










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión impresa ISSN 1010-2752versión On-line ISSN 2224-4697
Rev. Protección Veg. v.25 n.3 La Habana sep.-dic. 2010
TRABAJO ORIGINAL
EFECTO DE LA EXPRESIÓN DEL GEN QUE CODIFICA PARA LA PROTEÍNA DE TRANSFERENCIA DE LÍPIDOS GBII10 EN LA RESISTENCIA DE Nicotiana tabacum VAR. HAVANA PETIT SR1 A Peronospora hyoscyami f. sp. tabacina
EXPRESSION EFFECT OF GENE ENCODING LIPID TRANSFER PROTEIN GBII10 ON THE RESISTANCE OF Nicotiana tabacum var. HAVANA PETIT SR1 TO Peronospora hyoscyami f. sp. tabacina
O. Chacón*, R. Portieles**, I. Hernández**, Y. López**, O. Borrás**
*Laboratorio de Fitopatología, Instituto de Investigaciones del Tabaco (IIT), San Antonio de los Baños, La Habana, Cuba. Correo electrónico: osmani@iitabaco.co.cu; **Centro de Ingeniería Genética y Biotecnología. Ciudad de La Habana, Cuba
RESUMEN
El tabaco (Nicotiana tabacum) es un cultivo de importancia económica para Cuba. Su rendimiento y calidad pueden afectarse por el ataque de microorganismos que le producen enfermedades y entre los más importantes se encuentra el oomycete Peronospora hyoscyami f. sp. tabacina, agente causal del moho azul del tabaco. La especie Nicotiana megalosiphon es altamente resistente a este patógeno y entre sus mecanismos de defensa se encuentra la expresión de genes que codifican para proteínas de transferencia de lípidos (LTP). Con el objetivo de evaluar el efecto de las LTP sobre la resistencia de la especie N. tabacum a esta enfermedad, se determinó la expresión constitutiva de la LTP GBII10 obtenida de N. megalosiphon en plantas de la variedad susceptible de N. tabacum Havana Petit SR1. Como resultado se obtuvo un grupo de plantas transgénicas con un incremento de la resistencia a la enfermedad, lo que demuestra el efecto inhibitorio de la LTP GBII10 en el desarrollo del patógeno. Se sugiere utilizar este gen como marcador de selección de genotipos resistentes al moho azul del tabaco y en la obtención de transgénicos en otros cultivos de importancia económica, así como evaluar su efecto inhibitorio sobre otros patógenos.
Palabras clave: tabaco; proteína de transferencia de lípidos, mecanismos de defensa, Peronospora hyoscyami f. sp. tabacina
ABSTRACT
Tobacco (Nicotiana tabacum) is an economically important crop in Cuba. Its yield and quality can be affected by several pathogenic microorganisms. The oomycete Peronospora hyoscyami f. sp. tabacina is the causal agent of the tobacco blue mold, which is the most important disease of this crop,. The noncultivated tobacco species Nicotiana megalosiphon has been shown to be highly resistant to P. hyoscyami f. sp. tabacina. This pathogen induces the Lipid Transfer Protein (LTP) gene expression as a defense mechanism of the plant following infection. In order to evaluate the LTP effect on the N. tabacum resistance to blue mold disease, LTP GBII10 isolated from N. megalosiphon was expressed in susceptible plants of N. tabacum var. Havana Petit SR1. Constitutive expression of LTP GBII10 gene in transgenic tobacco enhanced resistance to the disease showing an inhibitory effect in the development of P. hyoscyami f. sp. tabacina. LTP GBII10 is suggested to be used in the marker assisted selection of tobacco resistant genotypes and in obtaining transgenic plants in other economically important crops. Also, the inhibitory effect of LTP GBII10 against plant pathogenic microorganisms could be evaluated.
Key words: tobacco, Lipid Transfer Proteins, defense mechanisms, Peronospora hyoscyami f. sp. tabacina
(Recibido 22-9-2009; Aceptado 19-5-2010)
INTRODUCCIÓN
El tabaco (Nicotiana tabacum L.) pertenece a la familia de las solanáceas. Es un cultivo de gran importancia económica para Cuba, con plantaciones que se extienden prácticamente a todas las provincias, en un estimado de 34 000 ha de tierras cultivables (1).
El rendimiento y la calidad de este cultivo puede verse afectado por diferentes enfermedades, entre ellas el Moho Azul del Tabaco (2,3,4,5). Su agente causal es el oomycete Peronospora hyoscyami f. sp. tabacina Skalický (5), que pertenece al reino Cromista (5,6,7,8,9,10,). Este patógeno es un biótrofo, con un ciclo de la enfermedad que oscila entre los siete y 10 días (2) y presenta una extrema movilidad, ya que sus esporas pueden viajar por el aire hasta 5000 km de distancia (11).
Las plantas de manera natural responden al ataque de patógenos a través de la activación de una variedad de mecanismos protectores, que evitan la replicación del patógeno así como su dispersión (12).
Entre las principales respuesta defensivas de N. tabacum a P. hyoscyami f. sp. tabacina, descritas hasta la fecha, se encuentra el efecto inhibitorio del terpenoide b-ionone sobre los esporangiósforos y las esporangiosporas de P. hyoscyami f. sp. tabacina (13,14). También, se identificó un grupo de proteínas denominadas T-Filoplaninas, secretadas por los tricomas de la superficie de las hojas de la planta, que inhiben la germinación de este oomycete y por tanto impide el desarrollo de la enfermedad (15).
Recientemente, en plantas de la especie resistente N. megalosiphon Van Heurck y Muell. Arg. inoculadas con P. hyoscyami f. sp. tabacina se identificó el gen GBII10 que codifica para la proteína de transferencia de lípidos (LTP) (16). Esta pertenece a una familia de proteínas pequeñas de 9 y 10 kDa, que contienen ocho residuos de cisteína con cuatro puentes disulfuros conservados (17) y se incluyen en el grupo PR-14 de las proteínas relacionadas con la patogénesis (PR) (18).
Las LTP se distribuyen en forma abundante en las membranas y paredes celulares y se expresan en respuesta a la infección por patógeno, lo que sugiere que son proteínas activas de la defensa de las plantas (19). Además, pueden inhibir el crecimiento de patógenos fungosos in vitro y son capaces de incrementar sinérgicamente las propiedades antimicrobianas de otros péptidos antimicrobianos como las defensinas y tioninas (20).
Teniendo en consideración la importancia de la enfermedad Moho Azul del Tabaco, así como la función de las LTP en la defensa de las plantas, el objetivo de este trabajo fue evaluar el efecto de la expresión constitutiva de la LTP GBII10 en plantas de la variedad susceptible de N. tabacum Havana Petit SR1 sobre la resistencia a P. hyoscyami f. sp. tabacina.
MATERIALES Y MÉTODOS
Material vegetal
Las semillas de plantas de tabaco, pertenecientes al género Nicotiana con diferente grado de resistencia a P. hyoscyami f. sp. tabacina: Sumatra 104 y Petit Havana SR1 (susceptibles) y N. megalosiphon (resistente), se utilizaron para la obtención de posturas, a partir de la técnica de semilleros en bandejas flotantes (21), con sustrato conformado por cáscara de arroz y turba negra, en una proporción (4:1). A los 30 días de edad, las posturas se transplantaron a macetas de seis pulgadas que contenían 1900g del mismo sustrato y se mantuvieron en casas de cultivo a 23°C, con una humedad relativa del 90% y períodos de luz y oscuridad de aproximadamente 12 horas, durante 15 días (22).
Construcción del vector de expresión y transformación de las plantas de tabaco
El ADNc correspondiente al gen que codifica para la proteína de transferencia de lípidos (LTP) GBII10, descrito por Borrás-Hidalgo et al. (16), se clonó en el vector de transformación pCambia 2300 para obtener plasmidios binarios (CAMBIA, Australia).
La transformación genética de plantas de N. tabacum var. Petit Havana SR1 se realizó según la metodología de Zambryski et al. (23). Para ello, se transformó la cepa GV2260 de Agrobacterium tumefaciens Smith y Townsend por el método del nitrógeno líquido (24) con los plasmidios binarios desarrollados. Seguidamente, se transformaron discos de hojas de plantas cultivadas in vitro, utilizando kanamicina (100 mg.L-1) como marcador de selección. Los discos de hojas se cocultivaron con los clones de A. tumefaciens recombinantes por 48 horas en medio MS líquido (25). La regeneración de los brotes de plantas (4-6 semanas) se realizó en el medio MS de regeneración y el enraizamiento de los brotes de plantas (1-3 semanas) se efectuó en el medio MS de enraizamiento. Las plantas enraizadas, con resistencia a kanamicina y con el gen LTP, se transplantaron a macetas para su adaptación en casas verdes durante 45 días.
Evaluación de la expresión del gen LTP GBII10 en plantas transformadas mediante la reacción en cadena de la polimerasa (PCR)
Se purificó el ADN genómico de las hojas de plantas transformadas y no transformadas, mediante un juego de reactivos de Promega (Genomic DNA Purification Kit). La PCR de los clones se realizó según la metodología de Sambroock et al. (26) y se utilizó la enzima Taq Pwo DNA Polimerasa (Heber Biotec).
Los cebadores utilizadas fueron GBII10F (5'CGTATCTCGAGCATGCTGC3') y GBII10R (5'GTGGTCGCTGCGATGCTAGC3'). Las condiciones para la PCR fueron de 35 ciclos a 95°C por 1 min, 53°C por 1 min y 72°C por 1 min, seguido por 5 min a 72°C. El resultado de la reacción se visualizó mediante la tinción con bromuro de etidio, sobre un gel de agarosa al 1%, expuesto a la luz ultravioleta y para la corrida electroforética se utilizó una solución de 1xTBE, un voltaje de 110 volt, durante un tiempo de 30 min.
Inoculación con P. hyoscyami f. sp. tabacina
Se utilizó el aislamiento IT11 de P. hyoscyami f. sp. tabacina, suministrado por Instituto de Investigaciones de Sanidad Vegetal. Se obtuvo originalmente de parcelas experimentales pertenecientes al Instituto de Investigaciones del Tabaco, de la provincia La Habana en la campaña de 2007. Se conservó en plantas de N. tabacum var. Criollo 98, susceptible a la enfermedad y se homogenizó (27).
Se inocularon un total de 70 clones transgénicos, 70 plantas de N. tabacum var. Havana Petit SR1 (susceptible a la enfermedad), 20 plantas de N. tabacum var. Sumatra (altamente susceptible a la enfermedad) y 20 plantas de la especie N. megalosiphon (resistente a la enfermedad).
Para la inoculación de las plantas se colocaron en un grupo de cinco a seis hojas jóvenes ocho gotas de 10mL de una suspensión con una concentración de 5 x 103 esporas.mL-1, determinada con un hemacitómetro. Las plantas se taparon con bolsas plásticas negras húmedas durante un período de 12 horas para promover la infección y seguidamente se sometieron a fotoperíodos de 12 horas de luz y 12 horas de oscuridad. Se utilizaron como control plantas sanas inoculadas con agua.
Las plantas inoculadas con el patógeno y los controles se mantuvieron a una temperatura de 23°C. La evaluación de los síntomas se realizó 10 días después de la inoculación según la metodología recomendada por Echiltz (28).
RESULTADOS Y DISCUSIÓN
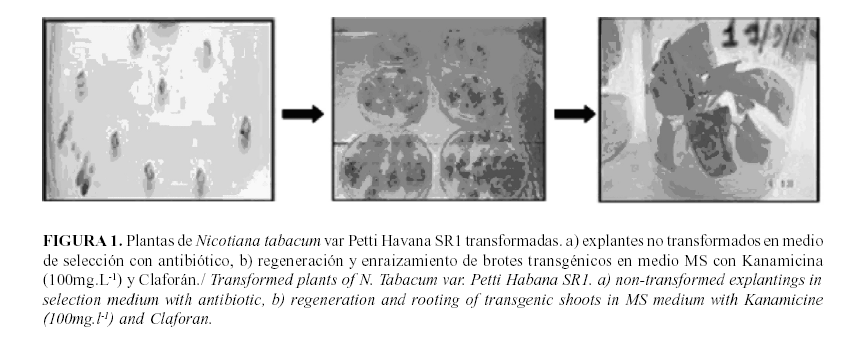
Como resultado de la transformación de las plantas de N. tabacum var. Petit Havana SR1 con el gen LTP, se obtuvo un 38,9% de eficiencia de transformación de brotes transgénicos en el medio de selección. De 351 brotes, 140 enraizaron en el medio con el antibiótico (Fig. 1). Es importante señalar que las plantas transformadas con la LTP no presentaron variaciones fenotípicas con respecto a las plantas no transformadas.
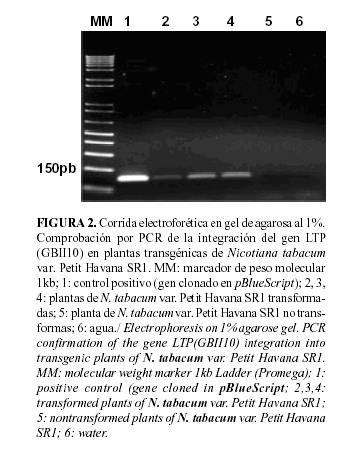
Cuando se obtiene una planta transgénica, un paso esencial es su caracterización molecular. Mediante PCR con cebadores específicos, que amplifican solamente el péptido maduro (150pb), se pudo comprobar la integración del gen LTP GBII10 en el genoma de las plantas de N. tabacum var. Petit Havana SR1 y el éxito de la transformación.
Como resultado de la amplificación, se observó una banda a la altura de 150 pares de bases en el ADN proveniente de las hojas de plantas transformadas, coincidiendo con la talla de la banda amplificada en el control positivo (gen LTP GBII10 clonado en pBluescript). En los controles negativos empleados (ADN de N. tabacum var. Petit Havana SR1 no transformadas y H2O) no hubo amplificación (Fig. 2).
En N. tabacum, el ciclo de infección comienza con la germinación de las esporangiosporas y la penetración del hospedante en aproximadamente dos horas y media, seguido de la invasión del espacio intercelular, alrededor de cuatro horas. El primer haustorio se forma seis horas después, cuando la hifa entra en contacto con las células del mesófilo (29,30). Las variedades de N. tabacum: Sumatra 104 y Petit Havana no transformada, se comportan de manera susceptible frente a P. hyoscyami f. sp. tabacina, por lo que se puede esperar un tiempo similar de ocurrencia de los eventos de infección.
En el patosistema establecido entre N. megalosiphon y P. hyoscyami f. sp. tabacina, las plantas no mostraron los síntomas de la enfermedad Moho Azul (Fig. 3). Esto indica claramente que la infección producida por este biótrofo resultó en una interacción planta-patógeno del tipo incompatible.
La expresión constitutiva del gen que codifica para la proteína de transferencia de lípidos LTP GBII10 en plantas transgénicas de N. tabacum var. Petit Havana SR1 (Fig. 3), trajo consigo un incremento de la resistencia de estas plantas a la infección con el patógeno.
Sin embargo, el silenciamiento de la LTP GBII10 en plantas de N. megalosiphon mediante vectores virales que inducen silenciamiento génico (VIGS), no afecta el grado de resistencia de esta especie al patógeno (16).
Esto indica que es probable que la proteína codificada por este gen, forma parte de los mecanismos de defensa no específicos de estas plantas y que elevados niveles de transcriptos correspondientes a este gen, pueden tener un efecto negativo para el desarrollo del patógeno. Las LTP son proteínas básicas de 9kDa que en las plantas constituyen el 4% de la proteína total. Ellas son capaces de transferir lípidos entre las membranas in vitro y en las plantas participan en varios procesos fisiológicos, incluyendo la resistencia a enfermedades (31).
En plantas de N. tabacum susceptibles, la sobreexpresión de las LTP CALTP1 y CALTP2, incrementó la resistencia al oomycete Phytophthora nicotianae Breda de Haan y a la bacteria Pseudomonas syringae pv. tabaci (Wolf y Foster; Young et al.). En este caso el aumento en la resistencia fue dependiente del nivel de transcripto (32).
Se obtuvieron un total de 70 clones transgénicos que presentaron integrado en su genoma el fragmento de 150 pb correspondiente al gen. Del total de clones, 68 expresaron el gen LTP GBII10 y mostraron el grado de resistencia a la enfermedad moho azul, correspondiente a la categoría resistente (28), 10 días después de la inoculación con el patógeno. Estos tuvieron igual comportamiento que el control resistente (N. megalosiphon), a diferencia de los controles susceptibles (N. tabacum var. Petit Havana SR1 no transformada y N. tabacum var. Sumatra), los cuales reflejaron los síntomas de manchas amarillas y necróticas, típicos de la enfermedad, correspondientes a las categorías susceptible y altamente susceptible respectivamente (28) (Fig. 3).
Estudios recientes demostraron el potencial antifúngico de las LTP. Se observó que plantas transgénicas de arroz que contienen el gen de la LTP Ace-AMP1 de cebolla (Allium cepa L.) mostraron actividad antimicrobiana hacia Magnaporte grisea (T.T. Hebert) M.E. Barr, Rhizoctonia solani Kühn y Xanthomonas oryzae sp. oryzae (Fang et al.) (33). En este mismo cultivo, la expresión de la LTP110 inhibe la germinación de las esporas de Pyricularia oryzae Cav. (34). También plantas transgénicas de trigo (Triticum aestivum L.) mostraron un incremento de la actividad antifúngica contra Blumeria graminis f. sp. tritici (DC.) Speer (31).
Adicionalmente, investigaciones con plantas de trigo (T. aestivum) revelaron que la sobreexpresión transiente de la LTP1, reduce la eficiencia de la penetración por el mildiu polvoriento (B. graminis f.sp. tritici) en un cultivar susceptible, en alrededor de un 28,3%. Esto indica el posible papel inhibidor de esta proteína sobre las estructuras del patógeno para realizar esta función (31).
Finalmente, varios autores señalan, que en plantas transgénicas de tabaco y de Arabidopsis thaliana (L.) Heynh., la expresión de la LTP2 de fríjol (Phaseolus vulgaris L.), incrementó la tolerancia a bacterias patógenas (35,36), lo cual es debido a las propiedades antibióticas que poseen las LTP (37).
Se ha informado que no todas las PR-proteínas son antimicrobianas, por lo que la identificación y el aislamiento de proteínas con estas características es importante para su uso en el desarrollo de plantas transgénicas con resistencia a patógenos. No obstante esta es una estrategia que no se realiza en el cultivo del tabaco en Cuba, pues las leyes del país prohíben elaborar cigarros y cigarrillos a partir de variedades transgénicas de N. tabacum. Por lo tanto, la LTP GBII10 obtenida de la especie N. megalosiphon pudiera ser utilizada en la obtención de transgénicos en otras solanáceas de importancia económica, que son afectadas por varios patógenos.
La expresión constitutiva de la LTP GBII10 en N. tabacum var. Petit Havana SR1 no afectó el fenotipo de las plantas transformadas e incrementó la resistencia de las mismas a la enfermedad moho azul. Por lo tanto, se recomienda determinar si este gen pudiera ser utilizado como marcador molecular para la selección de genotipos resistentes a esta enfermedad, así como evaluar el efecto inhibidor de la proteína codificada por este gen, sobre otros microorganismos patógenos, con el fin de crear las bases para el desarrollo de un producto biológico.
REFERENCIAS
1. Morán YM, Ramos PL, Domínguez M, Fuentes AD, Sánchez Y, Crespo JA. Tobacco leaf curl Cuba virus, a new begomovirus infecting tobacco (Nicotiana tabacum) in Cuba. Plant Pathol. 2006;55:570.
2. Lucas GB. The war against blue mold. Science. 1980;210:147-153.
3. Main CE. Blue Mold. In Compendium of Tobacco Diseases. Shew HD, Lucas GB, eds. American Phytopathological Society, St. Paul, MN. 1998; p.5-9.
4. Ristaino JB, Johnson A, Blanco-Meneses M, Liu B. Identification of the tobacco blue mold pathogen, Peronospora tabacina, by polymerase chain reaction. Plant Dis. 2007;91:685-691.
5. Voglmayr H. Progress and challenges in systematics of downy mildews and white blister rusts: new insights from genes and morphology. Eur J Plant Pathol. 2008;122:3-18.
6. Cavalier-Smith T. The kingdom Chromista. In: Geen JC, Leadbeater BSC, Diver WL, editors. The Chromophyte Algae: Problem and Perspective. Syst. Assoc. Spec. Oxford, UK. 1989;38:381-407.
7. Judelson HS, Tyler B, Michelmore MRW. Regulatory sequences for expressing genes in oomycetes fugi. Mol Gen Genet. 1992;234:138-146.
8. Dick MW. The straminipilous fungi: A new classification for biflagellate fungi and their uniflagellate relatives with particular reference to Lagnidiaceous fungi. C. A. B. Int. Mycol. Pap. 168; 1995.
9. Erwin DC, Ribeiro OK. Phytophthora Diseases Worldwide. St. Paul, MN: American Phytopathological Society. 1996; 592p
10.van de Peer Y, De Wachter R. Evolutionary relationships among the eukaryotic crown taxa taking into account site-to-site rate variation in 18S rRNA. J Mol Evol. 1997;45:619-630.
11.Main CE. The Blue Mold Disease of Tobacco. Blue Mold Forecast Homepage, 1997.
12.Malolepsza U, Rózalaska S. Nitric oxide and hydrogen peroxide in tomato resistance. Nitric oxide modulates hydrogen peroxide level in O-hydroxyethylorutin-induced resistance to Botrytis cinerea in tomato. Plant Physiol Biochem. 2005;43:623-635.
13.Schiltz P. Action inhibitrice de la b-ionone au cours du développement de Peronospora tabacina. Ann Tabac. 1974;11:207-216.
14.Salt SD, Tuzun S, Kuc J. Effect of b-ionone and abscisic acid on the growth of tobacco and resistance to blue mold: Mimicry of effects of stem infection by Peronospora tabacina Adam. Physiol Mol Plant Pathol. 1986;28:287-297.
15.Kroumova AB, Shepherd RW, Wagner GJ. Impacts of T-phylloplanin gene knockdown and of Helianthus and Datura Phylloplanins on Peronospora tabacina spore germination and disease potential. Plant Physiology. 2007;144:1843-1851.
16.Borrás-Hidalgo O, Thomma BPHJ, Collazo C, Chacón O, Borroto CJ, Ayra C, et al. EIL2 transcription factor and glutathione synthetase are required for defense of tobacco against tobacco blue mold. Mol. Plant Microbe Interact. 2006;19:399-406.
17.Lee JY, Kyeongsik M, Hoon Chaa D, Hae Sh, Kwang YH, Se WS. Rice non-specific lipid transfer protein: the 1.6 A crystal structure in the unliganded state reveals a small hydrophobic cavity. J Mol Biol. 1998; 276:437-448.
18.van Loon LC, Rep M, Pietersel CMJ. Significance of inducible defense related proteins in infected plants. Annu Rev Phytopathol. 2006;44:135-162.
19.Blein JP, The´venot PC, Marion D, Ponchet M. From elicitins to lipid-transfer proteins: a new insight in cell signaling involved in plant defense mechanisms. Trends Plant Sci. 2002;7:293-296.
20.Marion D, Douliez JP, Gautier MF, Elmorjani K. Plant lipid transfer proteins: relationships between allergenicity and structural, biological and technological properties. En: Mills ENC, Shewry PR. (Eds.). Plant Food Allergens, Blackwell Science, Oxford. 2004; p.57-69.
21.Espino E, Andino V, Quintana G. Instructivo Técnico para el cultivo del tabaco. Editorial SEDAGRI/AGRINFOR. Ministerio de la Agricultura. 1988; p. 67-80
22.Pérez E, Fernández A, Andino V. Tecnología para la eliminación del bromuro de metilo. Instituto de Sanidad Vegetal, MINAGRI, Ciudad de La Habana. 2000; 50p.
23.Zambryski P, Joos H, Gentello J, Leemans J, Van Montagu M, Schell J. Ti-plasmid vector for introduction of DNA into plant cells without altering their normal regeneration capacity. EMBO J. 1983;2:2143-2150.
24.Hofgen R, Willmitzer L. Storage of competent cells for Agrobacterium transformation. Nucl Acids Res. 1988;16:9877.
25.Murashige T, Skoog F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Plant Physiol. 1962;15:173-497.
26.Sambroock J, Fritsch EF, Maniatis T. Molecular cloning: A laboratory manual 2nd ed. New York: Cold Spring Harbor. 3; 1989.
27.Reuveni M, Nesmith WC, Siegel MR. Symptom development and disease severity in Nicotiana tabacum and N. repanda caused by Peronospora tabacina. Plant Dis. 1986;70:727-729.
28.Schiltz P. Tentative d' amelioration de l'essai collectif destiné á déterminer le pouvoir phatogéne de P. tabacina. CORESTA. 1974;1:16-22.
29.McKeen WE, Svircev AM. Early development of Peronospora tabacina in the Nicotiana tabacum leaf. Can J Plant Pathol. 1981;3:145-158.
30.Svircev A, McKeen WE. The haustorium of Peronospora hyoscyami f.sp. tabacina in the susceptible tobacco leaf cell. Can J Plant Pathol. 1982;4:119-128.
31.Ai-li LI, Cheng-sheng M, Rong-hua Z, Zhi-ying MA, Ji-zeng JIA. Assessment of Lipid Transfer Protein (LTP1) Gene in Wheat Powdery Mildew Resistance. Agricultural Sciences in China. 2006;5(4):101-105.
32.Sarowar S, Kim YJ, Kim KD, Hwang BK, Ok SH, Shin JS. Over expression of lipid transfer protein (LTP) genes enhances resistance to plant pathogens and LTP functions in long-distance systemic signaling in tobacco. Plant Cell Rep. 2009;28:419-427.
33.Roy-Barman S, Sautter C, Chattoo BB. Expression of the lipid transfer protein Ace-AMP1 in transgenic wheat enhances antifungal activity and defense responses. Transgenic Res. 2006;15:435-446.
34.Ge X, Chen J, Li N, Lin Y, Sun C, Cao K. Resistance function of rice lipid transfer protein LTP110. J Biochem and Mol Biol. 2003;36:603-607.
35.Molina A, Garcia-Olmedo F. Enhanced tolerance to bacterial pathogens caused by the transgenic expression of barley lipid transfer protein LTP2. Plant J. 1997;12:669-675.
36.Carvalho AO, Machado OLT, Cunha MA, Santos IS, Gomes VM. Antimicrobial peptides and immunolocalization of a LTP in Vigna unguiculata seeds. Plant Physiol. and Biochem. 2001;39:137-146.
37.Garcia-Olmedo FG, Molina A, Segura A, Moreno M. The defensive role of nonspecific lipid transfer proteins in plants. Trends Microbiol. 1995;3:72-74.


{kind=link}
{kind=link}
{kind=link}