










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión impresa ISSN 1010-2752
Rev. Protección Veg. vol.27 no.2 La Habana mayo-ago. 2012
ARTÍCULO ORIGINAL
Detección y caracterización molecular del Virus X de la Papa (PVX) en regiones productoras de papa de Colombia
Detection and molecular characterization of Potato virus X (PVX) in potato-growing regions of Colombia
J.F. Gil RamírezI, J.M. Cotes TorresII, M. Marín MontoyaI*
ILaboratorio de Biología Celular y Molecular, Facultad de Ciencias, Universidad Nacional de Colombia Sede Medellín, Calle 59A No 63-20, Medellín, Colombia.
IIDepartamento de Ciencias Agronómicas, Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia Sede Medellín, Calle 59A No 63-20, Medellín, Colombia.
RESUMEN
El cultivo de papa (Solanum tuberosum L.) se ve afectado por diferentes virus que reducen su rendimiento y la calidad del tubérculo-semilla. Determinar la distribución y la variabilidad genética de dichos patógenos, es fundamental para los programas de manejo integrado y mejoramiento genético. En este trabajo se evaluó la presencia y la variabilidad molecular de aislados de PVX en cultivos de papa de los departamentos de Antioquia, Cundinamarca, Boyacá y Nariño (Colombia). La evaluación de presencia se realizó mediante pruebas de ELISA, mientras que el análisis de variabilidad incluyó la secuenciación de una región parcial de la cápsida viral en 14 aislamientos y de porciones de los genes codificantes para las proteínas RdRp y Tgb1 para dos aislamientos. Los resultados indicaron la presencia del PVX en los cuatro departamentos con un índice de detección promedio del 8%, siendo Nariño la región con mayores niveles de detección (15%). Los análisis moleculares indicaron que la mayoría de aislamientos Colombianos se asociaron con cepas de Eurasia (Grupo I), mientras que dos aislamientos de Nariño lo hicieron con el grupo de cepas Andinas y del Reino Unido (Grupo II). La información generada en este trabajo permitirá apoyar los programas de certificación de semilla y mejoramiento genético de papa en Colombia.
Palabras clave: Alphaflexiviridae, ELISA, RT-PCR, Solanum tuberosum, variabilidad genética.
ABSTRACT
The potato (Solanum tuberosum L.) crop is strongly affected by different viruses that reduce yield and the quality of seed tubers. Knowledge of distribution and genetic variability of these pathogens is essential to support integrated pest management and breeding programs. In this research, the presence and molecular variability of PVX in potato crops was evaluated from the Colombian provinces of Antioquia, Cundinamarca, Boyacá and Nariño. The detection of this virus was performed by ELISA, while the variability was analyzed by sequencing a part of the viral capsid in 14 isolates as well as portions of genes codifying for proteins RdRp and Tgb1 for two isolates. Results indicated the presence of PVX in the four provinces with an average detection of 8%; although, in Nariño, PVX presence reached 15%. Molecular analysis indicated that most PVX Colombian isolates were associated with strains from Eurasia (Group I), while two isolates from Nariño grouped with Andean and some United Kingdom's strains (Group II). The information generated in this work will support programs of seed potato certification and breeding in Colombia.
Key words: Alphaflexiviridae, ELISA, RT-PCR, Solanum tuberosum, genetic variability.
INTRODUCCIÓN
El cultivo de la papa (Solanum tuberosum L.) representa uno de los renglones agrícolas de mayor importancia en Colombia, alcanzando cerca de 162.000 ha, un rendimiento promedio de 16,8 t.ha-1 y una producción anual que supera 2.721.396t. Este cultivo es afectado por diversas plagas y enfermedades, entre las que se destacan las polillas de los tubérculos (Tecia solanivora Povolny), el tizón tardío (Phytophthora infestans (Mont) de Bary) y diversas patologías de origen viral que incluyen Potato virus Y (PVY), Potato leafroll virus (PLRV), Potato yellow vein virus (PYVV) y Potato virus X (PVX). Diferentes trabajos indican que los problemas virales reducen los rendimientos y la calidad del tubérculo semilla en cultivos de papa de todo el mundo, razón por la cual su manejo debe constituir parte fundamental de los programas de manejo fitosanitario en este cultivo (1).
El PVX pertenece al género Potexvirus, familia Alphaflexiviridae (2) y su genoma está compuesto por una sola hebra de ARN con polaridad positiva de aproximadamente 6,4 kb. Su genoma codifica para al menos cinco marcos de lectura abiertos (ORFs) dentro de los que se encuentran la Replicasa viral (RdRp), proteínas de movimiento ubicadas en el Triple bloque de genes (Tgb) y la cápsida viral (CP). En su extremo 5' se ubica una caperuza de metil-guanosina y su extremo 3' es poliadenilado (3).
El PVX representa uno de los virus más prevalentes y que generan más pérdidas en los cultivos de papa a nivel mundial, generalmente en el rango de 5 a 20%. Este virus causa síntomas suaves, aunque también puede inducir mosaicos severos dependiendo del genotipo del hospedante, la raza viral y de las infecciones mixtas con otros virus, especialmente con potyvirus (4,5). En Colombia, el virus se ha informado afectando a los cultivos de papa en las diferentes zonas productoras del país, encontrándose en la mayoría de los casos, asociado a virus como PVY, PVS, PLRV y PYVV, siendo la interacción PVY, PVX y PLRV causante de una reducción de hasta un 61% en el rendimiento de algunas variedades nacionales (6).
Recientemente, se evaluó la presencia de virus en muestras de un grupo de 581 accesiones de papa de la Colección Central Colombiana y se encontró que el PVX estaba presente en el 13,9% de las accesiones (7). Estudios de incidencia en Colombia acoplados a análisis de rendimiento en el cultivo, demostraron que los rendimientos se reducen entre 3% y 50% dependiendo de la carga viral de los materiales debido a la sucesiva siembra de semilla contaminada, las condiciones climáticas y la capacidad de respuesta de las variedades a la infección viral (8).
Por su parte, análisis patogénicos en cultivares diferenciales de papa y tabaco portadores de los genes de resistencia Nb y Nx, determinaron la ocurrencia de al menos cuatro razas de PVX. La raza 1, no sobrepasa la resistencia conferida por ninguno de los genes; la raza 2, sólo el gen Nx; la raza 3, el gen Nb; y la raza 4, ambos genes (9). El aislamiento andino HB puede sobrepasar los dos genes y otro gen de extrema resistencia (gen Rx) (4). La proteína de la cápsida viral, es la responsable de elicitar los genes Rx y Nx, mientras que la proteína de movimiento 25 kDa elicita el gen de resistencia Nb (4,5).
Estudios de variabilidad genética, basados en secuencias tanto de genomas completos como de secuencias parciales de la cápsida viral de aislamientos de PVX de diferentes continentes, determinaron que las cepas de este virus se ubican en dos grupos que presentan una divergencia promedio del 19% en sus secuencias y que aparentemente representan dos orígenes geográficos diferentes: Eurasia (Grupo I) y Américas (Grupo II) (4,9). Sin embargo, algunos aislamientos americanos se han ubicado en el Grupo I y otros europeos en el II, lo cual evidencia introducciones como resultado del movimiento de material vegetal infectado entre dichas regiones (4).
El presente trabajo se realizó con el fin de determinar la presencia del virus PVX en cultivos de papa de cuatro departamentos de Colombia, así como la caracterización molecular de los aislamientos virales circulantes, como herramientas de apoyo a los programas de certificación de semilla, mejoramiento genético y detección temprana de los virus asociados a este cultivo en Colombia.
MATERIALES Y MÉTODOS
Evaluación de la presencia de PVX
Para esta evaluación se realizaron muestreos aleatorios en ocho cultivos de papa de diferentes variedades, ubicados de diez zonas productoras ubicadas en cuatro Departamentos de Colombia: Antioquia (zona 1: Oriente [La Unión, Sonsón], zona 2: Oriente cercano [Santuario, Marinilla, Carmen de Víboral], zona 3: Norte Antioqueño [Santa Rosa de Osos, Don Matías]), tres zonas en Cundinamarca (zona 4: Zipaquirá, zona 5: Villa Pinzón, zona 6: Occidente [Madrid, Cota]), dos zonas en Boyacá (zona 7: Turmeque, Ventaquemada, zona 8: Tunja, Siachoque) y dos zonas en Nariño (zona 9: Pasto, zona 10: Ipiales).
En cada cultivo se realizó una colección aleatoria de cuatro muestras conformadas por dos foliolos jóvenes, para generar una muestra compuesta (bulk). Se colectaron un total de 320 muestras (4 muestras x 8 cultivos x 10 zonas) y estas se evaluaron para la detección de PVX mediante pruebas de ELISA-DAS (10), utilizando anticuerpos policlonales de la compañía Agdia (Indiana, EEUU), siguiendo las instrucciones del fabricante. Los resultados colorimétricos fueron cuantificados en un equipo Multiscan (Labsystem, Finlandia), incluyendo en cada prueba un control positivo (suministrado en forma liofilizada) y un control negativo del kit. Los pozos fueron considerados con reacción positiva cuando la lectura de absorbancia a 405nm presentó un valor mínimo del doble de la lectura obtenida en el control negativo, siguiendo el criterio de Matthews (11).
Los resultados de la presencia viral se analizaron a través de un modelo lineal generalizado aleatorio considerando una distribución binomial para la variable presencia y utilizando una función de ligamiento logit. Los efectos aleatorios fueron: departamentos, zonas y cultivos dentro de zonas. Para el análisis estadístico se utilizó el procedimiento GLIMMIX del programa estadístico SAS versión 9.2.
Variabilidad genética de aislamientos de PVX
RT-PCR y secuenciación de ácidos nucleicos
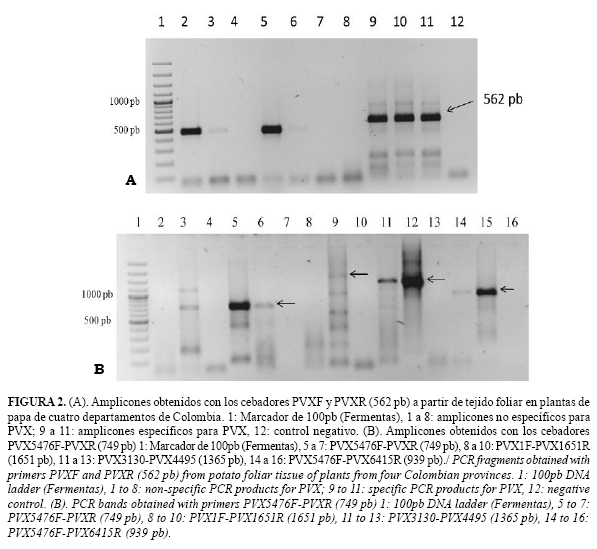
El ARN molde necesario para la técnica de RT-PCR se obtuvo mediante RNeasy plant mini kit (Qiagen, EEUU) a partir de extracciones de ARN total de folíolos de papa que fueron positivos mediante el ELISA-DAS. Para este procedimiento se maceraron 100mg de tejido foliar, utilizando 450µL de buffer RLT suministrado en el juego de reactivos y 4,5µL de b-mercaptoetanol, siguiendo las instrucciones del fabricante. Las reacciones de RT-PCR se realizaron en dos pasos con los cebadores PVXF (5'-TAG CAC AAC ACA GGC CAC AG-3') y PVXR (5'-GGC AGC ATT CAT TTC AGC TTC-3') dirigidos a las posiciones del genoma 5664-5683 y 6225-6205, respectivamente (12). Las reacciones de Retrotranscripción se realizaron con la enzima M-MuLV Transcriptasa Reversa (20U/µl) (Fermentas, Lituania) a 37°C por 60 min y el programa de PCR consistió en 95°C por 30s, seguido de 40 ciclos de 95°C por 30s, 53°C por 45s, 72°C por 1 min y una extensión final a 72°C por 5min. Los amplicones del tamaño esperado fueron purificados directamente del gel, mediante el kit QIAquick Gel Extraction (Qiagen) y secuenciados en un secuenciador ABI Prism 3730xl (PE Applied Biosystems).
Las secuencias obtenidas con cada cebador fueron editadas mediante el software BioEdit versión 6.0.6 (13), construyéndose secuencias consenso y confirmándose su identidad por comparación con secuencias disponibles en las bases de datos moleculares del NCBI (http://www.ncbi.nlm.nih.gov). El análisis de agrupamiento incluyó las secuencias obtenidas en el estudio y aquellas depositadas en el GenBank representando las variantes principales de PVX. Para esto se alinearon las secuencias mediante el software Clustal W (14) y la variabilidad genética de los aislamientos fue determinada haciendo uso del software Mega 4.0 (15) mediante la metodología Máxima Parsimonia. El soporte de la topología interna de los dendrogramas fue determinado por análisis de bootstrap con 1000 remuestreos (16).
Secuencias parciales del genoma de PVX
Con el fin de obtener un mayor nivel de información sobre las características del genoma del virus bajo estudio, adicionalmente seis pares de cebadores se evaluaron (Tabla 1) dirigidos a diferentes regiones del genoma viral de PVX (17). Para esto se seleccionaron dos aislamientos de PVX del Oriente cercano de Antioquia (Carmen de Víboral y La Ceja, seleccionados por la disponibilidad de material vegetal infectado) y se siguieron los procedimientos de extracción de ARN, RT-PCR y secuenciación descritos anteriormente. Para realizar el ensamblaje de los genomas parciales, las secuencias se editaron mediante el programa Chromas lite (18), generándose los consensos usando el programa Bioedit. Posteriormente, se verificó el marco de lectura correcto utilizando el servidor de Expasy (http://expasy.org/) y se procedió a su alineamiento con respecto al genoma completo de PVX (cepa Tula de Rusia) (EU021215). Las regiones alineadas fueron seleccionadas para evaluar sus niveles de identidad en nucleótidos (nt) y aminoácidos (aa) con la herramienta de matriz de identidad del programa Bioedit. Con las secuencias disponibles para cada aislamiento, se generó un ensamble en contigs mediante ContigExpress del software vector NTI (versión 11.0, Invitrogen).
RESULTADOS Y DISCUSIÓN
Evaluación de la presencia de PVX
Los resultados de los análisis serológicos indicaron la presencia del virus PVX en los cuatro departamentos evaluados con un promedio del 8%, siendo los cultivos de papa de Nariño los que presentaron los mayores índices de detección (15%), seguidos por Antioquia, Boyacá y Cundinamarca, para los que se obtuvieron índices del 8, 6 y 5%, respectivamente (Fig. 1A). Pasto y Oriente Cercano de Antioquia fueron en su orden las zonas con mayor presencia del virus (29 y 26%). Las demás zonas no presentaron valores superiores al 10% (Fig. 1B). Estos resultados enfatizan la importancia de perfeccionar los programas de certificación de semilla, pues a pesar que se encontraron niveles moderados, su efecto sinérgico con otros virus de papa como PVY y PLRV, puede conducir a grandes pérdidas en el rendimiento (19, 20, 21, 22).
Variabilidad genética de aislamientos de PVX
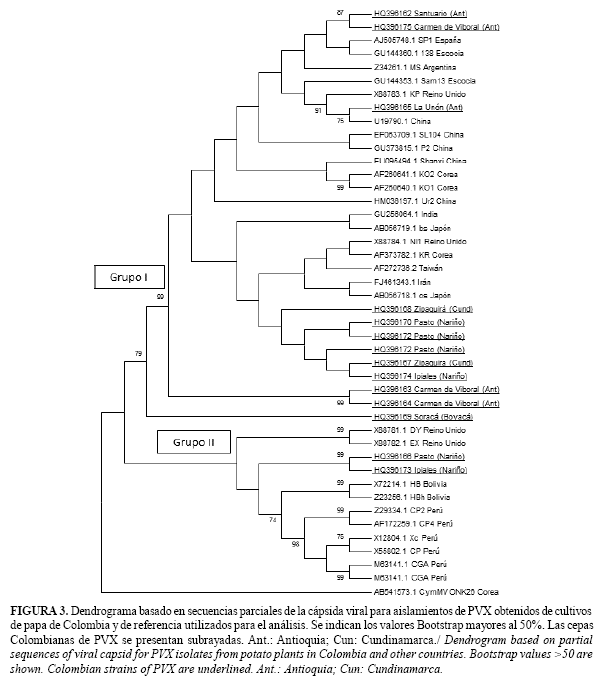
Las pruebas de RT-PCR produjeron los amplicones del tamaño esperado (~562 pb) con los cebadores PVXF y PVXR (Fig. 2). El análisis y comparación de las 14 secuencias obtenidas para los aislados Colombianos y 29 de referencia obtenidas del GenBank, representando diferentes razas y orígenes geográficos, indicó niveles de identidad para la región estudiada superiores al 77%; presentándose un grupo de 11 aislamientos colombianos que compartieron niveles superiores al 94% entre ellas y con el conjunto general de los aislamientos de referencia utilizados en el análisis (Grupo I) (Fig. 3).
Los aislamientos Colombianos (HQ396166, HQ396169 y HQ396173), de Perú (AF172259, Z29334) y Bolivia (X72214, Z23256), presentaron identidades menores con respecto al grupo principal, variando desde 78% al 87% y entre ellos del 80% al 97% (Grupo II) (Fig. 3). El nivel de identidad de la secuencia de referencia utilizada como grupo externo Cymbidium mosaic virus (CyMV, Potexvirus) (AB541573) con relación a los aislamientos de PVX fue inferior al 48%. Adicionalmente, el primer grupo contiene el aislamiento HQ396169, que difiere notablemente en su secuencia de la cápsida viral con respecto al conjunto de cepas del Grupo I, presentando una identidad con estas cercana al 87%.
Estos resultados muestran que las poblaciones de PVX en el cultivo de la papa presentan una variabilidad relativamente baja dentro de sus grupos, pero con dos variantes genotípicas claramente diferenciables mediante el análisis de secuencias.
Estudios globales de este virus han revelado la ocurrencia de dos posibles orígenes geográficos del patógeno: Eurasia y América y al menos cuatro grupos de razas del virus que pueden coinfectar paralelamente una planta (3, 4, 9). En el Grupo I del árbol presentado en este trabajo (Fig. 3), se encuentran asociados diferentes aislamientos de distintos orígenes geográficos que representan a su vez tres de las cuatro razas (1, 3 y 4), mientras que representantes de la raza 2 ocuparon principalmente el Grupo II.
A pesar de que la definición original de razas de PVX no dependía de diferencias en el genoma, sino de su reacción en plantas diferenciales, existe una relación entre las secuevariedades y las razas, por cuanto la cápsida del virus es la responsable de elicitar los genes de resistencia al virus y es de esperar que variaciones en su secuencia afectarán dicho proceso de reconocimiento por parte de los hospedantes. Es por esto que la distribución de los aislamientos Colombianos de PVX a lo largo del dendrograma y su asociación con aislamientos representativos de las cuatro razas del virus (razas 1 y 3 de Eurasia; razas 2 y 4 Americanas), plantea importantes implicaciones epidemiológicas en cuanto a la necesidad de restringir el movimiento de tubérculo semilla en el país y a la búsqueda de fuentes de resistencia que abarquen la amplitud del rango de variación que presenta este virus en los cultivos de papa de las diferentes regiones de Colombia. Adicionalmente a dicha situación, hasta el momento se desconoce en el país la presencia y los niveles de incidencia de virus informados en otros países andinos y de Centro América como PVM, PAMV, AMV, APMV, PVT, APLV, que putativamente podrían interactuar con las diferentes variantes detectadas de PVX, lo que requiere ser investigado con celeridad (23, 24).
El análisis de las secuencias parciales de los dos aislamientos de PVX provenientes de Antioquia (Carmen y Ceja) para las regiones de RdRp-3', Tgb-3' y CP, reveló identidades superiores al 95% frente a siete secuencias de referencia de aislamientos de Eurasia, y en algunos casos del 100% para aminoácidos. Las secuencias del aislamiento La Ceja corresponden a las siguientes accesiones: RdRp parcial y Tgb1 parcial (HQ433256), cápsida viral (HQ433257). Las secuencias del aislamiento Carmen corresponden a las siguientes accesiones del GenBank: RdRp parcial 3' (HQ433258), cápsida viral (HQ433255). Tanto este análisis como el de variabilidad de los aislamientos colombianos, permite aumentar la disponibilidad de secuencias del virus PVX en las bases de datos moleculares, lo que a su vez facilita que se incluyan nuestras variantes en programas de certificación de semilla de otras latitudes y de desarrollo de metodologías de diagnóstico de patógenos virales.
Los niveles de variabilidad genética que presenta el PVX en Colombia, informados en este estudio, plantean la necesidad de incluir la evaluación de diferentes genotipos virales en los programas de mejoramiento genético de variedades de papa y de manejo integrado de enfermedades virales que se realizan en el país a fin de fortalecer los mismos.
AGRADECIMIENTOS
Esta investigación se realizó gracias al apoyo económico del Ministerio de Agricultura y Desarrollo Rural de Colombia (proyecto 090-2007S4527-87-08), de la Universidad Nacional de Colombia - Sede Medellín, Fedepapa y Fritolay. Se reconoce el apoyo técnico de los miembros de los laboratorios de Biología celular y molecular de la Universidad Nacional de Colombia sede Medellín.
REFERENCIAS
1. Ministerio de Agricultura y Desarrollo Rural. Observatorio agrocadenas Colombia. Citado 30 de Agosto de 2008. Disponible: http://www.agrocadenas.gov.co.
2. International Committee on Taxonomy of Viruses (ICTV). Virus Taxonomy: 2009 Release. 2009. Citado 15 de Septiembre de 2009. Disponible en: http://www.ictvonline.org/index.asp.
3. Verchot-Lubicz J, Ye CM, Bamunusinghe D. Molecular biology of potexvirus: recent advances. J Gen Virol. 2007;88(6):1643-1655.
4. Cox A, Jones R. Genetic variability in the coat protein gene of Potato virus X and the current relationship between phylogenetic placement and resistance groupings. Arch Virol. 2010;155(8):1349-1356.
5. Yu XQ, Jia JL, Zhang CL, Li XD, Wang YJ. Phylogenetic analyses of an isolate obtained from potato in 1985 revealed potato virus X was introduced to China via multiple events. Vir Gen. 2010;40(3):447-451.
6. Guerrero O, Martínez G. Evaluación de pérdidas ocasionadas en la variedad de papa Puracé por los virus «Potato Virus X», «Potato Virus Y» y «Potato Leafroll Virus». Fitopatol Colomb. 1980;9(1):3-40.
7. Guzmán M, Román V, Franco L, Rodríguez P. Presencia de cuatro virus en algunas accesiones de la Colección Central Colombiana de papa mantenida en campo. Agron Col. 2010;28(2):225-233.
8. Sánchez de Luque C, Corzo P, Pérez O. Incidencia de virus en papa y su efecto sobre rendimientos en tres zonas agroecológicas de Colombia. Rev Lat Papa. 1991;4(1):36-51.
9. Nyalugwe E, Wilson C, Coutts B, Jones R. Biological properties of Potato virus X in potato: effects of mixed infection with Potato virus S and resistance phenotypes in cultivars from three continents. Plant Dis. 2012;96(1):43-54.
10.Clark MF, Adams AN. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J Gen Virol. 1977;34:475-483.
11.Matthews REF. Diagnosis of plant virus diseases. Canada: CRC Press. 1993; 374 p.
12.Nie X, Singh RP. A novel usage of random primers for multiplex RT-PCR detection of virus and viroid in aphids, leaves, and tubers. J Virol Methods. 2001;91(1):37-49.
13.Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser. 1999;41:95-98.
14.Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H, et al. Clustal W and Clustal X version 2.0. Bioinformatics. 2007;23(21):2947-2948.
15.Tamura K, Dudley J, Nei M, Kumar S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol Biol Evol. 2007;24:1596-1599.
16.Felsenstein J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution. 1985;39(4):783-791.
17.Ravin NV, Mardanova ES, Kotlyarov RY, Novikov VK, Atabekov JG, Skryabin KG. Complete sequencing of Potato virus X New strain genome and construction of viral vector for production of target proteins in plants. Biochemistry. 2008;73(1):44-49.
18.Technelysium. Chromas. 2.0. Citado 15 de Febrero de 2009. Disponible: http://www.technelysium.com.au/chromas_lite.html.
19.Wilson CR, Jones RAC. Occurrence of PVX strain group 1 in seed stocks of potato cultivars lacking resistance genes. Ann Appl Biol. 1995;127(3):479-487.
20.Stevenson WR, Loria R, Franc GD, Weingartner DP. Compendium of potato diseases, 2nd ed. St. Paul, Minnesota: APS Press. 2001.
21.Guzmán M, Caro M, García Y. Validación de la Técnica Serológica de Inmunoimpresión para Detección de Diferentes Virus que Afectan el Cultivo de Papa. In: Centro Virtual de Investigación de la Cadena Agroalimentaria de la Papa (CEVIPAPA). I Taller Nacional sobre Patógenos del Suelo, Virus e Insectos Plagas diferentes a Tecia solanivora. (Bogotá). 2004; 41p.
22.Zapata JL. Algunos Aspectos Sobre los Virus de la Papa en Colombia. In: Centro Virtual de Investigación de la Cadena Agroalimentaria de la Papa (CEVIPAPA). I Taller Nacional sobre Patógenos del Suelo, Virus e Insectos Plagas diferentes a Tecia solanivora. (Bogotá). 2004;24-40 p.
23.Salazar LF. Emerging and re-emerging potato diseases in the Andes. Pot Res. 2006;49:43-47.
24.Vásquez V, Montero-Astúa M, Rivera C. Incidencia y distribución altitudinal de 13 virus en cultivos de Solanum tuberosum (Solanaceae) en Costa Rica. Rev Biol Trop. 2006;54(4):1135-1141.
Recibido: 6-3-2012.
Aceptado: 21-7-2012.
*Autor de correspondencia: M. Marín Montoya. Laboratorio de Biología Celular y Molecular, Facultad de Ciencias, Universidad Nacional de Colombia Sede Medellín, Calle 59A No 63-20, Medellín, Colombia. Correo electrónico: mamarinm@unal.edu.co.

{kind=link}
{kind=link}
{kind=link}
{kind=link}