










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión impresa ISSN 1010-2752versión On-line ISSN 2224-4697
Rev. Protección Veg. v.25 n.3 La Habana sep.-dic. 2010
VARIABILIDAD GENÉTICA ENTRE GENOTIPOS PROMISORIOSDE TOMATE (Solanum lycopersicum) OBTENIDOS EN PROGRAMASDE MEJORAMIENTO FRENTE AL TYLCV
GENETIC VARIABILITY AMONG TOMATO PROMISSORY GENOTYPES OBTAINEDIN IMPROVEMENT PROGRAMS IN FRONT OF TYLCV
Yailen Arias*, Belkis Peteira*, Ivonne González*, Yamila Martínez*, Ileana Miranda**
*Grupo de Fitopatología, **Grupo Plagas Agrícolas, División Protección de Plantas, Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, La Habana, Cuba. Correo electrónico: yailenav@censa.edu.cu
RESUMEN
Una de las alternativas más usadas en la actualidad, para el manejo de fitopatógenos, es el empleo de variedades resistentes. La disponibilidad de una amplia variabilidad genética resulta de vital importancia para el éxito de un programa de mejoramiento. Los marcadores moleculares constituyen poderosas herramientas para la caracterización de la diversidad genética. El objetivo del trabajo fue conocer la variabilidad genética entre seis genotipos promisorios de tomate obtenidos en el programa de mejoramiento genético para la resistencia al TYLCV mediante el empleo de los RAPDs. Los resultados obtenidos mostraron, de forma general, una escasa variabilidad entre los genotipos estudiados.
Palabras clave: TYLCV; variabilidad genética; RAPD; Solanum lycopersicum
ABSTRACT
The introduction of varieties resistant to phytopatogens is one of the most used alternatives at present. The availability of a broad genetic variability is very important for the success of an improvement program. The molecular markers are powerful tools to study the genetic diversity characterization. The aim of this work was to know the genetic variability among six promissory tomato genotypes obtained in breeding programs for TYLCV resistance by means of RAPDs. Scarce genetic variability among studied genotypes was observed in the results.
Key words: TYLCV; genetic variability; RAPD; Solanum lycopersicum
(Recibido 14-3-2010; Aceptado 8-7-2010)
El TYLCV (Tomato Yellow Leaf Curl Virus) constituye en la actualidad uno de los patógenos virales más devastadores del cultivo del tomate (Solanum lycopersycum L.) (1,2). Por la importancia que tiene esta hortaliza en los hábitos alimenticios de gran parte de la población mundial (3), se han establecido pro-gramas de manejo que incluyen la aplicación de métodos tradicionales, el control físico, químico y biológico sobre el vector (4), así como el empleo de variedades resistentes, siendo este último método una alternativa más amigable para la salud del hombre y para el medio ambiente, en general (5,6).
Con el propósito de obtener nuevas variedades que sean altamente resistentes al TYLCV, en nuestro país se han establecido importantes programas de mejoramiento genético (7), los cuales dependen, en gran medida, de la disponibilidad de una amplia variabilidad genética (8,9).
En la actualidad, los marcadores moleculares constituyen una herramienta útil por su aplicación tanto en la identificación de accesiones como en la caracterización de la diversidad genética para el mejoramiento (10,11). Los marcadores RAPD (Ramdon Amplified Polimorphism DNA), particularmente, constituyen una técnica poderosa para la identificación de variaciones en la secuencia de ADN entre individuos (11,12).
El presente trabajo se realizó con el objetivo de estudiar la variabilidad genética existente entre seis genotipos promisorios de tomate, obtenidos en pro-gramas de mejoramiento genético frente al TYLCV, mediante el empleo de los RAPD.
Los genotipos utilizados en el análisis (Tabla 1) proceden del programa de mejoramiento desarrollado en el Instituto de Investigaciones Hortícolas «Liliana Dimitrova» (IIHLD), en la provincia de La Habana, Cuba. La extracción del ADN se realizó por el método de Dellaporta et al. (13) a partir de una mezcla de hojas jóvenes frescas, recolectadas de cinco individuos para cada genotipo, en plantas completamente sanas y con cuatro semanas de germinadas, las cuales habían sido mantenidas en condiciones semicontroladas de casa de cristal. Los precipitados finales obtenidos se disolvieron en solución amortiguadora TE (Tris 50 mM, EDTA 0,5 mM), se conservaron a -20°C y la concentración de ADN total se determinó mediante la lectura de la absorbancia a 260 nm (14).
ciclo de 10 minutos a 72°C. Los productos de la PCR se visualizaron por electroforesis en gel de agarosa al 1,5% en solución amortiguadora TBE (45mM Tris-Borato, 1mM EDTA), y se tiñeron con bromuro de etidio (14). El peso molecular de los fragmentos amplificados se estimó usando el marcador 1Kb DNA Ladder (Gibco BRL).
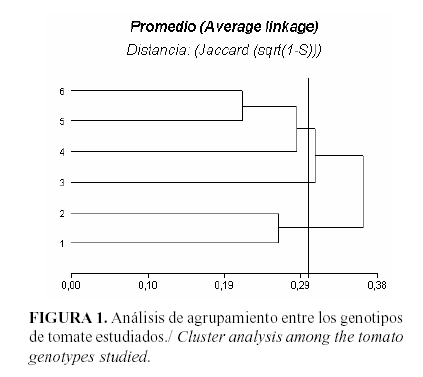
Los productos de la PCR se informaron como presentes [1] o como ausentes [0] en cada genotipo, creando así una matriz de valores binarios. Se analizaron las bandas más intensas y con los datos obtenidos se calculó el índice de similitud entre cada par de genotipos, según Jaccard (15), los que fueron empleados para la construcción de un dendrograma. Estos análisis se realizaron mediante el programa InfoStat (16).
Los 12 cebadores empleados fueron capaces de amplificar un total de 92 bandas, con un promedio de 7 bandas por cebador, destacándose el OPA2 con 11 bandas amplificadas, mientras que el OPA11 solo amplificó 5 bandas. También es importante señalar que de las 92 bandas amplificadas, solo 20 permitieron encontrar diferencias, en lo cual influyó principalmente el cebador OPA2 con 6 bandas polimórficas de las 11 amplificadas.
El dendrograma resultante de las distancias genéticas mostró la formación de tres grupos para un 70% de similitud aproximadamente. Uno de ellos quedó formado por los parentales, los cuales son genotipos que provienen del mismo cruzamiento, por lo que tienen progenitores en común (Fig.1 ). Sin embargo, se conoce que difieren en cuanto a la resistencia frente al TYLCV (17). Debido a esto es posible que los cebadores empleados no hayan sido capaces de amplificar las regiones que codifican para esta diferencia. En otra rama se agruparon la F1 y los dos retrocruzamientos, resultados que no son sorprendentes por el nivel de parentesco que existe entre los progenitores. El tercer grupo quedó formado solamente por la F2, que aunque también forma parte de la descendencia, existen mayores probabilidades de recombinación y segregación, lo que pudo influir en el hecho de que formara un grupo por separado.
Los marcadores RAPD pueden conducir, en ocasiones, a una subestimación de la cantidad de polimorfismo existente entre individuos, debido entre otros elementos, a que ellos son dominantes, por lo que no tienen posibilidad de diferenciar heterosis de homocigosis (18). Sin embargo a pesar de este inconveniente siguen siendo ampliamente usados como una primera aproximación en los estudios de variabilidad genética en las plantas (11).
Para el desarrollo de la técnica RAPD se utilizaron los siguientes cebadores arbitrarios: OPA2, OPA3, OPA4, OPA5, OPA8, OPA9, OPA10, OPA11, OPA12, OPA13, OPA20, OPF1, provenientes de la firma comercial Operon Technologies INC, Alameda, California. La reacción de amplificación se realizó en un volumen total de 25µL que contenía: 10mM Tris-HCl a pH 8.3; 50mM KCl, 2mM MgCl2, 0,001% de gelatina, 100µM de cada dNTPs, 0,2µM de cebador, 50ng de ADN genómica y 1U de Taq ADN polimerasa (Amplicen). La amplificación se produjo en un termociclador marca Master Cycler, programado para 45 ciclos de: 1 minuto a 94°C, 1 minuto a 36°C y 2 minutos a 72°C, y un
De forma general, se encontró escasa variabilidad entre los genotipos estudiados, lo que no resulta contradictorio ya que todos los genotipos están estrechamente emparentados. Resultados similares de baja variabilidad en el cultivo del tomate, mediante el empleo de los RAPD, han sido informados con anterioridad (19), atribuyendo la escasez de diferencias, principalmente, al proceso de desarrollo de variedades comerciales (9,19). Estos resultados han sido corroborados además, con el uso de marcadores isoenzimáticos, a través de los cuales se ha encontrado una gran homogeneidad dentro del género Lycopersicon (20) y también utilizando técnicas moleculares como los AFLP (Amplified Fragment Length Polymorphism) (21) y los SSR (Simple Sequence Repeat) (9). No obstante, se recomienda ampliar el número de cebadores a utilizar en este tipo de estudio, seleccionando aquellos que rinden más bandas producto de la amplificación.
REFERENCIAS
1. Péres de Castro A, Diez MJ, Nuez F. Inheritance of tomato yellow leaf curl virus resistance derived from Solanum pimpinellifolium UPV 16991. Plant Dis. 2007;91:879-885.
2. Raj SK, Singh R, Pandey SK, Singh BP. Agrobacterium-mediated tomato transformation and generation of transgenic lines expressing Tomato leaf curl virus coat protein gene for resistance against TYLCV infection. Curr Sci. 2005;88(10):1674-1679.
3. Florido M. Evaluación de la tolerancia al calor en muestras de germoplasma de tomate (Solanum L. sección Lycopersicon subsección Lycopersicon) conservado ex situ en Cuba. Tesis en opción al grado de Doctor en Ciencias Agrícolas. La Habana, Cuba; 2007.
4. Martínez Y, Quiñónez M, Fonseca D, Miranda I. Prospección nacional de begomovirus presentes en el cultivo del tomate en Cuba. Rev Protección Veg. 2003;18(3):168-175.
5. Martínez AK, Morales FJ, Vallejo FA. Carac-terización molecular de un begomovirus del tomate en el Valle del Cauca, Colombia, y búsqueda de fuentes de resistencia para el mejoramiento de la variedad Unapal Maravilla. Acta Agron. 2008;57(3):167-173.
6. González-Cabezuelo JM, Tomás DM, Lorente I, Abad J, Fernández-Muñóz R, Lozano R. Marcadores moleculares ligados a la resistencia a TYLCD en tomate. Actas de Horticultura nº 48. Sociedad Española de Ciencias Hortícolas. XI Congreso SECH. Albacete 2007.
7. Dueñas F, Martínez Y, Álvarez M, Moya C, Peteira B, Arias Y. Identificación de los genes ty-2 y ty-3 de resistencia a begomovirus y su grado de homocigosis en nuevas accesiones de tomate. Cultivos Tropicales. 2009;30(1):61-64.
8. Paredes W. Mejoramiento genético en plantas [monografía en Internet]. Universidad Nacional de San Agustín Arequipa PERÚ 2007[citado 27 Enero 2010]. Disponible en: http:// www.monografias.com/trabajos53/mejoramiento plantas/mejoramiento-plantas.shtml
9. Chen J, Wang H, Shen H, Chai M, Li J, Oi M, et al. Genetic variation in tomato population from four breeding programs revealed by single nucleotide polymorphism and simple sequence repeat markers. Scientia Horticulturae. 2009;122:6-16.
10.Azofeifa-Delgado A. Uso de marcadores moleculares en plantas; aplicaciones en frutales del trópico. Agronomía Mesoamericana. 2006;17(2):221-242.
11.Venkatachalam L, Sreedhar RV, Bhagyalakshmi N. The use of genetic markers for detecting DNA polymorphism, genotype identification and phylogenetic relationships among banana cultivars. Mol Phylogen and Evol. 2008;47:974-985.
12.González O, Fernández A, Fraga Y, Pino B, Hernández MM, Silava J J. Evaluación de la estabilidad genética mediante marcadores RAPD en plantas de Ipomoea batatas. Cultivos Tropicales. 2007; 28(2):39-43.
13.Dellaporta SL, Word J, Hichs JR. A plant molecular DNA minipreparation, version II. Plant Mol Biol Rep. 1983;1:19-21.
14.Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: a laboratory manual. 2nd ed. Cold Spring Harbor: N Y, USA; 1989.
15.Jaccard P. Extude comparative de la distribution florale dans one partion des Alpes et des Jura. Bull Soc Vaudoise Sci Nat. 1901;37:547-579.
16.InfoStat version 2009. Grupo InfoStat FCA, Universidad Nacional de Córdoba, Argentina.
17.Piñón, M. Obtención de líneas de tomate (Solanum lycopersicon L.) resistentes al Virus del Encrespamiento Amarillo de la Hoja del Tomate (TYLCV) en Cuba. Tesis en opción al grado de Doctor en Ciencias Agrícolas. INCA; 2009.
18.Méndez A, Román-Bravo R, Isea W, Villasmil Y, Jordana J. Los microsatélites (STR's), marcadores moleculares de ADN por excelencia para programas de conservación: una revisión. Archivos Latinoamericanos de Producción Animal. 2005;13(1):30-42.
19.Florido, M. Caracterización de variedades y especies silvestres de tomate atendiendo a características morfo-bioquímicas y tolerancia al calor. Tesis en opción al grado de Maestro en Biología Vegetal. La Habana, Cuba: Universidad de La Habana; 1999.
20.Florido, M, Álvarez M, Lara R M, Plana D, Valera M, Shagarodsky T, Moya C. Caracterización morfoagronómica y bioquímica de 20 accesiones de tomate (Lycopersicon spp). Cultivos Tropicales. 2002;23(4):61-69.
21.Florido M, Arencibia A, Plana D, Álvarez M, López J, Lara RM. Análisis de la diversidad genética en tomate (Solanum L. sección Lycopersicon subsección Lycopersicon) utilizando AFLP. Cultivos Tropicales. 2007;28(3):83-87.


{kind=link}
{kind=link}