










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión On-line ISSN 2224-4697
Rev. Protección Veg. v.26 n.1 La Habana ene.-abr. 2011
ARTÍCULO ORIGINAL
CARACTERIZACIÓN BIOQUÍMICA DE SEIS AISLAMIENTOS DE Beauveria bassiana (BALSAMO) VUILLEMIN
BIOCHEMICAL CHARACTERIZATION OF SIX ISOLATES OF Beauveria bassiana (BALSAMO) VUILLEMIN
Belkis Peteira*, Ivonne González*, Yailen Arias*, A. Fernández Turro**, Ileana Miranda*, B. Martínez*
*Centro Nacional de Sanidad Agropecuaria, Autopista Nacional y Carretera de Tapaste, Apartado 10, San José de las Lajas, Mayabeque, Cuba. Correo electrónico: bpeteira@censa.edu.cu; **Facultad Agroforestal, Centro Universitario de Guantánamo, Carretera a Santiago de Cuba, Km 1½, Guantánamo, Cuba
RESUMEN
Beauveria bassiana (Balsamo) Vuillemin es un agente de control biológico para artrópodos plaga. En el proceso de desarrollo de un agente de control biológico como producto, es necesario el estudio de su modo de acción, los metabolitos que en el participan y su expresión en las cepas del hongo, a fin de garantizar conocimiento útil para la selección, optimización de la producción, formulación e identidad de la cepa. El objetivo del presente trabajo fue la caracterización bioquímica de seis cepas de B. bassiana a través de un estudio de dinámica de inducción en diferentes medios de cultivo. Para ello se sembraron discos de micelio de seis cepas de Beauveria bassiana en medio basal como control, medio basal suplementado con gelatina y medio basal suplementado con quitina. Se realizaron determinaciones de proteínas totales, actividad quitinasa y actividad glucanasa, a los 3, 5 y 7 días postinoculación. De manera general, las cepas 33 y 9 mostraron los mayores niveles de producción de los parámetros evaluados. El medio suplementado con quitina resultó más favorable para la producción de proteínas totales y de quitinasa, mientras que el medio suplementado con gelatina fue el mejor para la producción de glucanasa.
Palabras clave: Beauveria bassiana; actividad enzimática; proteínas totales; quitinasas; glucanasas
ABSTRACT
Beauveria bassiana (Balsamo) Vuillemin is a promisoring biological control agent for invertebrate pests. In the development of a product based on a biological control agent, the study of the mode of action and the metabolites involved and their expression by the promisoring strains is needed for selection, mass rearing, formulation and strain identity. The aim of this work was the biochemical characterization of six B. bassiana isolates by a time course experiment in different culture media. Mycelia disk of six B bassiana isolates were grown in a basal medium as the control, basal medium supplemented with chitin and basal medium supplemented with gelatine. Total proteins, chitinase and glucanase activities were assayed at 3, 5 and 7 days post inoculation. In general, isolates 33 and 9 showed the highest levels for all the parameters studied. The medium supplemented with chitin was the best for total protein and chitinase activity expression, while the media supplemented with gelatine was the best for glucanase production.
Key words: Beauveria bassiana; enzimatic activity; total proteins; chitinases; glucanases
INTRODUCCIÓN
La lucha biológica se plantea en la actualidad como una alternativa amigable con el medio ambiente, más favorable que el control químico. El hongo entomopatógeno Beauveria bassiana está siendo estudiado de manera intensiva como agente de control biológico, para insectos y otras plagas artrópodos, por su amplia gama de organismos diana (1, 2). Además de ser un parásito necrótrofo, B. bassiana puede ser saprófito facultativo y puede existir en la plantas como endófito (3, 4, 5).
El potencial de este microorganismo en el control biológico de insectos ha permitido su inclusión en diversos programas de manejo integrado de plagas que atacan cultivos de importancia económica en el mundo. En Cuba, actualmente se utiliza en el control biológico de insectos plaga como la broca del café (Hypothenemus hampei Ferrari) (Coleoptera: Scolytidae) (6).
La acción del hongo sobre el insecto es facilitada principalmente por una gran variedad de metabolitos secundarios producidos a lo largo del proceso de infección. El proceso patogénico se inicia en la cutícula del insecto, con la germinación de los conidios y la producción de hifas invasoras, las cuales penetran los tejidos a través de los intersticios y partes blandas del insecto, dando inicio de esta forma a la actividad enzimática degradativa de la cutícula. Las hifas se ramifican, colonizan y llegan hasta la cavidad hemocélica del insecto, donde se produce una masa micelial por el crecimiento del hongo. En muchos casos, el insecto puede ser colonizado totalmente por el hongo. Además, se liberan toxinas, las cuales están implicadas en el bloqueo del desarrollo fisiológico y pueden causar la muerte del insecto (7). La presencia de enzimas hidrolíticas suele facilitar cada etapa de infección del hongo y adicionalmente puede ser importante en la invasión del hemocele del insecto (8).
En el proceso de desarrollo de un agente de control biológico como producto, es necesario él estudio de su modo de acción, los metabolitos que en el participan y su expresión en las cepas promisorias, a fin de garantizar aspectos del conocimiento que tributan a las fases de selección, optimización de la producción, formulación e identidad de la cepa, entre otras. El objetivo del presente trabajo fue la caracterización bioquímica de seis aislamientos de B. bassiana a través de un estudio de dinámica de inducción en diferentes medios de cultivo.
MATERIALES Y MÉTODOS
En el presente trabajo se emplearon las cepas de Beauveria bassiana 3, 5, 9, 14, 23 y 33, procedentes del cepario del Laboratorio de Micología Vegetal de la Facultad Agroforestal, Centro Universitario de Guantánamo, seleccionados por poseer alta capacidad como agentes de control biológico de la broca del café Hypothenemus hampei (Ferr.). Todas las cepas se conservaron en medio Agar-Malta (Biocen) a 4ºC y para su uso se subcultivaron en medio PDA (Papa Dextrosa Agar, Biocen) en placas Petri de 9 cm, las cuales se incubaron a 28ºC durante 3 días, momento en que se tomó el micelio para el desarrollo del experimento de dinámica.
Para la inducción de proteínas en medio líquido se ensayaron tres variantes diferentes: a) medio basal líquido que contenía extracto de levadura 1g.L-1 y peptona 4g.L-1 (empleado como control), b) medio basal líquido suplementado con quitina (5g.L-1 ) y c) medio basal líquido suplementado con gelatina al 0,2% (p/v). Los dos primeros medios se esterilizaron a 120ºC durante 20 min, mientras que el medio que contenía gelatina se esterilizó a 115ºC durante 15 min.
Para todas las variantes, se dispensaron 20mL del medio en frascos estériles de 100mL, los cuales se inocularon con cuatro discos de micelio de 5mm de diámetro de la periferia de la colonia pura de cada cepa. Posteriormente, se incubaron estáticamente a 28ºC, en la oscuridad. Para cada tratamiento se realizaron tres repeticiones. Los frascos se retiraron de la incubadora al tercer, quinto y séptimo días después de la inoculación, los sobrenadantes se filtraron a través de papel de filtro Whatman 3 y se conservaron a -20ºC, hasta su utilización en los análisis enzimáticos.
Para la determinación de los contenidos de proteínas totales se siguió el protocolo descrito por Bradford (9), realizando las lecturas de la absorbancia a 595nm del complejo proteína-Azul de Coomasie G-250, en un espectrofotómetro (Lasso Spec III, Lasso Biotech LTDA) y empleando para la curva patrón una solución de 1mg.mL-1 de albúmina de suero bovina (BSA).
La actividad quitinasa se determinó con 0,2 mL de quitina coloidal 10 mg.mL-1 preparada acorde a Boller et al. (10) como sustrato y se mezcló con 0,5 mL de muestra sobrenadante. Para el resto de la determinación se siguió el protocolo descrito por González et al. (11). La curva patrón se determinó utilizando N-acetilglucosamina, a partir de una solución madre de 1mg.mL-1 a la cual se le realizó el mismo procedimiento que a las muestras. La lectura se realizó a 585 nm. El cálculo de la actividad enzimática se realizó según la expresión:
Donde DO: densidad óptica; cot: cotangente del ángulo de la curva patrón; Vens: volumen de ensayo; Dil: dilución; Tincub: tiempo de incubación y Venz: volumen de enzima. Las unidades de actividad fueron: mmoles de producto formado.min-1.mL-1 de enzima.
Para la determinación de la actividad enzimática de las 1,3 b-glucanasas se utilizó una variante en microplacas de 96 pozos desarrollada por Zheng y Wozniak (12).
La actividad específica para ambas enzimas se determinó según la expresión:
En todas las determinaciones se realizaron tres réplicas. Para simplificar el análisis estadístico de los datos obtenidos se empleó el Análisis Multivariado de Componentes Principales por el paquete estadístico InfoStat (13), el cual permitió además determinar el (los) medio(s) y el (los) día (días) que favorecieron la inducción de la actividad proteínas totales, quitinasas y glucanasas (por separado para cada determinación, así como definir el (los) aislamiento(s) que presenta los mayores niveles de expresión de estas enzimas.
RESULTADOS Y DISCUSIÓN
Beauveria bassiana (Balsamo) Vuillemin (Hypocreales; Clavicipitaceae), con una amplia gama de hospedantes de unas 700 especies de insectos, es un bioinsecticida registrado empleado en el manejo de plagas insectiles de diferentes cultivos (14, 15).
A pesar de la prevalencia de B. bassiana en un gran número de artrópodos, es conocido que la mayoría de los aislamientos de este hongo tienen una gama de hospedantes restringida y que existen algunos ejemplos de aislamientos de B. bassiana a partir de diferentes insectos o de suelo, que son altamente virulentos en otras plagas diana. Es por esto que es necesario el estudio de esta característica de diferentes aislamientos para seleccionar los más virulentos (16).
La morfología de los conidios no ha mostrado diferencias considerables en las comparaciones entre distintos aislamientos (17). Tampoco existe correlación de la virulencia con el tamaño de los conidios, la producción de pigmento rojo ni el crecimiento micelial (18). Las características culturales han sido empleadas para separar algunas poblaciones de B. bassiana, pero los aislamientos individuales no siempre pueden ser distinguidos por estos métodos. Una adecuada y segura separación de cepas de B. bassiana es extremadamente importante para su explotación en el control de plagas (17).
Los hongos entomopatógenos tienen diferentes atributos que determinan su virulencia frente a sus hospedantes, incluyendo la producción de enzimas hidrolíticas (19). Durante el proceso de infección Beauveria spp. produce enzimas proteolíticas y toxinas, mientras los insectos hospedantes responden con reacciones de defensa humoral y celular (16). El mecanismo de infección de B. bassiana se inicia cuando el conidio se adhiere a la cutícula del insecto hospedante susceptible. El conidio germina en la superficie del cuerpo del insecto y de modo mecánico penetra el integumento a través del tubo germinativo. Durante este proceso intervienen enzimas extracelulares relacionadas con la patogénesis tales como: proteasas, lipasas, ureasas y quitinasas entre otras, que hidrolizan los componentes cuticulares. La presencia de enzimas hidrolíticas suele facilitar cada etapa de infección del hongo y adicionalmente puede ser importante en la invasión del hemocele del insecto (20).
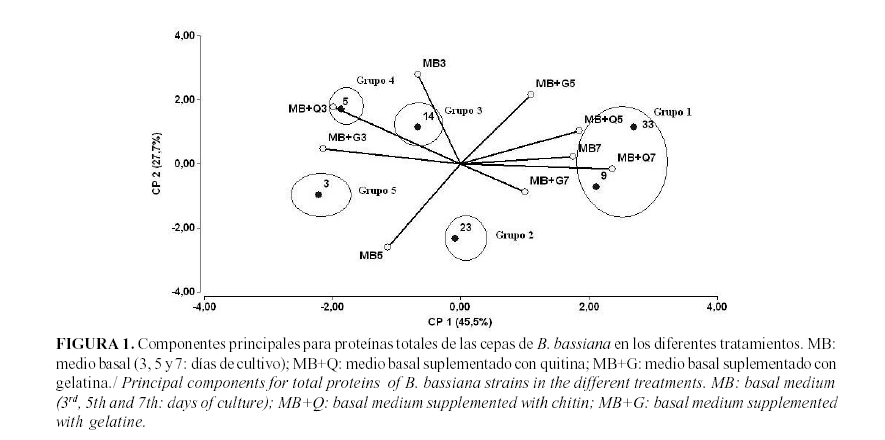
Teniendo en cuenta su modo de acción, estas enzimas hidrolíticas han atraído la atención como objeto de estudio. Aunque las proteínas totales no han sido identificadas como un parámetro importante, si deben ser tenidas en cuenta para el cálculo de la actividad específica de las enzimas. El análisis de las seis cepas estudiadas mostró que las cepas 9 y 33 fueron las mayores productoras de proteínas (Fig. 1). Los más altos niveles de síntesis de proteínas totales en estas cepas fueron expresados en los medios suplementados con quitina, detectándose al quinto día para la cepa 33 y al séptimo para la 9. La cepa que presentó los niveles de proteínas totales más bajos fue la 3.
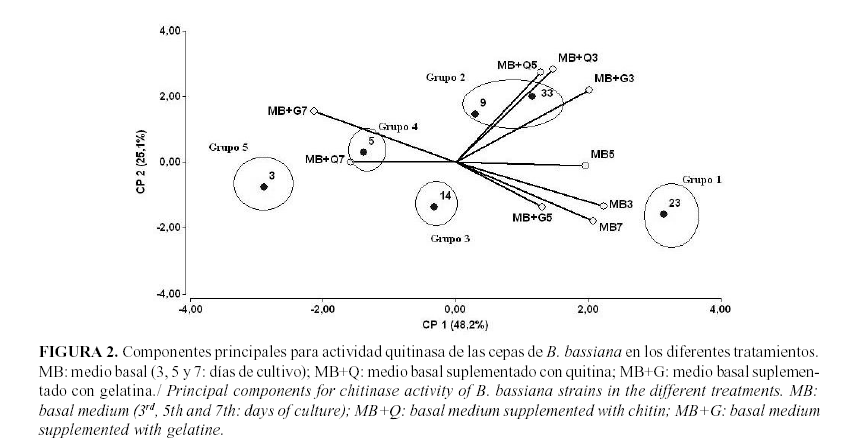
El análisis de las enzimas con actividad quitinasa muestra resultados similares (Fig. 2). Aunque en este caso la cepa 23 es la más productora de esta enzima, la cual se expresó en mayores niveles en el medio basal a los tres días; las cepas 9 y 33 aparecen localizadas en el segundo grupo, estimuladas nuevamente por la adición de quitina al medio y alcanzando los mayores niveles al quinto y tercer día, respectivamente. Es de notar que tanto en proteínas totales como en quitinasas, la cepa 33 alcanza los mayores niveles de expresión más temprano que la otra cepa con mejor producción del parámetro evaluado.
Existen estudios sobre el modo de acción de los agentes de control biológico en los cuales el medio de cultivo es suplementado con macerados provenientes del organismo diana. El integumento de los insectos (constituido por quitina) ha sido empleado como fuente de nutrición para hongos especializados. González et al. (21), estudiaron la relación entre la patogenicidad de cuatro aislamientos de B. bassiana (Bb9007, Bb9009, Bb9015 y Bb9205) aislados de insectos de los órdenes Coleoptera, Hemiptera y Lepidoptera, cultivados en medio SDA (Sabouraud Dextrosa Agar) suplementado con integumento de broca al 0,5%, en solución amortiguadora de fosfato pH 7,0. Los resultados de estos investigadores mostraron incrementos altamente significativos, superiores al 80%, en los cuatro aislamientos evaluados y concluyeron que la mayoría de los aislamientos de B. bassiana puede causar alta mortalidad a la broca, si permanentemente se proporciona al hongo el sustrato requerido para desarrollarse e inducir las enzimas requeridas para su patogenicidad (21).
En las formulaciones de diferentes hongos promisorios como agentes de control biológico es común emplear aditivos que mejoren la eficacia de los mismos. Uno de estos aditivos es la quitina. En el caso de B. bassiana, Gerding-González et al. (22), desarrollaron un estudio para evaluar la concentración óptima de quitina en una formulación con pellet de alginato de sodio, con el objetivo de aumentar la producción de conidios de la cepa Qu-B306 de B. bassiana. Con el empleo de quitina al 2% se lograron incrementos de hasta tres veces superiores al número de conidios inicial, mientras niveles superiores de quitina provocaron disminución del número de conidios por pellet (22). Es posible que en nuestros resultados la presencia de quitina en el medio actuara también como estimulante para la producción y secreción de este tipo de enzimas hidrolíticas ante la presencia del sustrato sobre el cual la enzima desarrolla su actividad biológica. La inclusión de quitina, por lo tanto, es un aspecto a estudiar y a tener en cuenta en estudios futuros de conservación de las cepas promisorias o de formulaciones de estas cepas autóctonas.
Algunas enzimas han sido caracterizadas para B. bassiana. Por ejemplo en medio líquido YPDA (Extracto de Levadura Papa Dextrosa Agar)/ YDA (Extracto de Levadura Dextrosa Agar), diferentes aislamientos del hongo fueron capaces de producir y liberar al medio de cultivo una [alfa]-amilasa digestiva. No se observaron variaciones en la intensidad de la actividad en esta banda para los diferentes aislamientos estudiados. Los autores de este estudio plantean que este es un método simple y sensible que podrá usarse satisfactoriamente no sólo para la evaluación de diferentes hongos entomopatogénicos, sino también, para hacer una determinación rápida de posibles variaciones genéticas debidas a la presencia o ausencia de una enzima especifica.(23). Nuestros resultados apoyan estas aseveraciones ya que los medios de cultivo empleados son sencillos, baratos, se emplean volúmenes muy pequeños y el método permite en pocos días después de la inducción realizar las determinaciones y contar con un parámetro mas a tener en cuenta para la evaluación del desempeño de hongo.
No obstante, las proteasas han sido las enzimas más estudiadas en B. bassiana. Se ha informado de la existencia de una proteasa (nombrada CDEP-1) la cual fue expresada en la levadura Pichia pastoris y usada como aditivo a tres gradientes de aspersión de la cepa Bb0062 de B. bassiana sobre adultos de Myzus persicae. Este grupo de investigación, propone que la proteasa en las aspersiones del hongo pudiera actuar sobre la cutícula del hospedante y los conidios del hongo o afectar la interacción solamente en el curso de la infección, pero no como causa directa de la muerte del insecto, lo cual es una consecuencia de la micosis atribuida a la proliferación de las células del hongo en la hemolinfa del hospedante, después de la penetración de la cutícula. Sus resultados confirmaron que la proteasa CDEP-1 incrementa la virulencia del hongo debido a la aceleración de la germinación de los conidios y la penetración de la cutícula, lo cual constituye un nuevo punto de vista para la utilización de proteasas en el control microbiano (20).
Estudios basados en la sobreexpresión de la quitinasa Bbchit1 resultaron en un incremento de la virulencia de B. bassiana. Los transformantes de B. bassiana que segregaban una fusión de la proteasa y la quitinasa (CDEP1:Bbchit1), penetraron la cutícula del insecto mucho más rápido, con una reducción de la dosis letal media en un 60,5%. Este trabajo representa un avance significativo hacia el desarrollo de patógenos de insectos hipervirulentos para un control de plagas efectivo (24) y pone de manifiesto la importancia de las quitinasas.
En la actualidad existen diversos datos que confirman la existencia de otras enzimas insecticidas de B. bassiana. Diferentes estudios se han desarrollado para investigar las enzimas quitinolíticas en B. bassiana entre los que se incluyen las quitinasas de 45 y 110 kDa (Havukkala et al., 1993), una proteína similar a la quitinasa con 28 kDa (Fuguet et al., 2004), una quitinasa con 33 kDa (Fang et al., 2005) y una proteína quitinolítica con 55 kDa (Murad et al., 2007) (citados por Kima et al. (25). Recientemente, la superproducción de una endoquitinasa mejoró la patogenicidad en comparación con una cepa salvaje. Kima et al. (25), trataron de seleccionar, entre las tres proteínas relacionadas con la penetración y contenidas en fracciones obtenidas por cromatografía líquida, un factor para predecir la actividad insecticida en el sobrenadante de la cepa SFB-205 de B. bassiana, ensayado in vitro sobre Aphis gossypii. La disminución de la población de áfidos fue más influenciada de manera significativa por la fracción quitinasa, de forma dependiente de la dosis, por lo que concluyen que la cantidad de quitinasa pudiera ser empleada como uno de los factores para determinar la calidad del sobrenadante de la cepa SFB-205 de B. bassiana en la optimización de la producción (25). De ahí la importancia de la cuantificación de la actividad enzimática en este caso, además de los estudios de las diferentes isoformas que pueden expresarse en un aislamiento determinado.
B. bassiana ha sido aislada como colonizador endofítico de algunas especies vegetales. También se ha mostrado su efecto protector en las plantas contra fitopatógenos o inhibiendo el crecimiento micelial in vitro de fitopatógenos foliares y de suelo, entre los que se encuentran: Gaeumannomyces graminis var. tritici; Sclerotium cepivorum, Armillaria mellea, Rosellinia necatrix; Fusarium oxysporum, Botrytis cinerea, Rhizoctonia solani, Pythium ultimum, P. debaryanum, y Septoria nodorum (6, 9, 10). Aun cuando las glucanasas no han sido estudiadas en este hongo entomopatógeno, teniendo en cuenta lo anterior, resultó interesante conocer el comportamiento de la expresión de esta enzima en particular (Fig. 3).
Para esta enzima, la cepa 33 fue la que alcanzó los niveles mas elevados de expresión de la actividad, en este caso en el medio suplementado con gelatina, al séptimo día. No existen referencias de estudios de glucanasas para B. bassiana en la literatura.
Consideramos que es válido incluir este tipo de estudio empleando el medio suplementado con quitina y la determinación de quitinasa en los trabajos de selección de cepas promisorias como biocontroles de insectos. Por otra parte, sería importante realizar estudios del efecto de los aislamientos incluidos en este trabajo, sobre algunos de los fitopatógenos más importantes de Cuba, especialmente con el aislamiento 33, que fue en este estudio el más promisorio en cuanto a producción de enzimas extracelulares relacionadas con el proceso de patogénesis.
REFERENCIAS
1. Diane JH, Brett HK, Lewis MW, Keyhani NO. Surface characteristics of the entomopathogenic fungus Beauveria (Cordyceps) bassiana. Microbiology 2007;153:34483457.
2. Nicolaza Pariona, Castellanos P, León Elizabeth. Entomocidal ability of native strains Beauveria sp. on Schistocerca piceifrons peruviana (Lynch Arribalzaga, 1903). Rev. Peru biol. 2007; 14(2):253-258.
3. María del MG, Wang Amy. Efecto de biocontroladores aislados en fincas productoras de cebolla sobre la pudrición blanca (Sclerotium cepivorum). Agronomía Costarricense 2008; http://www.allbussiness.com/agronom-costarricense/41311-1.htlm (fecha de consulta 1 de abril del 2010).
4. Ownley BH, Griffin MR, Klingeman WE, Gwinn KD, Moulton. KJ, Pereira RM. Beauveria bassiana: Endophytic colonization and plant disease control. J Invertebrate Pathol. 2008;98:267-270.
5. Ownley BH, Gwinn KD, Vega FE. Endophytic fungal entomopathogens with activity against plant pathogens: ecology and evolution. Biocontrol 2010; 55(1):113-128.
6. Programa de manejo agroecológico de la broca del café (PromaBroca). INISAV-CNSV-GEAM-CENSA. Ministerio de la Agricultura. Ciudad de la Habana, Octubre del 2004.
7. Arboleda JWV, Fernando DB, Valencia AJ. Detection of Beauvericin in the entomopathogenic fungus Beauveria bassiana by using polyclonal antibodies. Rev. Colomb. Entomol. 2004; 30(2):125-130.
8. Ruiz RV. Enfoque agroecológico del empleo de entomopatógenos para el control de plagas. Conferencia dictada en el Octavo Seminario de Agroecología Agromedicina y Medio Ambiente. Tunja 22 y 23 de Noviembre del 2004.
9. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal Biochem. 1976;73:248-250.
10.Boller T, Ghri A, Mauch F, Vogeli U. Chitinase in bean leaves: induction by ethylene, purification, properties and possible function. Plant. 1983;157:22-31.
11.González Ivonne, Infante Danay, Peteira Belkis, Martínez B, Arias Yailén, González Noyma, et al. Caracterización bioquímica de aislamientos de Trichoderma spp. promisorios como agentes de control biológico. II. Expresión de actividad glucanasa. Rev Protección Veg. 2011;26(1):23-29.
12.Zheng Y, Wozniak CA. Adaptation of a 1,3 â-glucanase assay to microplate format. Biotechniques. 1997;22:922-926.
13.InfoStat versión 2009, Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. 2009.
14.Mohan MC, Reddy NP, Devi UK, Kongara R, Sharma HC. Growth and insect assays of Beauveria bassiana with neem to test their compatibility and synergism. Biocontrol Science and Technology. 2007; 17(10):1059-1069.
15.Senthilrajaa G, Ananda T, Durairajb C, Kennedyb JS, Sureshb S, Raguchandera T, et al. A new microbial consortia containing entomopathogenic fungus, Beauveria bassiana and plant growth promoting rhizobacteria, Pseudomonas fluorescens for simultaneous management of leafminers and collar rot disease in groundnut. Biocontrol Science and Technology. 2010;20(5):449-464.
16.Zimmermann G. Review on safety of the entomopathogenic fungi Beauveria bassiana and Beauveria brongniartii. Biocontrol Science and Technology. 2007;17(5/6):553-596.
17.Thakur R, Rajak RC, Sandhu SS. Biochemical and molecular characteristics of indigenous strains of the entomopathogenic fungus Beauveria bassiana of Central India. Biocontrol Science and Technology. November 2005; 15(7):733-744.
18.Talaei-Hassanloui R, Kharazi-Pakdel A, Goettel M, Mozaffari J. Variation in virulence of Beauveria bassiana isolates and its relatedness to some morphological characteristics. Biocontrol Science and Technology. 2006;16(5):525-534.
19.Zibaee A, Bandani AR. Purification and characterization of the cuticle-degrading protease produced by the entomopathogenic fungus, Beauveria bassiana in the presence of Sunn pest, Eurygaster integriceps (Hemiptera: Scutelleridae) cuticle. Biocontrol Science and Technology 2009; 19(8):797-808.
20.Zhanga YJ, Fengb MG, Fana YH, Luoa ZB, Yanga XY, Wua D, et al. A cuticle-degrading protease (CDEP-1) of Beauveria bassiana enhances virulence. Biocontrol Science and Technology. 2008; 18(6):543-555.
21.González MTG, Valencia AJ, Bustillo AEP. Incremento de la patogenicidad de Beauveria bassiana sobre Hypothenemus hampei, utilizando integumento del insecto en el medio de cultivo. Manejo Integrado de Plagas (Costa Rica). 2001; 60: 31-35.
22.Gerding-González M., France A, Sepulveda ME, Campos J. Use of chitin to improve a Beauveria bassiana alginate-pellet formulation. Biocontrol Science and Technology. 2007; 17(1):105-110.
23.Flórez MMC, López NJC, Valencia AJ. Actividad de [alfa]-amilasas del hongo entomopatógeno Beauveria bassiana cultivado en medio líquido. (Sección Agrícola) Revista Colombiana de Entomologia. 2005; http://www.accessmylibrary.com/article-1G1-153899025/actvidad-de-alfa-amilasas-html. (fecha de consulta 1 de abril del 2010).
24.Fang W, Feng J, Fan Y, Zhang Y, Bidochka MJ, St. Leger RJ, et al. Expressing a fusion protein with protease and chitinase activities increases the virulence of the insect pathogen Beauveria bassiana . J Invertebrate Pathol. 2009; 102(2): 55-159.
25.Kima JS, Rohc JY, Choid JY, Jec YH. Influence of two FPLC fractions from Beauveria bassiana SFB-205 supernatant on the insecticidal activity against cotton aphid. Biocontrol Science and Technology. 2010; 20(1):77-81.
(Recibido 15-4-2010; Aceptado 29-10-2010)


{kind=link}
{kind=link}
{kind=link}