










Meu SciELO
 Serviços customizados
Serviços customizadosServiços Personalizados
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Protección Vegetal
versão On-line ISSN 2224-4697
Rev. Protección Veg. v.25 n.2 La Habana maio-ago. 2010
Artículo original
DIVERSIDAD GENÉTICA DE ESPECIES SILVESTRES DEL GÉNERO Nicotiana I: CARACTERIZACIÓN MEDIANTE MARCADORES BIOQUÍMICOS
GENETIC DIVERSITY OF WILD SPECIES OF Nicotiana genus I: CHARACTERIZATION USING BIOCHEMICAL MARKERS
Marlyn Valdés de la Cruz*, Clara González*, Regla M. Lara**, María Isabel Román***,Yuniet Hernández**, Reina Margarita Hernández****, Mileidy Cabrera*****, G. Torrecilla*****
*Facultad de Biología, Universidad de la Habana, Calle 25 No. 455 % I y J Vedado Ciudad de la Habana,Cuba. Correo electrónico: marlyn@fbio.uh.cu. **Instituto Nacional de Ciencias Agrícolas (INCA),Carretera a Tapaste km 3.5 San José de las Lajas Provincia La Habana, Cuba. ***Institutode Investigaciones en Viandas Tropicales, Finca 3 Carolina, Santo Domingo Villa Clara, Cuba.****Centro Universitario de la Isla de la Juventud, Carretera Aeropuerto Km 3 1/2, Isla de la Juventud, Cuba. *****Estación Experimental de Tabaco de Cabaiguan, Carretera Sta Lucía Km 2, Cabaiguán, Sancti Spíritus, Cuba
RESUMEN
La caracterización de especies silvestres del género Nicotiana resulta de gran interés en los programas de mejoramiento genético de la especie Nicotiana tabacum L. ya que pueden ser empleadas como fuente de genes de resistencia a plagas, a la vez que permiten incrementar la variabilidad genética en la única especie comercial del género en el país. El presente trabajo tuvo como objetivo caracterizar la variabilidad genética en 10 especies del banco de germoplasma del género Nicotiana en Cuba, mediante el análisis de nueve sistemas isoenzimáticos y las proteínas totales. Se emplearon los sistemas peroxidasas, polifenoloxidasas, esterasas, anhidrasa carbónica, superóxido dismutasa, diaforasa, fosfatasas ácidas, fosfatasas alcalinas, enzima málica, así como las proteínas totales. Se realizaron las electroforesis en geles de poliacrilamida (PAGE) en lámina vertical. El grado de polimorfismo total entre las especies para los sistemas estudiados fue de un 92,7%, siendo los sistemas peroxidasas, fosfatasas ácidas y enzima málica los más polimórficos con un 100%. Estos sistemas junto a las fosfatasas alcalinas y las proteínas totales fueron los de mayor importancia en la identificación y caracterización de estas especies por presentar patrones de bandas propios para cada una de ellas. Este resultado constituye el primer estudio isoenzimático en especies silvestres del banco de germoplasma del género Nicotiana en nuestro país, lo que permitió conocer el grado de variabilidad presente en dichas especies.
Palabras clave: Nicotiana; diversidad genética; polimorfismo; marcadores bioquímicos
ABSTRACT
The characterization of wild species of the genus Nicotiana is very important for the genetic breeding programs of Nicotiana tabacum L. because they can be a source of pest resistance genes to increase the genetic variability in the only commercial species of this genus in Cuba. The objective of this work was to characterize the genetic variability in ten species of the Nicotiana genus of the germoplasm bank in Cuba through the analysis of nine isozyme systems and total proteins. The isozymes assayed were peroxidase, polyphenol oxidase, esterase, carbonic anhidrase, superoxide dismutase, diaphorase, acid phosphatase, alkaline phosphatase and malic enzyme, as well as total proteins. Isozyme electrophoresis was performed on vertical slab polyacrylamide gels (PAGE). The total polymorphism level among species in the systems studied was 92,7%, where the most polymorphic systems were peroxidase, acid phosphatase and malic enzyme with 100%. These polymorphic systems, jointly with alkaline phosphatase and total protein, were the most important markers for the identification and characterization of Nicotiana species since they allowed distinguishing band patterns for each of them. This result is a first approach to the isozyme studies in wild species from the germoplasm bank in Cuba, leading to the knowledge of the variability level of those species.
Key words: Nicotiana; genetic diversity; polymorphism; biochemical markers
INTRODUCCIÓN
En el curso de la historia los recursos fitogenéticos han contribuido a la estabilidad de los agroecosistemas, proporcionando la materia prima fundamental para el surgimiento del fitomejoramiento científico moderno. En este sentido, los bancos de germoplasma surgen como una respuesta a la necesidad de conservar el patrimonio genético vegetal. La caracterización de la variabilidad genética contenida en ellos constituye un factor decisivo para garantizar su utilización eficiente y racional en los programas de mejoramiento genético (1,2).
El género Nicotiana es un miembro de la familia Solanaceae y se ha dividido en tres subgéneros (Rustica, Tabacum y Petunioides), los que contienen alrededor de 76 especies reconocidas (3,4). De ellas, Nicotiana tabacum L. y Nicotiana rustica L. son las únicas especies cultivadas comercialmente en el mundo, el resto son consideradas como silvestres y la mayoría poseen caracteres morfológicos muy diferentes a los de las variedades comerciales. Sin embargo, estas especies silvestres conservan genes de resistencia a plagas, así como para rasgos cualitativos y fitoquímicos importantes que no están presentes en las variedades cultivadas (5).
Se ha encontrado una pérdida considerable de la variabilidad genética en las variedades comerciales, detectada mediante estudios isoenzimáticos y de marcadores moleculares de ADN (6,7,8,9); debido fundamentalmente a la preferencia de los cultivadores por determinadas variedades, a los métodos uti-lizados en la obtención de las mismas a partir de progenitores comunes y a la selección artificial continua a favor de aquellos caracteres de interés comercial (9,10). En tal sentido, las especies silvestres ofrecen un potencial genético para su empleo en los programas de mejoramiento con el fin de introducir genes de resistencia a plagas como la pata prieta (Phytophthora parasitica var. nicotianae Breda de Haan), moho azul (Peronospora hyosciamy f.sp tabacina Adam) (11,12,13), además de contribuir a incrementar la variabilidad genética de las variedades comerciales.
Para el estudio de la variabilidad genética entre individuos se han utilizado varios tipos de marcado-res: morfológicos, isoenzimas, proteínas y marcadores basados en ADN, siendo las isoenzimas los pri-meros marcadores moleculares que se emplearon en la mejora genética con múltiples aplicaciones (14,15,16). Las isoenzimas son marcadores general-mente codominantes, fenotípicamente neutros y la metodología empleada para su análisis es rápida, sencilla y económica (16,17,18).
El objetivo del presente trabajo fue caracterizar la variabilidad genética en 10 especies del banco de germoplasma del género Nicotiana en Cuba, mediante el análisis de nueve sistemas isoenzimáticos y las proteínas totales.
MATERIALES Y MÉTODOS
Se estudiaron diez especies silvestes del género Nicotiana de diferentes subgéneros, secciones y orí-genes (4), pertenecientes al banco de germoplasma de la Estación Experimental de Tabaco de Cabaiguán ubicado en la provincia de Sancti Spíritus (Tabla 1).
Las semillas de cada especie se sembraron en bandejas de poliestireno expandido con alvéolos (cepellones) que contenían una mezcla de materia orgánica y zeolita (1:1), siguiendo la tecnología de bandejas flotantes. Las mismas se colocaron en condiciones de cultivo protegido o túneles con luz y temperatura natural (19). Pasados los 45 días, las plantas se trasplantaron al campo sobre suelo pardo sialítico, donde la fertilización, el riego y las condiciones fitosanitarias se realizaron según lo establecido por Espino (19) para el cultivo del tabaco. Las muestras se extrajeron de plantas de 60 días de trasplantadas, para lo cual se colectaron al azar hojas jóvenes y sanas de cinco individuos por especie.
Análisis bioquímicos
Los extractos foliares se obtuvieron a partir de 5g de las hojas colectadas de cada especie, macerados con 10 gotas de una solución de sacarosa al 20% y se conservaron a -15oC hasta su utilización. Las corridas electroforéticas se realizaron en gel de poliacrilamida (PAGE), utilizando una cámara de corrida en lámina vertical y sistema de tampones discontinuos de Tris–glicina 0,04M pH 8,3; con una intensidad de la corriente de 50mA (17).
Para cada sistema estudiado se aplicaron las tinciones específicas (Tabla 2). Después de teñidos los geles se lavaron con agua destilada y se conservaron en una solución de ácido acético al 10% hasta el momento en que se midieron las movilidades electroforéticas (17). Se realizaron dos réplicas por muestra en cada gel y estos se repitieron tres veces para cada sistema analizado.
Análisis estadístico
Para determinar los patrones de bandas de las isoenzimas y las proteínas totales de las especies estudiadas se aplicó el programa MAT-GEN (22). A partir de los resultados obtenidos se construyó la matriz binaria, asignándosele a la presencia de bandas (1) y a la ausencia (0) para cada sistema. La mis-ma se procesó por el paquete de programas estadísticos NTSYS-PC versión 2.1 (23), se empleó el coeficiente de Dice (24) y se aplicó el método de las medias aritméticas de grupos no ponderados (UPGMA) del programa SAHN para producir el dendrograma. Los grupos en este se formaron a partir del 50% de afinidad.
RESULTADOS
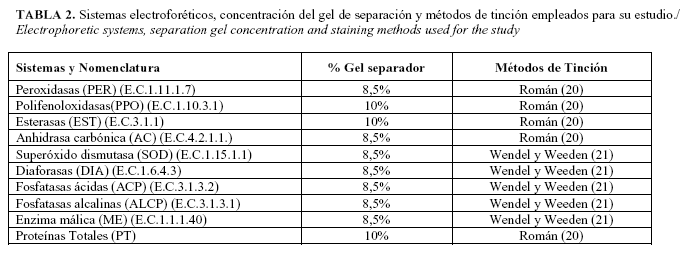
Para las isoenzimas peroxidasas (Fig. 1) aparece un total de 12 bandas, todas polimórficas, lo que favorece que se presenten diez zimotipos, cada uno propio para las especies estudiadas. Nicotiana alata Link et Otto presenta el mayor número de isoformas con once bandas y el menor número aparece en la especie Nicotiana suaveolens Lehm. con cinco bandas.
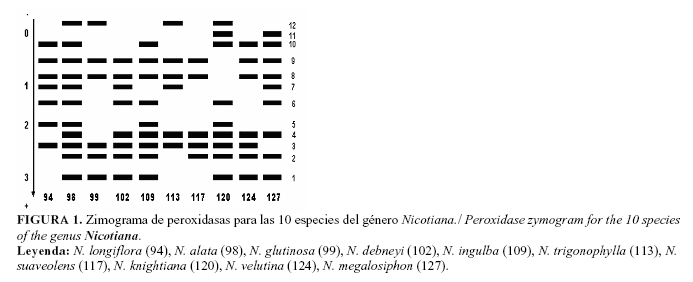
El zimograma para el sistema isoenzimático polifenoloxidasas (Fig. 2), presenta un total de siete bandas y solamente la sexta es común a las diez especies. Se observan cinco zimotipos diferentes, don-de las especies N. alata, N.glutinosa L., N. debneyi Domin., N. ingulba J.M. Black., N. trigonophylla Dunal. y N. velutina Wheeler, muestran patrones de bandas idénticos y son además las que poseen el total de isoformas. La especie N. knightiana Goodps. sólo presenta tres bandas, siendo la especie de menor número dentro del sistema. El polimorfismo obtenido en este caso permite diferenciar las especies N. longiflora Cav., N. suaveolens, N. knightiana y N. megalosiphon Van Huerck and Mull Arg. ya que para el resto resultó ser monomórfico.
En el sistema esterasas se puede observar un total de nueve bandas (Fig. 3); la número cinco es co-mún a todos los materiales y la seis sólo se encuentra ausente en la especie N. alata. En este sistema se presenta un total de seis zimotipos, uno de los cuales es común a las especies N. glutinosa, N. ingulba Black y N. megalosiphon. El mayor número de isoformas corresponde a la especie N. trigonophylla con seis bandas y el menor a las especies N. suaveolens y N. knightiana con dos bandas, las que tienen zimotipos comunes.
Las especies N. debneyi y N. velutina presentan también idéntico patrón de bandas. La banda de ma-yor migración anódica sólo aparece en N.alata, lo que la hace diferente del resto de las especies estudiadas.
Al observar el zimograma del sistema anhidrasa carbónica (Fig. 4) se detecta un total de nueve bandas, sólo la quinta es común a todos los materiales. Este sistema presenta siete zimotipos diferentes, ya que coinciden las especies N. longiflora y N. knightiana con cuatro bandas. Las especies N. debneyi, N. suaveolens y N. velutina presentan idénticos patrones electroforéticos con el menor número de bandas (dos), mientras que las especies N. alata, N. trigonophylla y N. megalosiphon muestran el mayor número de bandas (seis).
En el zimograma del sistema superóxido dismutasa (Fig. 5) aparecen un total de siete bandas y solamente la sexta está presente en todos los materiales. Existen ocho zimotipos diferentes y sólo coinciden en su patrón de bandas las especies N. suaveolens y N. knightiana, así como N. alata y N. debneyi; estas últimas conjuntamente con N. longiflora presentan el menor número de bandas (dos), mientras que la especie N. trigonophylla posee el mayor número de bandas (cinco).
Para las isoenzimas diaforasas (Fig. 6) se presentan un total de nueve bandas, la cuarta es común a todas las especies y la banda cinco está ausente sólo en N. knightiana. Se muestran nueve zimotipos diferentes, N. alata y N. suaveolens tiene el mayor número de bandas (seis) y la especie N. velutina el menor con tres bandas solamente.
En el sistema fosfatasas ácidas (Fig. 7) se puede observar un total de nueve bandas, todas polimórficas y cada especie presenta un zimotipo propio. La especie N. trigonophylla posee el mayor número de bandas con ocho y las especies N. glutinosa y N. debneyi el menor número de bandas con sólo tres.
En el sistema fosfatasas alcalinas (Fig. 8) se muestran diez bandas diferentes, donde la cuarta banda está presente en todas las especies; sin embargo, es posible identificar zimotipos propios para cada una de ellas. La especie de N. trigonophylla presenta el mayor número de bandas (seis) y N. suaveolens el menor número de bandas (dos).
Los resultados correspondientes a la enzima málica se muestran en la Figura 9 en el que se obtiene un total de 13 bandas diferentes, ninguna común a todos los materiales, cada especie presenta un zimotipo propio. La especie N. trigonophylla posee el mayor número de bandas con un total de nueve, y las especies N. knightiana y N. velutina el me-nor número de bandas, con solo dos ubicadas en diferentes posiciones.
Al analizar el proteinograma (Fig. 10) se observa un total de 24 bandas. Las número 12 y 13 son comunes a todas las especies. N. trigonophylla es la que presenta un mayor número de isoformas con un total de 19 y N. knightiana el menor número con nueve bandas. Esta distribución de bandas hace que no existan especies con el mismo patrón, aunque las mayores similitudes se encuentran entre las especies N. velutina y N. megalosiphon con 14 bandas comunes y las especies N. trigonophylla y N. suaveolens que poseen 13 bandas comunes.
Los resultados del análisis cuantitativo de los electroforetogramas, para las especies estudiadas se muestran en la Tabla 3. Se observa un alto grado de variabilidad, ya que de 110 bandas analizadas el 92,7% resultaron polimórficas. Los sistemas peroxidasas, fosfatasas ácidas, fosfatasas alcalinas y la enzima málica así como las proteínas totales presentaron los mayores valores de polimorfismo y coincide con los que presentan zimotipos propios para cada especie. El porcentaje de polimorfismo para cada sistema es alto, superior al 85 % en todos los casos.
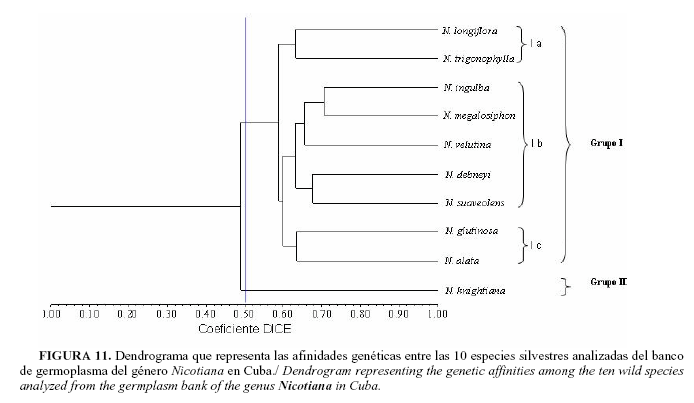
En los resultados del análisis de conglomerado de las 10 especies del género Nicotiana para los nueve sistemas isoenzimáticos analizados y proteínas totales se aprecia la formación de dos grupos a partir de un índice de similitud de 0.50 (Fig. 11). En el grupo I se incluyen las especies estudiadas pertenecientes al subgénero Petunioide y en el grupo II, la especie N. knightiana del subgénero Rustica.
Dentro del grupo l se puede distinguir la formación de tres subgrupos. En el I se incluyen las especies N. longiflora y N trigonophylla, pertenecientes a las secciones Alatae y Trigonophyllae respectivamente. En el subgrupo l-b se incluyen las especiesN. debneyi, N. ingulba, N. suaveolens, N. velutina y N. megalosiphon, todas pertenecientes a la sección Suaveolentes. El subgrupo l-c incluye las especies N. alata y N. glutinosa las que pertenecen a las secciones Alatae y Undulatae, respectivamente.
DISCUSIÓN
A pesar del desarrollo alcanzado en los estudios genéticos por los marcadores moleculares basados en el ADN, los análisis isoenzimáticos han demostrado mantener su utilidad en la determinación de la diversidad genética en poblaciones, géneros y especies. Las técnicas isoenzimáticas son relativamente económicas, prácticas y factibles de utilizar, aún cuando se analiza un gran número de muestras, ya que mediante la comparación de los patrones de bandas de las isoenzimas analizadas, se puede proporcionar una estimación del grado de relación genética entre los genotipos analizados (17,25).
En un análisis de conjunto de todos los sistemas electroforéticos empleados se observa el alto nivel de polimorfismo obtenido para las especies analizadas, lo que demuestra la variabilidad genética presente entre ellas. Estos resultados contrastan con los bajos niveles de polimorfismo obtenidos por Pérez (6) y Rodríguez (7) (21,9% y 23% respectivamente), en estudios realizados en variedades comerciales de N. tabacum con el empleo de los mismos sistemas, todo lo cual evidencia la necesidad de incrementar la variabilidad genética en los cultivares de tabaco y demuestra las potencialidades que tienen las especies silvestres en este sentido.
Por otra parte, el hecho de obtener patrones de bandas propios de cada especie para los sistemas peroxidasas, fosfatasas ácidas, fosfatasas alcalinas y enzima málica, así como las proteínas totales, per-mite considerarlos como marcadores de gran importancia en la identificación del genoma y recomendarlos en la caracterización de especies del banco de germoplasma del género Nicotiana.
En la literatura revisada sólo se encontró el trabajo de Smith et al. (26), en el que utiliza los dos sistemas isoenzimas peroxidasas y esterasas como marcadores para determinar las relaciones genéticas entre un grupo de especies de este género, incluidas todas las analizadas en la presente investigación. Los resultados obtenidos para las isoenzimas peroxidasas coinciden plenamente con los informados por estos autores, en cuanto al rango en que se encuentra el número de bandas obtenidos para las especies, el no obtener alguna banda común a todas las especies y por tanto propia del género, así como la obtención de patrones de bandas propios de cada especie.
Sin embargo, en cuanto a los resultados informados por dichos autores para las esterasas, no existe coincidencia con los resultados obtenidos en este trabajo pues informaron un mayor número de bandas (15). Además, no aprecian bandas propias del género, aunque detectan zimotipos propios de cada especie. Las diferencias encontradas pueden deberse a que, aunque son las mismas especies, las condiciones de cultivo y época en que se realizó la investigación son diferentes y se conoce que las isoenzimas son marcadores de expresión génica por lo que están influidas por las condiciones ambientales.
Varios autores informan la utilidad de los marcadores bioquímicos en el análisis de especies que resultan imposibles de diferenciar por sus características morfológicas. Tal es el caso de Astragalus gossypinus Fisch y Astragalus persicus Fisch. and C.A.Mey de la familia Fabaceae, donde los sistemas superóxido dismutasa, esterasas y peroxidasas tuvieron importante contribución por su alto polimorfismo (27). Igualmente, han sido aplicados en la identificación de especies silvestres diploides de trigo (Triticum boeoticum Bioss. y Triticum uratum Thumanian ex Gandilyan), con el empleo de las isoenzimas peroxidasas, polifenoloxidasas y superóxido dismutasa. Estos sistemas tienen un rol muy importante en los mecanismos de defensa de las plantas, además de considerarse estables en la expresión enzimática y son confiables para los estudios genéticos (28).
Los resultados publicados de diferentes investigaciones afines confirman la eficiencia de los sistemas peroxidasas, polifenoloxidasas, esterasas y anhidrasa carbónica en la identificación de especies en varios géneros como Nicotiana (29), Eucaliptus (30), Swietenia (31) y Musa (20). Mientras que al analizar 11 especies mexicanas de Datura (Solanaceae) los sistemas superóxido dismutasa y diaforasas fueron los más importantes (32).
Finalmente, al analizar los resultados del dendrograma, se puede apreciar que la formación de los grupos está en correspondencia con la categoría taxonómica de subgénero, pues en el grupo I se in-cluyen todas las especies pertenecientes al subgénero Petunioides y el grupo II lo integra la única especie estudiada perteneciente al subgénero Rustica (4). Sin embargo, en el caso de los subgrupos formados dentro del grupo I las especies que lo componen no se corresponden con la categoría de sección; ya que las dos especies pertenecientes a la sección Alatae se separan en subgrupos diferentes y se relacionan a miembros de otras secciones. De esta forma, N. longiflora se une a N. trigonophylla de la sección Trigonophyllae en el subgrupo I (a), aún cuando estas especies no coinciden en su comportamiento frente al moho azul y tienen diferente número cromosómico, 2n=2x=20 y 2n=2x=24, respectivamente. Por otra parte, N. alata se une a la especie N. glutinosa de la sección Undulatae para formar el subgrupo I(c), aunque ambas coinciden en ser susceptibles al moho azul y tienen números cromosómicos diferentes, 2n=2x=18 y 2n=2x=24, respectivamente.
Sin embargo, todas las especies estudiadas pertenecientes a la sección Suaveolentes integran el subgrupo I(b), coinciden en ser resistentes al moho azul y de origen australiano. Esta sección se caracteriza porque presenta un números cromosómicos diferentes, N. suaveolens y N. velutina poseen 2n=2x=32, N. ingulba y N. megalosiphon 2n=2x=40 y el anfidiploide N. debneyi 2n=4x=48 cromosomas. Se plantea que esta variación dentro de la sección pudo deberse a que la misma se originó a partir de un ancestro anfidiploide con 2n=4x=48 y que producto de sucesivos eventos de aneuploidía y reordenamientos en el complemento cromosómico pudieron haber evolucionado las distintas especies con diferente número de cromosomas (8). La especie N. knightiana la única que integra el grupo II es susceptible al moho azul y con número cromosómico 2n=2x=24.
Los resultados obtenidos confirman la validez y utilidad de la aplicación de los marcadores bioquímicos al permitir la caracterización de las especies estudiadas y agruparlas de acuerdo a sus afinidades genéticas. Esta información puede ser utilizada en los programas de mejoramiento genético de la especie comercial N. tabacum. para planificar cruzamientos inter específicos que posibiliten ampliar la variabilidad genética de la misma. Es de destacar que no se ha encontrado referencia alguna de estudios isoenzimáticos desarrollados en especies silvestres del género Nicotiana en Cuba, por lo que esta investigación podría considerarse como la primera que se desarrolla en tal sentido.
A pesar de la utilidad demostrada de los marcadores bioquímicos para la caracterización de la variabiabilidad genética, hay que tener en cuenta las limitaciones de los mismos. Las isoenzimas presentan ciertas desventajas para detectar todas las posibles diferencias a nivel de ADN. Esto se debe a las modificaciones que sufren los ARN en los procesos post-transcripcionales, la redundancia del código genético y al hecho de que no todas las sustituciones de aminoácidos provocan alteraciones en las movilidades electroforéticas (17). La técnica también presenta la limitante de ser sólo válida para el estudio de genes estructurales que codifican para proteínas solubles; además de que la actividad enzimática depende de la edad, el tipo de tejido utilizado y la conservación del mismo (33). Por todo ello es recomendable complementar los estudios bioquímicos con análisis mediante marcadores moleculares de ADN y, de esta manera, profundizar en el conocimiento de la variabilidad genética existente entre las especies del género Nicotiana.
REFERENCIAS
1. Hidalgo R. Variabilidad genética y caracterización de especies vegetales. Boletín Técnico No. 8. Instituto Nacional de Recursos Fitogenéticos (IPGRI), Cali, Colombia. 2003; 2-26.
2. Lewis R, Nicholson J. Aspects of the evolution of Nicotiana tabacum L. and the status of United State Nicotiana germoplasm collection. Genet Resour Crop Evol. 2007;(54):727-740.
3. Goodspeed TH. The genus Nicotiana. Chronica Botanica, Waltman, Mass; 1954.
4. Knapp S, Chase MW, Clarkson JJ. Nomenclatural changes and a new sectional classification in Nicotiana (Solanaceae). Taxon. 2004;(52):73-82.
5. Siva-Raju K, Sheshumadhav M, Murthy TGK. Molecular diversity in the genus Nicotiana as revealed by randomly amplified polymorphic DNA. Physiol Mol Biol Plant. 2008;14(4):377-382.
6. Pérez E. Caracterización morfoagronómica, citogenética e isoenzimática de ocho variedades de tabaco negro (Nicotiana spp.) de interés económico. Tesis de Maestría. Facultad de Biología. Universidad de La Habana; 2002.
7. Rodríguez L. Análisis del comportamiento de siete variedades de tabaco cubano (N. tabacum L.) ante el estrés biótico provocado por Peronospora hyosciamy f.sp. tabacina. Tesis de Diploma. Facultad de Biología. Universidad de La Habana; 2006.
8. Khan MQ, Narayan RKJ. Phylogenetic diversity and relationships among species of genus Nicotiana using RAPDs analysis. Afr J Biotechnol. 2007; 6(2):148-162.
9. Zhang HY, Liu XZ, He CS, Yang YM. Genetic diversity among flue-cured tobacco cultivars based on RAPD and AFLP markers. Braz Arch Biol Technol. 2008; 51(6):1097-1101.
10. Zhang HY, Liu XZ, He CS, Zheng CM. Random amplified DNA polymorphism of Nicotiana tabacum L. cultivars. Biol Plant. 2005;49:605-607.
11. Milla R, Levin JS, Lewis RS, Rufty RC. RAPD and SCAR markers linked to an introgressed gene conditioning resistance to Peronospora tabacina D.B. Adam. in tobacco. Crop Sci. 2005;(45):23462354.
12. Li BC, Bass WT, Cornelius PL. Resistance to tobacco black shank in Nicotiana species. Crop Sci. 2006;(46):554-560.
13. Julio E, Verrier J-L, Dorlhac de Borne F. Development of SCAR markers linked to three disease resistances based on AFLP within Nicotiana tabacum L. Theor Appl Genet 2006; (112):335-346.
14. Milián MD. Caracterización de la variabilidad genética en la colección cubana del género Xanthosoma. Tesis en opción al grado científico de Doctor en Ciencias Biológicas, Cuba; 2008.
15. Valdés-Infante J. Utilización de marcadores morfoagronómicos y marcadores de ADN para el desarrollo de una metodología que contribuya al mejoramiento genético del guayabo (Psidium guajava L.) en Cuba. Tesis en opción al grado científico de Doctor en Ciencias Biológicas, Cuba; 2009.
16. Ruíz C. Desarrollo de marcadores moleculares de aplicación en genómica y programas de mejora de cítricos. Tesis doctoral, Universidad de Valencia, Facultad de Ciencias Biológicas, España; 2002.
17. González, C. Detección del polimorfismo genética mediante marcadores bioquímicos en plantas. En: Marcadores Moleculares, nuevos horizontes en la genética y la selección de plantas. Cornide MT, (Ed) Felix Varela, La Habana; 2002. p.36-66.
18. Picca A, Helguera M, Salomón N, Carrera A. Marcadores moleculares. Capítulo 4. 2006; p.61-68. (En línea). Disponible en: (http://www.argenbio.org. (Consultado: 2 jun 2008).
19. Espino E. Guía para el cultivo del tabaco. (Ed) AGROINFO. Ministerio de la Agricultura, La Habana; 2009. p.17-46.
20. Román MI. Estudio de la diversidad genética en el género Musa en Cuba. Tesis en opción al grado científico de Doctor en Ciencias Biológicas, Cuba; 2004.
21. Wendel JF, Weeden NF. Visualization and interpretation of plant isozymes. In: Soltis DE, Soltis PS, (editors). Isozymes in Plant Biology. Discordies Press, Portland. Oregon.1989. p. 14.
22. Sigarroa A, Cornide M.T. Manual MAT-GEN. Manual del usuario. Folleto, Facultad de Biología, 1995; 37pp.
23. Rohlf FJ. NTSYS-PC, numerical taxonomy and multivariate analysis system. Version 2.1. 2001. Exeter Software, Setauket, New York. (En línea). Disponible en: http://www.exetersoftware.com/cat/ntsyspc.htlm. (Consultado: 4 Nov 2007).
24. Dice LR. Measures of the amount of ecologic association between species. Ecology. 1945; (26):297-302.
25. Azad AK, Jones JG, Haq N. Assessing morphological and isozyme variation of jackfruit (Artocarpus heterophyllus Lam.) in Bangladesh. Agroforest Syst. 2007;(71):109-125.
26. Smith HH, Hamill DE, Weaver EA, Thompson KH. Multiple molecular forms of peroxidases and esterases in species and amphiploids of Nicotiana. Isozyme Bull. 1970, 3:35-36.
27. Zarre S, Khodaei Z, Karamali Z, Niknam V, Mirmasoumi M. Isoenzyme variation patterns and species concept in Astragalus gossypinus and Astragalus persicus complexes (Fabaceae) in Iran. Biochem Syst Ecol. 2007; (35):757-763.
28. Cheniany M, Ebrahimzadeh H, Salimi A, Niknam V. Isozyme variation in some populations of wild diploid wheats in Iran. Biochem Syst Ecol. 2007; (35):363-371.
29. Valdés M, González C, Xiqués X, García H, Román MI, Rojas A. Perfiles isoenzimáticos para la caracterización de progenitores e híbridos en Nicotiana spp. Revista Cubana de Química 2004; XVI(3):588-602.
30. Geada D. Variabilidad isoenzimática del género Eucalyptus en la región Occidental de Cuba. Tesis de Maestría. Facultad de Biología. Universidad de La Habana, 2003.
31. González C, Geada D, Valdés M, Román MI, Xiqués X, García M. Relación filogenética entre dos especies del género Swietenia y su híbrido mediante estudios isoenzimáticos. Rev Jard Bot Nac. 2005; XXV- XXVI:175-178.
32. Jiao M, Luna-Cavazos M, Bye R. Allozyme variation in Mexican species and classification of Datura (Solanaceae). Plant Syst Evol. 2002; 232:155–166.
33. Solis L, Andrade A. ¿Qué son los marcadores moleculares? La ciencia y el hombre. Universidad Veracruzana. Rev Divul Cient Tecn. 2005; XIII (1):1-5.
(Recibido 4-3-2010; Aceptado 17-5-2010)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}