










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión On-line ISSN 2224-4697
Rev. Protección Veg. v.25 n.3 La Habana sep.-dic. 2010
IDENTIFICACIÓN DE AISLAMIENTOS AUTÓCTONOS DEPSEUDOMONAS FLUORESCENTES CON ACTIVIDAD ANTAGÓNICAANTE Curvularia spp.
IDENTIFICATION OF NATIVE ISOLATES OF FLUORESCENT PSEUDOMONAS WITH ANTAGONISTIC ACTIVITY AGAINST Curvularia spp.
Annia Hernández-Rodríguez*, D. León-Plasencia*, Narovis Rives- Rodríguez**, Acela Díaz-de la Osa*, M. Almaguer-Chávez*, Yanelis Acebo-Guerrero*
*Facultad de Biología. Universidad de la Habana. Calle 25 #455 e/J e I. Vedado, Ciudad de la Habana, Cuba. Correo electrónico: annia@fbio.uh.cu, acebo@fbio.uh.cu; **Instituto de Investigaciones de Granos. Km 16 ½, Autopista Novia del Mediodía, Bauta, La Habana, Cuba
RESUMEN
La identificación de rizobacterias con potencialidades para el control biológico y estimulación del crecimiento vegetal resulta de especial interés para el cultivo del arroz (Oryza sativa L.) en Cuba. Este trabajo tiene como objetivo identificar aislamientos autóctonos de pseudomonas fluorescentes con actividad antagónica ante Curvularia pallescens y Curvularia trifolii. Los resultados demostraron que todos los aislamientos tienen efecto antagónico ante los aislamientos fúngicos probados, mostrando porcentajes de inhibición desde 49% hasta un 83%. Las cepas bacterianas fueron incluidas dentro de las especies Pseudomonas putida (14 de ellas) y Pseudomonas fluorescens (tres). De modo general, se demuestra las potencialidades de estas rizobacterias como agentes de control biológico de patógenos fúngicos en el cultivo del arroz.
Palabras clave: PGPB; rizobacterias; pseudomonas; arroz
ABSTRACT
The identification of rhizobacteria with potential for the biocontrol and plant growth promotion is of great interest for the improvement of rice crop (Oryza sativa L.) in Cuba. This work was aimed at identifying native fluorescent pseudomonas isolates with antagonistic activity against Curvularia pallescens and Curvularia trifolii. The results showed that all the isolates selected had antagonistic effect against the fungal isolates tested, showing inhibition percentages ranging from 49% to 83%. They were classified as Pseudomonas putida (14 of them) and Pseudomonas fluorescens (three). In general, it suggests the potential of these rhizobacteria as biocontrol agents against fungal pathogens in rice crop.
Key words: PGPB; rhizobacteria; pseudomonas; rice
(Recibido 25-1-2010 ; Aceptado 10-5-2010)
INTRODUCCIÓN
El arroz (Oryza sativa L.) es una gramínea originaria del continente asiático, cultivada y consumida por los humanos desde hace más de 7000 años (1). Su producción y distribución se concentran principalmente en Asia, aunque no debe descartarse su consumo en el resto del mundo. En Cuba, el consumo de arroz per cápita asciende a 60 kg por persona anualmente (2).
En el cultivo del arroz, las enfermedades microbianas constituyen uno de los factores que inciden en la obtención de bajos rendimientos y manchado de los granos, destacándose las de origen fúngico por ser las de mayor frecuencia de aparición. La enfermedad conocida como «manchado del grano» del arroz o «pecky rice», causada por un complejo fúngico y bacteriano, se presenta en la mayor parte de las regiones productoras de arroz del mundo y en Cuba, Curvularia spp. es uno de los géneros involucrados en su aparición. Entre las principales especies de Curvularia que provocan afectaciones a los granos de arroz en Cuba se encuentran C. pallescens (Boedijn) y C. trifolii (Kuffman)(3).
Esta enfermedad causa pigmentación y reducción de la germinación (3), por lo que afecta los componentes del rendimiento y la calidad de los granos (4). Se ha demostrado que las bajas temperaturas, las precipitaciones continuas y una humedad relativa elevada en el momento de la floración y durante la maduración del grano son factores que influyen en su desarrollo (5).
Como parte de la agricultura sostenible, se trata de controlar estos patógenos y lograr altos rendimientos del cultivo mediante una combinación adecuada de fertilizantes químicos y productos biológicos. En este sentido, la aplicación de inoculantes bacterianos como las Bacterias Promotoras del Crecimiento Vegetal (PGPB) ha constituido una alternativa ecológica, que favorece la conservación del medio ambiente y el agroecosistema (6,7,8).
Diversos estudios han demostrado la eficacia del empleo de PGPB para controlar enfermedades en el cultivo del arroz (9,10,11,12,13). Las bacterias pertenecientes al grupo de las pseudomonas fluorescentes se encuentran entre las más utilizadas como antagonistas bacterianos porque tienen la capacidad de producir una amplia variedad de metabolitos con efecto antagónico ante patógenos en la rizosfera, y constituyen un excelente ejemplo de la combinación de múltiples mecanismos para ejercer un efectivo control biológico (14,15,16,17,18,19).
Las investigaciones que involucran cepas de rizobacterias autóctonas de los ecosistemas arroceros en estudio son de suma importancia, ya que se plan-tea que estas podrían tener una mayor eficiencia en el control biológico de patógenos presentes en las mismas condiciones naturales. Sin embargo, es importante seleccionar aislamientos utilizando como primer criterio de selección la actividad antagónica frente al patógeno de interés, para luego proceder a la identificación y caracterización de los más promisorios mediante el uso de la taxonomía polifásica. Este trabajo tiene como objetivo identificar aislamientos autóctonos de pseudomonas fluorescentes con actividad antagónica frente a C. pallescens y C. trifolii.
MATERIALES Y MÉTODOS
Origen y clasificación de los aislamientos: Se seleccionaron 17 aislamientos de pseudomonas fluorescentes procedentes de la rizosfera de la variedad de arroz J-104 (20) sobre suelo Gley Vértico Crómico-Nodular Ferruginoso (12) y que habían mostrado actividad antagónica ante Pyricularia grisea (Sacc.). Los mismos se encontraban conservados en medio Luria Broth (LB) (8) con glicerol (1:1) a 4° C en la Colección de Cultivos del Departamento de Microbiología y Virología de la Facultad de Biología de la Universidad de La Habana. Se utilizaron además las cepas de referencia Pseudomonas aeruginosa (Migula) 7NSK2 y Pseudomonas putida (Trev.) WCS358 procedentes de la Colección de Cultivos de la Universidad de Gante en Bélgica, y la cepa Pseudomonas fluorescens (Migula) J-143 proveniente del Cepario Nacional de Biofertilizantes, Facultad de Biología, Universidad de La Habana, Cuba. Para la identificación y clasificación de los aislamientos puros se realizaron determinaciones micromorfológicas y culturales en Medio King B (Merck), según lo informado por Bergey (21).
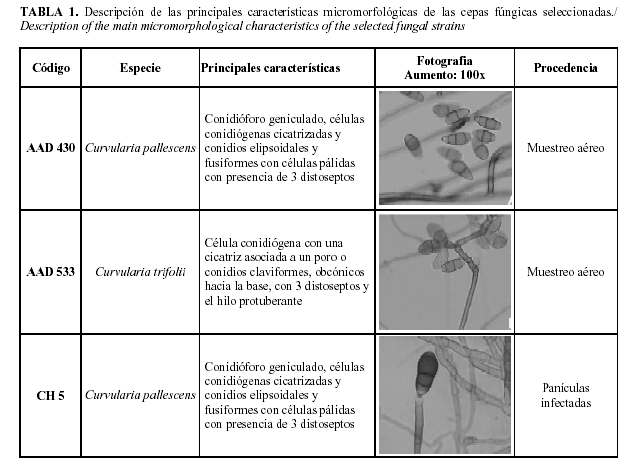
Actividad antagónica de los aislamientos frente a cepas de Curvularia spp.: Se realizó un bioensayo in vitro para determinar el efecto antagónico de los aislamientos bacterianos seleccionados frente a las tres cepas fúngicas: C. pallescens AAD 430, C. pallescens CH5 y C. trifolii AAD 533, previamente aisladas de un agroecosistema arrocero perteneciente al Instituto de Investigaciones de Granos, Bauta, La Habana, Cuba (22) (cuyas características micromorfológicas se recogen en la Tabla 1 (23). Las cepas fúngicas se encontraban conservadas en medio Papa Dextrosa Agar (PDA) (Biocen) a 4°C en la Colección de Cultivos Fúngicos de la Facultad de Biología de la Universidad de La Habana.
Para el desarrollo del experimento in vitro se siguió la metodología descrita por Hernández et al. (24). Se realizaron suspensiones bacterianas en solución salina fisiológica, ajustando la concentración celular a 108 células. mL-1, según la escala de McFarland. Se tomaron 100 µL de las suspensiones y se sembraron por diseminación en medio Agar King B. Los hongos se sembraron previamente en medio de cultivo King B (Merck) según lo informado por Trujillo et al. (26), y se incubaron a 30ºC durante 7 días (25). Las placas Petri previamente sembradas con el hongo se perforaron utilizando un obturador de 5 mm de diámetro.
Los discos se colocaron en el centro de las placas Petri con medio King B sembradas con los aislamientos seleccionados, a razón de un disco para cada placa y cinco réplicas por tratamiento, luego se incubaron a 30ºC durante 7 días. Se estableció un control donde no se aplicó el aislamiento bacteriano. La actividad antagónica de las bacterias se determinó a través de mediciones del diámetro de crecimiento del hongo en presencia de las bacterias. Con estos datos se determinó el porcentaje de inhibición de los aislamientos sobre los fitopatógenos, según metodología descrita por Hernández et al. (24). El experimento se repitió cinco veces.
Identificación de los aislamientos: Se realizó mediante la utilización del sistema API 20NE (bioMérieux SA), un micrométodo estandarizado que combina 8 pruebas convencionales y 12 pruebas de asimilación para la identificación de bacilos Gram negativos. Los resultados fueron interpretados mediante el software (API Web StandAlone®).
Extracción y amplificación del DNA: Para la extracción del DNA los aislamientos seleccionados fueron cultivados en medio LB (Merck) líquido durante 16 horas a 28°C (en agitación a 125 rpm). Luego se centrifugó el contenido de los tubos durante 10 minutos a 10000 rpm y el DNA se extrajo siguiendo la metodología descrita por Branwel et al. (27). Finalmente se procedió al secado del precipitado al vacío (Bomba de vacío Memmert), se resuspendió el producto en 150 mL de solución TE y se conservó a -200C para su posterior utilización.
La amplificación de la subunidad rRNA16S de los aislamientos de P. fluorescens y P. putida fue ejecutado por PCR. Para la identificación de P. fluorescens se utilizaron los cebadores:
16sPSEfluF (5'TGCATTCAAAACTGACTG3') y 16PSER (5'AATCACACCGTGGTAACCG3') (28).
Para P. putida se utilizó el cebador REPc (5'GTAGGAGCGGGTTTACCCGCGAA3') (29). Todos los cebadores se sintetizaron en el Centro de Ingeniería Genética y Biotecnología, La Habana, Cuba.
Las amplificaciones se realizaron en un equipo termociclador (Mastercycler, Eppendorf). Se utilizaron las mezclas de reacción y programas descritos por Scarpellini et al. (28) (P. fluorescens) y Aranda-Olmedo et al. (29) (P. putida). Como control negativo se utilizaron 20 µL de mezcla de reacción más 5 µL de agua destilada. Como control positivo se realizó la amplificación del ADN de las cepas de referencia y patrón P. fluorescens J-143 y P. putida WCS358.
Los productos de la PCR fueron analizados mediante electroforesis en gel de agarosa al 1,5 % en Tampón TBE 1X (0,09mM Tris-borato, 0,002 mM EDTA), teñido con una solución de bromuro de etidio (1 µg.mL-1), de acuerdo al procedimiento descrito por Sambrook et al. (30).
Análisis Biométricos: De forma preliminar, a todas las variables se le realizó la prueba de normalidad de Shapiro y Wilk, y homogeneidad de varianza. Los datos presentaron distribución normal, por lo que se les realizó la prueba de Tukey (paramétrica). Se utilizó el paquete estadístico Statistica (versión 6.1).
RESULTADOS
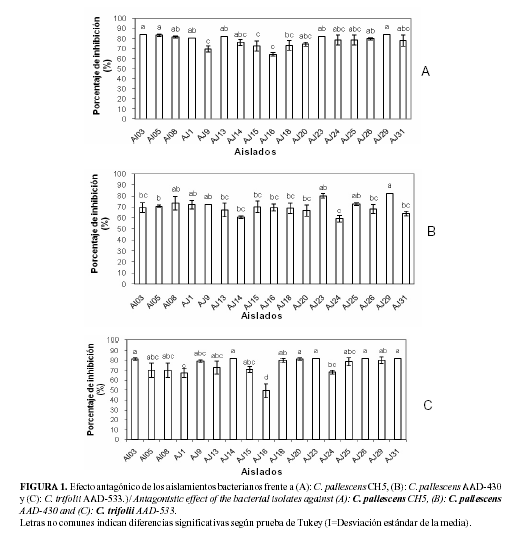
Actividad antagónica de los aislamientos frente a C. trifolii y C. pallescens: En el experimento in vitro los aislamientos con características típicas del género Pseudomonas mostraron actividad antagónica frente a las cepas fúngicas en estudio. Al enfrentar los 17 aislamientos bacterianos a la cepa CH5 de C. pallescens, se demostró que todos presentan efecto antagónico ante el fitopatógeno (Fig. 1A), con valores del 64% hasta un 83% de inhibición del crecimiento del hongo. Con respecto a la cepa C. pallescens AAD-430, se observó que las cepas inhibieron el crecimiento fúngico desde un 59% hasta el 82% (Figura 1B). En el caso del enfrentamiento de los aislamientos a la cepa C. trifolii AAD-533 (Fig. 1C), se demostró que estas tenían la capacidad de inhibir el crecimiento del hongo desde un 49% hasta un 82%.
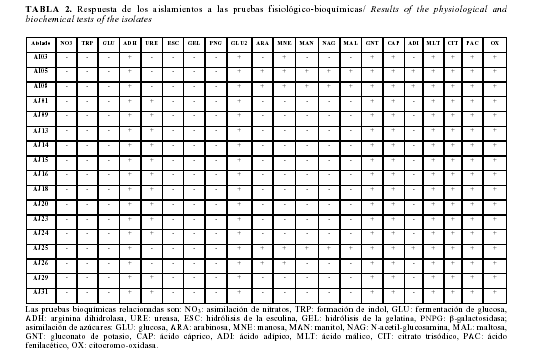
Identificación de los aislamientos seleccionados mediante métodos convencionales: El análisis de los resultados de las pruebas fisiológico bioquímicas mediante el micrométodo API20NE (bioMérieux S.A., Francia) (Tabla 2) demostró que los aislamientos AI05, AI08 y AJ25 pertenecen a las especie P. fluorescens y los aislamientos AI03, AJ9, AJ13, AJ14, AJ15, AJ16, AJ18, AJ20, AJ23, AJ24, AJ25, AJ26, AJ29 y AJ31 a la especie P. putida.
Las pruebas bioquímicas relacionadas son: NO3: asimilación de nitratos, TRP: formación de indol, GLU: fermentación de glucosa, ADH: arginina dihidrolasa, URE: ureasa, ESC: hidrólisis de la esculina, GEL: hidrólisis de la gelatina, PNPG: â-galactosidasa; asimilación de azúcares: GLU: glucosa, ARA: arabinosa, MNE: manosa, MAN: manitol, NAG: N-acetilglucosamina, MAL: maltosa, GNT: gluconato de potasio, CAP: ácido cáprico, ADI: ácido adípico, MLT: ácido málico, CIT: citrato trisódico, PAC: ácido fenilacético, OX: citocromo-oxidasa.
Amplificación del DNA de los aislamientos seleccionados: En los perfiles obtenidos, las cepas previamente descritas como P. fluorescens forman una banda de aproximadamente 800 pb (Fig. 2B), lo que corrobora los resultados obtenidos por los métodos convencionales y confirma que las cepas AI05, AI08 y AJ25 pertenecen a la especie P. fluorescens. Asimismo, en los perfiles relacionados con la especie P. putida se observó una sola banda de aproximadamente 630 pb correspondiente al ADNr 16s de P. putida (Fig. 2A). Estos resultados confirman que las cepas AI03, AJ9, AJ13, AJ14, AJ15, AJ16, AJ18, AJ20, AJ23, AJ24, AJ25, AJ26, AJ29 y AJ31 se encuentran incluidas en la especie P. putida.
DISCUSIÓN
La efectividad de las bacterias promotoras del crecimiento vegetal en el control biológico de patógenos ha sido informada en diferentes estudios (24, 31, 32, 33). Esta investigación ha revelado que las cepas nativas de pseudomonas fluorescentes, identificadas como P. fluorescens y P. putida, tienen la capacidad de ejercer efecto antagónico ante las cepas de C. pallescens (AAD-430 y CH5) y C. trifolii (AAD-533) probadas, especies que son reconocidas como patógenos causantes del manchado del grano en el cultivo del arroz (34), enfermedad que causa pérdidas económicas en Cuba.
Estos resultados se corresponden con los informados por Trujillo et al. (26) y Hernández et al. (24) que demostraron la actividad antagónica de cepas de P. fluorescens ante los patógenos fúngicos Fusarium y Curvularia. Actividad antagónica similar ante patógenos fúngicos ha sido encontrada en estudios realizados por Rives et al. (12) y Acebo (20) que demostraron la efectividad de cepas de Pseudomonas contra P. grisea.
Este comportamiento coincide además con lo informado por Srivastava y Shalini (35) que utilizaron cepas de P. fluorescens para inhibir el crecimiento de Curvularia lunata (Wakk.), patógeno que también está asociado a la enfermedad del manchado del grano.
Se ha demostrado que Pseudomonas constituye un excelente ejemplo de la combinación de múltiples mecanismos para ejercer un efectivo control biológico. Se destaca por producir diversos metabolitos con actividad antimicrobiana hacia otras bacterias y hongos. Entre ellos pudieran citarse la producción de sideróforos (36,37), metabolitos antifúngicos (38) y enzimas líticas (39). Además de la competencia por nutrientes y hábitat (40) y el establecimiento o inhibición de sistemas de moléculas señales en el patógeno (14,41,42).
La producción de sideróforos podría influir en los resultados que se presentan, ya que los bioensayos in vitro se desarrollaron en medio King B que tiene bajo contenido de hierro y favorece la producción de este tipo de metabolitos (43,44). Acebo (20) demostró que estas cepas tienen la capacidad de producir sideróforos bajo condiciones limitantes de hierro en el medio de cultivo Agar Casamino Ácido (al que se adiciona la solución indicadora CAS), observando la formación del halo de color naranja alrededor de la colonia bacteriana, que denota un resultado positivo, a partir de las 7 horas de crecimiento para algunos aislados (AI03, AJ13, AJ29 y AJ31).
En los resultados también podría haber influido la producción de compuestos autoinductores del tipo acil homoserin lactonas (AHL) por las cepas, lo que fue previamente demostrado por Acebo (20). Las AHLs resultan importantes en la regulación de diferentes funciones bacterianas, como la síntesis de antibióticos, producción de factores de virulencia y crecimiento bacteriano expansivo (45).
En este estudio no se observó efecto diferencial de las cepas ante los patógenos fúngicos estudiados (Fig. 2), lo que se corresponde con los resultados informados por Hernández et al. (24), al enfrentar rizobacterias ante cepas de Fusarium verticillioides (Sacc.). Sin embargo, Jaiganesh et al. (46) y Acebo (20), señalaron que al establecerse la interacción entre los antagonistas y los fitopatógenos, existía una variedad de respuestas en función de las cepas, lo que podría estar dado por las características propias de cada microorganismo, que responden con diferentes niveles de resistencia ante el antagonista aplicado.
De modo general, se demuestra que los aislamientos de pseudomonas fluorescentes que presentaban actividad antagónica ante P. grisea, también la presentan ante las cepas de Curvularia estudiadas, lo que indica las potencialidades de estas rizobacterias como agentes de control biológico de patógenos fúngicos en el cultivo del arroz. Este comportamiento, unido a la producción de metabolitos con actividad antifúngica, podría indicar las potencialidades de las cepas de PGPB en estudio en el control biológico, sin embargo, se deben realizar estudios in vivo e in situ para confirmar esta hipótesis.
AGRADECIMIENTOS
Este estudio fue financiado por la Universidad de La Habana como parte del proyecto Alma Mater ``Empleo de Bacterias Promotoras del Crecimiento Vegetal para el desarrollo de agroecosistemas arroceros sostenibles''. Los autores agradecen a Aleida Romero Sacerio por el apoyo en la realización de la parte práctica del trabajo.
REFERENCIAS
1. Arregocés O, Rosero M, González J. Guía de estudio. Morfología de la Planta de Arroz: Cali; 2005; 20p.
2. MINAGRI. Instructivo Técnico de Arroz., Centro Nacional de Sanidad Vegetal; La Habana; 2006.p.50-55.
3. Pinciroli M, Sisterna MN, Bezus R, Vidal AA. Manchado del grano de arroz: efecto de la fertilización nitrogenada. Revista de la Facultad de Agronomía, La Plata. 2004;105(2):88-96.
4. Neninger H, Barrios LM, Hidalgo EI. Micobiota asociada y patogénica presente en semillas de arroz (Oryza sativa L.) nacional e importada en Cuba. Phytopathology 2002; 92 (6) S128.
5. Gutiérrez SA. Micoflora asociada a granos manchados de arroz. En: XI Jornadas Fitosanitarias Argentinas, Río Cuarto, Córdoba, Argentina: 2002; 29p.
6. Bloemberg GV, Lugtenberg BJJ. Molecular basis of plant growth promotion and biocontrol by rhizobacteria. Curr Op Plant Biol. 2001; 4:343-350.
7.Matiru VN, Dakora F. Potential use of rhizobial bacteria as promoters of plant growth for increased yield in landraces of African cereal crops. African J Biotech. 2004;3: 1-7.
8. Hernández-Rodríguez A, Heydrich-Pérez M, Diallo B, El Jaziri, M., Vandeputte OM. Cell-free culture medium of Burkholderia cepacia improves seed germination and seedling growth in maize (Zea mays) and rice (Oryza sativa). Plant Growth Regul. 2010;60:191-197.
9.Velusamy P, Immanuel JE, Gnanamanickam SS, Thomashow L. Biological control of rice bacterial blight by plant-associated bacteria producing 2,4diacetylphloroglucinol. Can J Microbiol. 2006;52:56-65.
10.Joshi F, Archana G, Desai A. Siderophore Cross-Utilization Amongst Rhizospheric Bacteria and the Role of Their Differential Affinities for Fe3+ on Growth Stimulation Under Iron-Limited Conditions. Curr Microbiol. 2006;53:141-147.
11.Rives N, Acebo Y, Hernández-Rodríguez A. Bacterias promotoras del crecimiento vegetal en el cultivo del arroz (Oryza sativa L.). Perspectivas de su uso en Cuba. Cultivos Tropicales 2007;28(2):29-38.
12.Rives N, Acebo Y, Almaguer M, García JC, Hernández A. Actividad antagónica frente a Pyricularia grisea (Sacc.) y fitoestimulación en el cultivo del arroz de cepas autóctonas de Pseudomonas putida (Trev.). Rev Protección Veg 2009;24(2):106-116.
13.Jhaa B, Thakura MC, Gontiaa I, Albrechtb V, Stoffelsb M, Schmidb M, et al. Isolation, partial identification and application of diazotrophic rhizobacteria from traditional Indian rice cultivars. Eur J Soil Biol. 2009;45:62-72.
14.Bolwerk A, Lagopodi AL, Wijfjes AHM, Lamers GEM, Chin-A-Woeng TFC, Lugtenberg, BJJ, et al. Interactions in the Tomato Rhizosphere of Two Pseudomonas Biocontrol Strains with the Phytopathogenic Fungus Fusarium oxysporum f. sp. radicis-lycopersici. MPMI. 2003;16(11):983-993.
15.Persello-Cartieaux F, Nussaume L, Robaglia C. Tales from the underground: molecular plantrhizobacteria interactions. Plant Cell Environm. 2003;26:189-199.
16.Schouten A, van den Berg G, Edel-Hermann V, Steinberg C, Gautheron N, Alabouvette C, et al. Defense Responses of Fusarium oxysporum to 2,4Diacetylphloroglucinol, a Broad-Spectrum Antibiotic Produced by Pseudomonas fluorescens. MPMI 2004;17(11):1201-1211.
17.Allaire M. Diversité fonctionnelle des Pseudomonas producteurs d'antibiotiques dans les rhizosphères de conifères en pépinières et en milieu naturel Thèse de Maître ès Sciences. Québec, Canada: Université Laval. 2005, 90 p.
18.Cazorla FM, Duckett SB, Bergström ET, Noreen S, Odijk R, Lugtenberg BJJ, et al. Biocontrol of Avocado Dematophora Root Rot by Antagonistic Pseudomonas fluorescens PCL1606 Correlates With the Production of 2-Hexyl 5-Propyl Resorcinol. MPMI 2006;19(4):418-428.
19.Yang D, Wang, B, Wang J, Chen Y, Zhou M. Activity and efficacy of Bacillus subtilis strain NJ-18 against rice sheath blight and Sclerotinia stem rot of rape. Biological Control 2009;51(1): 61-65.
20.Acebo Y. Aislamiento y caracterización de cepas de pseudomonas fluorescentes con actividad antagónica ante Pyricularia grisea. Departamento de Microbiología y Virología. La Habana, Facultad de Biología, Universidad de La Habana. Tesis en opción al Título Académico de Maestro en Microbiología; 2009. 88p.
21.Bergey`s Manual of Systematic Bacteriology. 9na Edition. Editors N.R. Krieg and J. Holt. Ed. Williams and Wilkins. Baltimore. London; 1994.
22.Almaguer M, Rojas TI, Batista A, Benitez M, Rives N, Hernández A. Detección de Pyricularia grisea y Curvularia sp. en áreas arroceras en Instituto de Investigaciones del arroz. Revista Cubana del Arroz 2007; 174-177.
23.Mena Portales JDJ. Taxonomía del complejo Bipolaris, Curvularia, Drechslera y Exserohilum en Cuba. Instituto de Ecología y Sistemática, La Habana; 2004; 108p.
24.Hernández A, Heydrich M, Acebo Y, Velázquez MG, Hernández AN. Antagonistic activity of Cuban native rhizobacteria against Fusarium verticillioides (Sacc.) Nirenb. in maize (Zea mays L.). Appl Soil Ecol. 2008;36:184-186.
25.Harrigan WF, Mac Cance M. Métodos de Laboratorio en Microbiología. E. Academia. León, España; 1968; 426p.
26.Trujillo I, Díaz A, Hernández-Rodríguez A, Heydrich M. Antagonismo de cepas de Pseudomonas fluorescens y Burkholderia cepacia contra hongos fitopatógenos del arroz y el maíz. Rev Protección Veg. 2007;2:7-14.
27.Bramwel PA, Barallon RV, Rogers HJ, Bailey MJ. Extraction and PCR amplification of DNA from the rhizoplane. Molecular Microb Ecol Man; 1995;1.4.2:1-20.
28.Scarpellini M, Franzetti L, Galli A. Development of PCR assay to identify Pseudomonas fluorescens and its biotype. FEMS Microbiology Letters. 2004;236(2):257-260.
29.Aranda-Olmedo I, Tobes R, Manzanera M, Ramos JL, Marqués S. Species-specific repetitive extragenic palindromic (REP) sequences in
Pseudomonas putida. Nucleic Ac Res. 2002;30(8):1826-1833.
30.Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning. Ed. Cold Spring Harbor Laboratory Press, 2nd Edition; 1989; 1090 pp.
31.Bashan Y, de-Bashan LE. Plant Growth-Promoting Bacteria. Encyclopedia of soils in the environment. D. Hillel. Oxford, U.K., Elsevier; 2005;1:103-115.
32.Berg G, Krechel A, Ditz M, Sikora RA, Ulrich A, Hallmann J. Endophytic and ectophytic potato-associated bacterial communities differ in structure and antagonistic function against plant pathogenic fungi. FEMS (Fed Eur Microbiol Soc) Microbiol Ecol 2005;51:215-229.
33.Compant S, Duffy B, Nowak J, Clément C, Ait Barka E. Use of Plant Growth-Promoting Bacteria for Biocontrol of Plant Diseases: Principles, Mechanisms of Action, and Future Prospects. Appl Environ.Microbiol. 2005;71:4951-4959.
34.Estrada G, Sandoval I. Patogenicidad de especies de Curvularia en arroz. Fitosanidad. 2004;8:23-26.
35.Srivastava R Shalini S. Antifungal Activity of Pseudomonas fluorescens against different plant pathogenic fungi. EJEAFChe 2008;7:2881-2889.
36.Cornelis P, Matthijs S. Diversity of siderophore mediated iron uptake systems in fluorescent pseudomonas: not only pyoverdines. Environ. Microbiol. 2002;4:787-798.
37.Ahmad F, Ahmad I, Khan MS. Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol Res. 2008;163(2):173-181.
38.Rezzonico F, Zala M, Keel C, Duffy B, Moënne-Loccoz Y, Défago G. Is the ability of biocontrol fluorescent pseudomonas to produce the antifungal metabolite 2,4-diacetylphloroglucinol really synonymous with higher plant protection? New Phytologist. 2007;173:861-872.
39.Han J, Sun L, Dong X, Cai Z, Sun X, Yang Y, et al. Characterization of a novel plant growth-promoting bacteria strain Delftia tsuruhatensis HR4 both as a diazotroph and a potential biocontrol agent against various plant pathogens. System Appl Microbiol. 2005;28:66-76.
40.Martínez-Granero F, Rivilla R, Martín M. Rhizosphere Selection of Highly Motile Phenotypic Variants of Pseudomonas fluorescens with Enhanced Competitive Colonization Ability. Appl Environ Microbiol. 2006;72(5):3429-3434.
41.Bertani I, Venturi V. Regulation of the N-Acyl Homoserine Lactone-Dependent Quorum-Sensing System in Rhizosphere Pseudomonas putida WCS358 and Cross-Talk with the Stationary-Phase RpoS Sigma Factor and the Global Regulator GacA. Appl Environ Microbiol. 2004;70(9):5493-5502.
42.Preston GM. Plant perceptions of plant growth-promoting Pseudomonas. Phil.Trans. R. Soc. Lond; 2004; B 359:907-918.
43.Loper JE, Henkels MD. Utilization of Heterologous Siderophores Enhances Levels of Iron Available to Pseudomonas putida in the Rhizosphere. Appl Environ Microbiol. 1999;65(12):5357-5363.
44.Haas D, Défago G. Biological control of soil-borne pathogens by fluorescent pseudomonas. Nature Reviews Microbiol. 2005;3:307-319.
45.Keller L, Surette MG. Communication in bacteria: an ecological and evolutionary perspective. Nature Reviews Microbiol. 2006;4:249-258.
46.Jaiganesh V, Eswaran A, Balabaskar P, Kannan C. Antagonistic activity of Serratia marcescens against Pyricularia oryzae. Not Bot Hort Agrobot Cluj. 2007;35(2):48-54.


{kind=link}
{kind=link}
{kind=link}
{kind=link}