










Meu SciELO
 Serviços customizados
Serviços customizadosServiços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Protección Vegetal
versão impressa ISSN 1010-2752
Rev. Protección Veg. vol.28 no.3 La Habana set.-dez. 2013
ARTÍCULO ORIGINAL
Inducción de mecanismos de defensivos en plantas de tomate (Solanum lycopersicum L. cv. Amalia) con el uso de aceites esenciales1
Induction of defence mechanisms in tomato (Solanum lycopersicum L. cv. Amalia) plants by essential oils
Ivonne González, Yailén Arias, Yaíma Sánchez, Miriam M. Rojas, Esteban González, Oriela Pino, Belkis Peteira
Dirección de Sanidad Vegetal. Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, Mayabeque, Cuba. Correo electrónico: marquetti@censa.edu.cu.
RESUMEN
Se estudió el efecto de diferentes aceites esenciales en la inducción de mecanismos defensivos en tomate (Solanum lycopersicum L. cv. Amalia). Las plantas se asperjaron con diferentes aceites e inductores como el Bion, y se tomaron muestras de hojas a las 24, 72 y 144 horas posteriores a la aspersión. Se determinó la concentración de proteínas totales, así como la actividad enzimática específica de peroxidasas, polifenoloxidasas, fenilalanina amonio liasas y quitinasas. Los aceites esenciales obtenidos a partir de subproductos de la industria de cítricos y de melaleuca (Melaleuca quinquenervia (Cav) S.T. Blake) indujeron las mayores actividades enzimáticas de peroxidasas, polifenoloxidasas y quitinasas. El ácido jasmónico y los fosfolípidos fueron los responsables de la inducción de fenilalanina amonio liasas.
Palabras clave: ácido jasmónico, Bion, Fitomas, fosfolípidos.
ABSTRACT
In this work, the effect of different essential oils was evaluated using tomato cv. Amalia plants. The plants were sprayed with different oils and inductors as Bion, and leave samples were taken at 24, 72 and 144 hours after spray. Total protein concentration and specific enzymatic activities of peroxidases, polyphenoloxidases, phenylalanine ammonia lyase and chitinases were determined. The essential oils obtained from citric fruit scraps and Melaleuca quinquenervia (Cav) S.T. Blake induced higher peroxidases, polyphenoloxidases and chitinases enzymatic activities. Jasmonic acid and phospholipids were responsible for phenylalanine ammonia lyase induction.
Key words: jasmonic acid, Bion, Fitomas, phospholipids.
INTRODUCCIÓN
Diversos productos aplicados usualmente para el control de plagas en los cultivos, constituyen sustancias químicas nocivas para el ambiente. La explotación de los mecanismos de inducción de defensa natural se presenta como una alternativa, no convencional y ecológicamente amigable, para la protección de las plantas, contribuyendo a la reducción de la aplicación del control químico y al desarrollo de una agricultura sostenible.
La resistencia inducida se puede definir como un incremento en la expresión de mecanismos de defensas naturales de plantas contra varios tipos de patógenos, provocada por varios factores: patógenos causantes de la reacción necrótica de hipersensibilidad, cepas patógenicas avirulentas o atenuadas, elicitores de origen patógenico (glucanos, proteínas, lípidos, otros.), elicitores abióticos, incluyendo productos químicos sintéticos inofensivos, tales como el ácido 2,6-dicloroisonicotínico (INA), ácido b-aminobutírico (BABA), benzotiadiazol (BTH, también conocido como Bion o ASM) (1), y recientemente se señaló que metabolitos secundarios, obtenidos a partir de plantas, pueden ser utilizados como inductores de resistencia (2).
Hasta la fecha, numerosas plantas fueron utilizadas por los campesinos cubanos como repelentes y/o materias primas para la preparación de extractos de manera artesanal y se demostró la actividad plaguicida de más de 60 plantas en condiciones de laboratorio, semicontroladas y campo (2).
Desde el punto de vista químico, se lograron resultados promisorios con alcaloides, terpenoides, cumarinas y aceites esenciales. Estos últimos evidenciaron propiedades antibacterianas, antifúngicas, acaricidas, repelentes e insecticidas, entre los que se cuentan los obtenidos a partir de Citrus sinensis (L) Osbeck, Melaleuca quinquenervia (Cav) S.T. Blake y Piper aduncum subsp. ossanum (C. DC) Saralegui. Como plaguicidas botánicos se pueden aplicar en cultivos protegidos, viveros, tratamiento de semillas, manejo de plagas del almacén; entre otros; sin embargo, productos novedosos pueden desarrollarse, utilizando metabolitos secundarios en mezclas con productos fitosanitarios y como inductores de resistencia (2).
La capacidad de los productos naturales de inducir resistencia a plagas en las plantas es poco conocida. No obstante, estudios previos indicaron que el extracto acuoso de hojas de Ocimum gratissimum L. a concentraciones del 10, 25, 40 y 50% (w/v) indujo la producción de fitoalexinas en cotiledones de soya y mesocotilos de sorgo, así como resistencia sistémica en pepino contra Colletotrichum lagenarium (Berk. & Mont.) Arx, reflejado por la reducción en la incidencia de la enfermedad y por la producción de quitinasas (3). De igual modo, recientemente se demostraron las potencialidades del aceite de Syzygium aromaticum (L.) Merr. et Perry como inductor de resistencia en plantas de tomate cv. Santa Cruz Kada contra Xanthomonas vesicatoria (ex Doidge) Vauterin et al. agente causal de la Mancha Bacteriana del tomate (4).
El objetivo de este trabajo fue estudiar la inducción de enzimas de resistencia por parte de diferentes aceites esenciales en plantas de tomate cv. Amalia.
MATERIALES Y MÉTODOS
Material vegetal
El estudio se realizó en el Centro Nacional de Sanidad Agropecuaria (CENSA), (Mayabeque, Cuba). Las plantas de tomate cv. Amalia se sembraron en macetas que contenían suelo previamente esterilizado durante cuatro horas a 120ºC. Se mantuvieron en condiciones semicontroladas en casa de cristal, a una temperatura de aproximadamente 23ºC, humedad relativa entre 80-85% y fotoperíodo natural.
Transcurridos 30 días desde la germinación, las plantas se asperjaron con las siguientes soluciones: Bion® (1g.L-1, según instrucciones del manufacturador, Novartis, Alemania), Fitomas® (1ml.L-1), Biojás (ácido jasmónico, 10mg.L-1 según instrucciones del manufacturador, Instituto Central de Investigaciones de Derivados de la Caña de Azúcar-ICIDCA), fosfolípidos (0,1g.L-1) y aceites esenciales de tres plantas.
Los aceites esenciales se extrajeron por hidrodestilación con equipo Clevenger durante tres horas, a partir de P. aduncum subsp. ossanum (C. DC.) Saralegui, M. quinquenervia (Cav) S.T. Blake y de subproductos industriales de cítricos (Citrus spp.), según lo establecido en la norma ISO 65-71:84 (5).
Se evaluaron las concentraciones de P. aduncum subsp. ossanum (0,19% v/v) (34PL), M. quinquenervia (0,19% v/v) (99PL) y de subproductos de cítricos (0,39% v/v) (114PL), los cuales fueron previamente disueltos en placebo (116,7g.L-1 de maltodextrina y 233,3g.L-1 de goma arábiga) y fosfolípidos (0,1g.L-1). Las plantas asperjadas con agua y placebo se establecieron como controles. Los muestreos se realizaron a las 24, 72 y 144 horas, después de la aspersión con los diferentes tratamientos.
Para la extracción de proteínas, se maceró el material vegetal foliar (5g) en nitrógeno líquido y se homogenizó con solución tampón de extracción (acetato de sodio 0,1 M; pH 5,2; que contenía 0,1ml de b-mercaptoetanol en 100 ml de solución de extracción), en proporción 2:1 (ml: gramos de masa fresca). El homogenado se agitó en zaranda durante 30 minutos en baño con hielo. Posteriormente se filtró a través de cuatro capas de gasa y se centrifugó a 6440g a 4ºC durante 30 minutos en centrifuga refrigerada (Beckman). El extracto de proteínas fue almacenado a -20ºC.
Se utilizó el sobrenadante para la determinación de la concentración de proteínas totales por el método descrito por Bradford (6), se realizaron las lecturas de la absorbancia a 595 nm del complejo proteína-Azul de Coomasie G-250. El cálculo de la concentración de la muestra se realizó a partir de una solución patrón de 1mg.ml-1 de albúmina de suero bovina para confeccionar la curva patrón.
La detección de la actividad peroxidasa, se realizó según el método continuo descrito por Frick et al. (7). Se utilizaron como sustratos el guayacol y el peróxido de hidrógeno. Se determinó la velocidad de la reacción de oxidación del guayacol por la enzima, en presencia de peróxido de hidrógeno, se registraron los valores de absorbancia a 470 nm durante dos min. cada 15 segundos.
La actividad enzimática de las polifenoloxidasas se determinó a través del método continuo según Alexander, se empleó como sustrato de la enzima el pirogallol (8). La velocidad de la reacción se determinó registrando la densidad óptica a una longitud de onda de 420 nm durante dos min. por intervalos de 15 segundos.
La actividad fenilalanina amonio liasa fue cuantificada según el método discontinuo propuesto por Paynet et al. (9), que utiliza como sustrato fenilalanina. En este caso, la velocidad de la reacción se calculó a partir de las lecturas de absorbancia a 275 nm, donde se midió el ácido cinámico formado. Este reactivo fue utilizado para hacer la curva patrón, a partir de una solución estándar de 1 mg.ml-1.
Se determinó la actividad quitinasa con 0,2 ml de quitina coloidal 10 mg.ml-1, preparada acorde a Boller et al. (10). La curva patrón fue sometida al mismo procedimiento que las muestras y se utilizó N-acetilglucosamina, a partir de una solución madre de 1mg.ml-1. La lectura se realizó a 585 nm.
Para determinar los tratamientos que indujeron los mayores niveles de actividades enzimáticas se empleó el análisis de varianza simple. La comparación entre las medias se realizó con el uso de la prueba de comparación de rangos múltiples de Duncan en los casos donde existieron diferencias (p<0,05) utilizando el paquete estadístico InfoStat versión 2011 (11).
RESULTADOS Y DISCUSIÓN
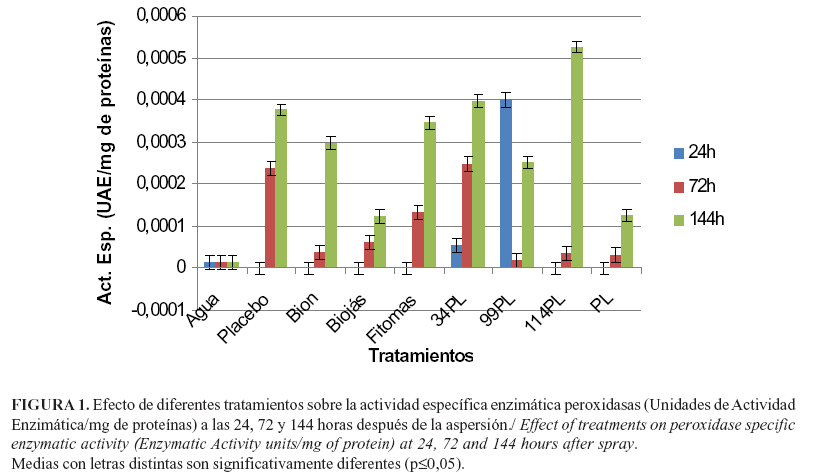
Los tratamientos produjeron inducción de la actividad enzimática de los sistemas ensayados. En el caso de las peroxidasas (POs), los mayores niveles de actividad se expresaron en las plantas tratadas con el aceite extraído a partir de subproductos de la industria de cítricos. Los aceites obtenidos a partir de P. aduncum subsp. ossanum y M. quinquenervia también produjeron niveles elevados de actividad PO, pero en esta última se lograron los mayores valores de expresión a las 24 horas y posteriormente disminuyó la actividad específica de dicha enzima (Figura 1).
Durante los diferentes días de análisis, en los controles tratados con el placebo se incrementaron los valores en la actividad PO, alcanzando a las 144 horas niveles similares a los de 34PL, 99PL y Fitomas®. En el caso del placebo, sus componentes maltodextrina (constituida por un oligosacárido de glucosa obtenido a partir de la hidrólisis enzimática parcial del almidón de maíz (12)), y goma arábiga (mezcla heterogénea de polisacáridos con cantidades variables de D-galactosa, L-arabinosa, L-ramnosa y algunos ácidos derivados como el ácido D-glucorónico o el 4-O-metil-D-ácido glucorónico (13)), no se señalaron propiedades como inductores de resistencia en las plantas. Sin embargo, se señaló que algunas moléculas como pequeños péptidos, lipoligosacáridos y lipopolisacáridos pueden elicitar respuestas defensivas basales o inespecíficas en las plantas (14), lo que pudiera ser la explicación de este efecto observado, aspecto que debe ser objeto de estudios futuros.
Las POs catalizan la liberación de proteínas estructurales de forma insoluble provenientes de la pared celular, en presencia de H2O2, el que además, induce la formación de polímeros de lignina vía actividad peroxidasa (15).
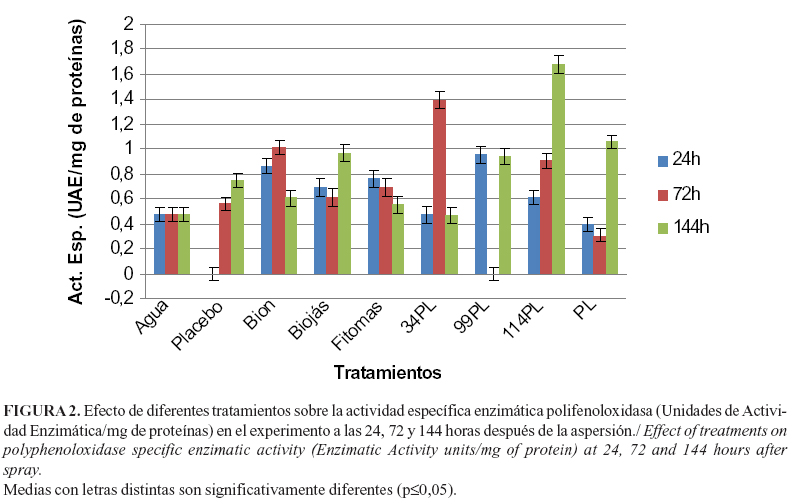
Por su parte, las plantas tratadas con 114PL expresaron también los niveles más elevados de actividad polifenoloxidasas (PPOs), seguidas de aquellas que fueron asperjadas con la mezcla que contenía 34PL (Figura 2). Los restantes tratamientos mostraron niveles inferiores de actividad específica enzimática PPOs, con diferencias significativas con los tratamientos, donde se empleó 114PL y 34PL.
Las PPOs constituyen una clase de enzimas que utilizan el oxígeno molecular para la oxidación de mono y O-difenoles a O-dihidroxiquinonas. Estas son moléculas muy reactivas que se enlazan covalentemente a los grupos nucleofílicos NH2 y SH de los aminoácidos y proteínas, reduciendo la disponibilidad de los aminoácidos esenciales a los herbívoros y/o la digestibilidad de las proteínas, o interfiriendo directamente con las enzimas. Las PPOs también están implicadas en la resistencia de las plantas a las bacterias fitopatógenas. La sobreexpresión de una PPO de papa (Solanum tuberosum L.) en tomate cultivado, mostró que estas plantas transgénicas fueron más resistentes a la bacteria fitopatógena Pseudomonas syringae van Hall (16).
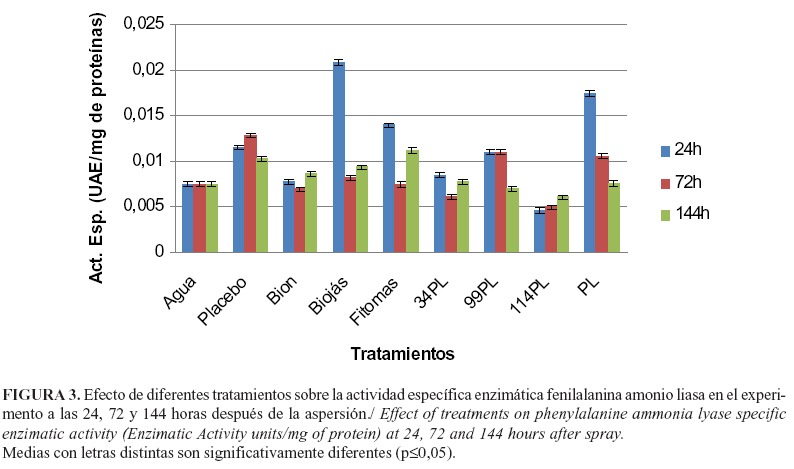
En el caso de la enzima fenilalanina amonio liasa (PAL), los valores más significativos se alcanzaron en las plantas asperjadas con Biojás, seguidos de las tratadas con fosfolípidos y Fitomas® con diferencias significativas entre ellos y con los restantes tratamientos (Figura 3).
Estos resultados no coinciden con los de Inbar et al. (17), quienes plantearon que el BTH fue el mejor entre muchos inductores probados en plantas de tomate contra varios patógenos bacterianos y fúngicos. De igual modo, no concuerdan con los de Peteira et al. (18) quienes encontraron que la aplicación de Fitomas® y Bion® fue efectiva en la disminución de las poblaciones de Steneotarsonemus spinki Smiley en arroz, con los mejores resultados en el caso de la aplicación de este último producto. Los altos niveles de actividad fenilalanina amonio liasa encontrados a las 24 horas en las plantas tratadas con Biojás, se corresponden con los obtenidos por Sharan et al. (19) cuando trataron suspensiones de células de tabaco con metil jasmonato, un derivado esterificado del ácido jasmónico.
El incremento en la actividad de esta enzima de la vía de los fenilpropanoides aumenta la producción de ácido trans-cinámico a partir de fenilalanina, el cual es trasformado a través de otras dos reacciones enzimáticas hasta para-cumaril CoA; precursor de un amplio grupo de productos que incluyen la lignina, los fenilpropenos y los flavonoides (16). La lignina confiere impermeabilidad, resistencia mecánica y protege a la planta contra la degradación producida por muchos patógenos (20). Los fenilpropenos cumplen una función dual, pues están involucrados en la defensa de la planta contra ataque de herbívoros y permiten la atracción de los polinizadores. Por su parte, los flavonoides, protegen a la planta de las radiaciones UVA y UVB (16).
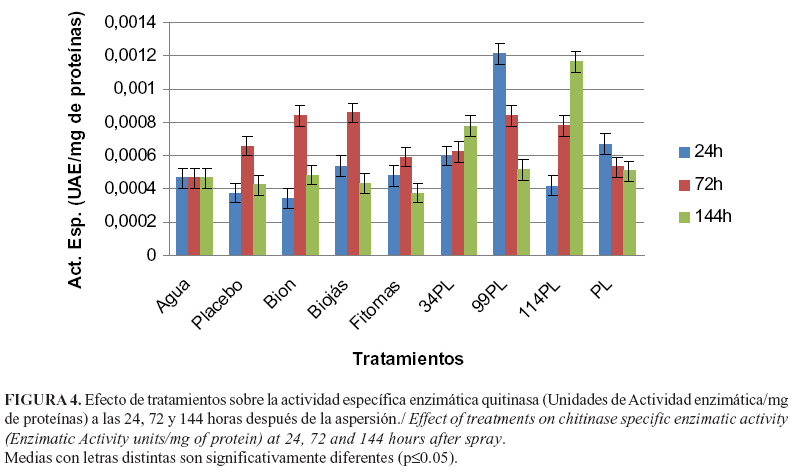
Los tratamientos empleados provocaron un incremento en la actividad específica quitinasa en la mayoría de los casos (Figura 4). Las plantas tratadas con 99PL y 114PL produjeron los mayores valores de actividad enzimática, a las 24 y 72 horas de asperjadas respectivamente. La elevada expresión de esta enzima confiere protección contra el ataque de patógenos con quitina en su estructura (21).
Los resultados evidenciaron que los aceites esenciales estudiados poseen potencialidades como inductores de mecanismos de defensa en tomate, pudiendo propiciar resistencia frente a un amplio grupo de fitopatógenos y plagas, aspecto que debe ser objeto de investigaciones futuras donde se evalúen los mecanismos de resistencia en presencia de organismo plaga. Esta habilidad de los aceites esenciales, junto a características como elevada disponibilidad, buena eficacia, amplio espectro de acción, baja residualidad y toxicidad, los convierte en productos promisorios para el manejo de plagas en nuestro país (22, 23).
AGRADECIMIENTOS
Agradecemos a la Dra. Georgina Michelena del ICIDCA por proporcionarnos el Biojás y el Fitomas® para la realización del estudio. También agradecemos de forma especial a la Dra. Mayra Rodríguez por la cuidadosa revisión de este manuscrito. Se agradece a los técnicos Belkis Pino y Cecil González.
REFERENCIAS
1. Zuluaga CM, Patiño LF, Collazos JC. Integración de inducción de resistencia con bacterias quitinolíticas en el control de la Sigatoka Negra (Mycosphaerella fijiensis Morelet) en banano. Rev Fac Nal Agr Medellín. 2007;60(2):3891-3905.
2. Pino O, Sánchez Y, Rojas MM. Plant secondary metabolites as alternatives in pest management. II: An overview of their potential in Cuba. Rev Protección Veg. 2013;28(2):95-108.
3. Tavares F, Freitas KR, Stangarlin JR, Ferrarese ML, Scapim CA, Bonaldo SM. Induction of plant defense responses by Ocimum gratissimum L. (Lamiaceae) leaf extracts. Summa Phytopathol. 2009;35(3):191-195.
4. Lucas GC, Alves E, Borges-Pereira R, Zacaroni AB, Perina FJ, de Souza MR. Indian clove essential oil in the control of Tomato Bacterial Spot. J Plant Pathol. 2012;94(1):45-51.
5. International Standarization Organization. ISO 65-71. Spices, condiments and herbs. Determinationof volatile oil content. 1984. (Norma ISO).
6. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal Biochem. 1976;73:248-250.
7. Frick F. Oxidative enzymes. Physiol Plant Pathology. 1976;4:617-627.
8. Alexander AG. Sucrose enzyme relationship in immature sugar cane. J Agric Univ P Rico. 1964;4813:165-231.
9. Paynet M, Martin C, Girand M. Activité phenylalanine ammniae lyase et hypersensibilite an virus de la mosaique du tabac. Acad Sci Paris. 1971;273:537-539.
10.Boller T, Ghri A, Mauch F, Vogeli U. Chitinase in bean leaves: induction by ethylene, purification, properties and possible function. Plant. 1983;157:22-31.
11.Di Rienzo JA, Casanoves F, Balzarini MG, González L, Tablada M, Robledo CW. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, InfoStat versión 2011. Argentina. URL http://www.infostat.com.ar.
12.Livesey G, Tagami H. Interventions to lower the glycemic response to carbohydrate foods with a low-viscosity fiber (resistant maltodextrin): meta-analysis of randomized controlled trials. Am J Clin Nutr. 2009;89:114-125.
13.Gracia O. La goma arábiga: una maravilla natural que beneficia a todos: http://depa.fquim.unam.mx/amyd/archivero/GomaArabiga_1838.pdf (Consultado 16-08-2013).
14.Conrath U. Priming of induced plant defense responses. Adv Bot Res. 2009;51:361-395.
15.de la Noval B, Pérez E. Inducción de respuestas de defensa. En: Ed. Pérez MM. Instituto Nacional de Ciencias Agrícolas, Inducción de respuestas de defensa en la interacción planta microorganismos (micorrizas arbusculares y Rhizobium). Cuba. 2004, pp: 5.
16.Glas JJ, Schimmel BCJ, Alba JM, Escobar-Bravo R, Schuurink RC, Kant MR. Plant glandular trichomes as targets for breeding or engineering of resistance to herbivores. Int J Mol Sci. 2012;13:17077-17103.
17.Inbar M, Doostdar H, Sonoda KM, Leibee GL, Mayer RT. Elicitors of plant defensive systems reduced insect densities and disease incidents. J Chem Ecol. 1998;24:135-149.
18.Peteira B, Fernández A, Rodríguez H, González E. Efecto del Bion y del Fitomas como inductores de resistencia en plantas de arroz infestadas con Steneotarsonemus spinki. Rev Protección Veg. 2008;23(1):32-37.
19.Sharan M, Taguchi G, Gonda K, Jouke T, Shimosaka M, Hayashida N, et al. Effects of methyl jasmonate and elicitor on the activation of phenylalanine ammonia-lyase and the accumulation of scopoletin and scopolin in tobacco cell cultures. Plant Sci. 1998;132(1):13-19.
20.Peteira B, Dueñas F, Arias Y, Martínez Y, Pino O. Caracterización de materiales promisorios de tomate obtenidos en el programa de mejoramiento para la resistencia al TYLCV. Fitopatología. 2008;43(3):105-119.
21.González I, Infante D, Martínez B, Arias Y, González N, Miranda I, et al. Inducción de quitinasas y glucanasas en cepas de Trichoderma spp. Promisorias como agentes para el control biológico. Biotecnología Aplicada. 2012;29:7-11.
22.Pino O, Sánchez Y, Rodríguez H, Correa TM, Demedio J, Sanabria JL. Caracterización química y actividad acaricida del aceite esencial de Piper aduncum subsp. ossanum frente a Varroa destructor. Rev Protección Veg. 2011;26(1):52-61.
23.Sánchez Y, Pino O, Jorge-Lazo F, Abreu Y, Naranjo E, Iglesia A. Actividad promisoria de aceites esenciales de especies pertenecientes a la tribu Pipereae frente a Artemia salina y Xanthomonas albilineans. Rev Protección Veg. 2011;26(1):45-51.
Recibido: 30-8-2013.
Aceptado: 1-11-2013.
1Resultados obtenidos en el marco del proyecto: Diagnóstico y manejo de plagas en granos con énfasis en el desarrollo y uso de productos bioactivos (Programa Nacional de Salud Animal y Vegetal, Cuba).

{kind=link}
{kind=link}
{kind=link}
{kind=link}