple single nucleotide polymorphisms interrogation assay for reliable Potato virus Y group and variant characterization. J Virol Methods. 2008;147:108-117.
11. Shukla DD, Ward CW. Amino acid sequence homology of coat proteins as a basis for identification and classification of the potyvirus group. J Gen Virol. 1988;69:2703-2710.
12. Rajamaki ML, Maki-Valkama T, Makinen K, Valkonen JPT. Infection with potyviruses. In: Talbot NJ, editor. Plant–Pathogen Interactions. Blackwell Publishing, Sheffield; 2004. p. 68-91.
13. Dolja VV, Haldeman R, Robertson NL, Dougherty WG, Carrington JC. Distinct functions of capsid protein in assembly and movement of tobacco etch potyvirus in plants. EMBO J. 1994;13:1482-1491.
14. Kelloniemi J, Kristiina M, Valkonen J. Three heterologous proteins simultaneously expressed from a chimeric potyvirus: Infectivity, stability and the correlation of genome and virion lengths. Virus Research. 2008;135:282-291.
15. Revers F, Le Gall O, Candresse T, Maule AJ. New advances in understanding the molecular biology of Plant/Potyvirus interactions. The Amer Phytopathol Soc. 1999;12(5):367-376.
16. Carrington J, Jensen PE, Schaad MC. Genetic evidence for an essential role for potyvirus CI protein in cell-to-cell movement. Plant J. 1998;14(4):393-400.
17. Puustinen P, Makinen K. Uridylylation of the Potyvirus VPg by Viral Replicase NIb Correlates with the Nucleotide Binding Capacity of VPg. J Biol Chem. 2004;279(37):38103-38110.
18. Viet Ha C. Detection and identification of potyvirus and geminivirus in Vietnam. Tesis presentada en opción al grado de Doctor en Filosofía. Universidad Tecnológica de Queensland. Australia; 2007.
19. Waltermann A, Maiss E. Detection of 6K1 as a mature protein of 6 kDa in plum pox virus-infected Nicotiana benthamiana. J Gen Virol. 2006;87(8):2381-2386.
20. Schaad MC, Lellis AD, Carrington JC. VPg of tobacco etch Potyvirus is a host genotype-specific determinant for long distance movement. J Virol. 1997;71:8624-8631.
21. Hafren A, Makinen K. Purification of viral genome-linked protein VPg from potato virus A-infected plants reveals several post-translationally modified forms of the protein. J Gen Virol. 2008;89(6):1509-1518.
22. Satheshkumar PS, Gayathri P, Prasad K, Savithrih S. Natively Unfolded; VPg Is Essential for Sesbania Mosaic Virus Serine Protease Activity. J Biol Chem. 2005;280(34):30291-30300.
23. Aramburu J, Galipienso L, Matas M. Characterization of potato virus Y isolates from tomato crops in northeast Spain. Eur J Plant Pathol. 2006;115:247-258.
24. Alegbejo M. Aphid transmission of pepper veinal mottle virus. Samaru J Agric Res. 1986;4(1-2):7177.
25. Hull R, Al-Hakim A. Nucleic acid hybridization in plant virus diagnosis and characterization. TIBTECH. 1988;6:213-218.
26. Abou-Jawdah Y, Sobh H, El-Zammar S, Fayyad A, Lecoq H. Incidence and management of virus diseases of cucurbits in Lebanon. Crop Protection. 2000; 19:217-224.
27. Holman J. Los áfidos de Cuba. Instituto del Libro; 1974.
28. Ferreres A, Pérez P, Gemeno C, Ponz F. Transmission of spanish pepper and potato PVY isolates by aphid (Homoptera: Aphididae) vectors: epidemiological implications. Environ Entomol. 1993;22(6):1260-1265.
29. Sánchez M, Cermeli M, Machado W, Centeno F, Brown E. Diversidad de áfidos (Homoptera: Aphididae) capturados con trampas amarillas en el cultivo de pimentón (Capsicum annuum L.) y su relación con los factores climáticos. Bol Entomol Venez. 2000;15(1):61-83.
30. Gonsalves D, Trujillo EE. Tomato Spotted Wilt Virus in papaya and detection of the virus by ELISA. Plant Dis. 1986;70:501-506.
31. Quiñones M, Fonseca D, Gómez O, Miranda I, Piñón M, Martínez Y. Optimización y aplicación de la hibridación de ácidos nucleicos no radioactiva para el diagnóstico del virus del encrespamiento amarillo de la hoja del tomate (TYLCV) en el programa de mejoramiento genético. Rev Protección Veg. 2003;18(3):176-182.
32. Quiñones M, Fonseca D, Martínez Y. Comparación de métodos de hibridación de ácidos nucleicos para el diagnóstico del virus del encrespamiento amarillo de la hoja del tomate (TYLCV) en Cuba. Rev Protección Veg. 2004;19(1):26-32.
33. Blacksell SD, Khounsy S, Westbury HA. The effect of simple degradation and RNA stabilization on classical swine fever virus RT-PCR and ELISA methods. J Virol Methods. 2004;118:33-37.
34. Chen J, Chen JP, Langeveld SA, Derks AFLM, Adams MJ. Molecular characterization of Carla-and Potyviruses from Narcissus in China. J Phytopathol. 2003;151:26-29.
35. Chen J, Zheng HY, Shi YH, Adams MJ, Wei CB, Lin L, et al. Detection and characterisation of a second potyvirus from Thunberg fritillary in China. Arch Virol. 2006;151:439-447.
36. Raboudi F, Ben Moussa A, Makni H, Marrakchi M, Makni M. Serological detection of plant viruses in their aphid vectors and host plants in Tunisia. OEPP Bull. 32:495-498.
37. Acebo Y, Hernández A. Las técnicas moleculares para la identificación de bacterias de importancia agrícola. Su contribución a la conservación del medio ambiente. Rev Prot Amb. 2004;5:34-40.
38. Fox A, Evans F, Browning I. Direct tuber testing for Potato Y potyvirus by real time RT-PCR and ELISA: reliable options for post-harvest testing?. OEPP Bull. 2004;35:93-97.
39. Mota LDC, Della Vecchia MGS, Gioria R, Kitajima EW, Rezende JMA, Camargo LEA, et al. Pfaffia mosaic virus: a new potyvirus found infecting Pfaffia glomerata in Brazil. Plant Pathol. 2004;53:368-373.
40. Huang CH, Hu WC, Yang TC, Chang YC. Zantedeschia mild mosaic virus, a new widespread virus in calla lily, detected by ELISA, dot blot hibridization and IC-RT-PCR. Plant Pathol. 2007;56:186-189.
41. Louro D, Accotto GP, Vaira AM. Occurrence and diagnosis of Tomato chlorosis virus in Portugal. Eur J Plant Pathol. 2000;106:589-592.
42. González L, Soto M, Ortíz MI, Peralta EL. Determinación de parámetros analíticos de las técnicas de hibridación de ácidos nucleicos para la detección del viroide del tubérculo ahusado de la papa (PSTVd). Fitopatología. 2001;36(1):24-33.
43. Chirgwin JM, Przybyla AE, MacDonald RJ, Rutter WJ. Isolation of biologically active ribonucleic acid from sources enriched in ribonuclease. Biochemistry. 1979;18:5294-5299.
44. Martínez Y, Fonseca D, Quiñones M, Peralta EL. Evaluación de parámetros analíticos para la detección molecular de geminivirus que afectan el cultivo del tomate en Cuba. Rev Protección Veg. 2001;16(1):55-61.
45. González L. Principales técnicas moleculares aplicadas al estudio de las plantas. Facultad de Biología, Universidad de La Habana; 2006.
46. Inoue-Nagata AK, Fonseca MEN, Resende RO, Boiteux LS, Monte DC, Dusi AN, et al. Pepper yellow mosaic virus, a new potyvirus in sweetpepper, Capsicum annuum. Arch Virol. 2002;147:849-855.
47. González G, Font C, Valdés S. Diagnóstico de virus vegetales a nivel de grupo en el cultivo del pimentón (Capsicum annuum L.) mediante la técnica de microscopía óptica. Fitosanidad. 2002;6(3):3-9.
48. Depestre T. An approach to peppers breeding in Cuba Capsicum. Newsletter. 1999; 18:16-20.
49. Clark MF, Adams AN. Characteristics of the Microplae Meted of Enzyme Linked Immunoabsorbent Assay (ELISA) for the Detection of Plant Viruses. J Gen Virol. 1977;34:475-483.
50. Trutta AAC, Sousa ARR, Nascimento AVS, Pereira RC, Pinto CMF, Zerbini F, et al. Identidade e propriedades de isolados de potyvírus provenientes de Capsicum spp. Fitopatol bras. 2004;29(2):160168.
(Recibido 3-10-2008; Aceptado 3-12-2009)


{kind=link}
{kind=link}