










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión impresa ISSN 1010-2752
Rev. Protección Veg. vol.29 no.3 La Habana sep.-dic. 2014
ARTÍCULO ORIGINAL
Distribución del Virus de la hoja amarilla de la caña de azúcar en Cuba
Distribution of the Sugarcane yellow leaf virus in Cuba
Osmany de la C. Aday DíazI, María de la Luz La O-HechavarríaII, María de los Ángeles Zardón NavarroII, Eida Rodríguez LemaII, José María Mesa LópezII, Yaquelin Puchades IzaguirreIII, Javier Delgado PadrónIV, Félix René Díaz MujicaI
IEstación Territorial de Investigaciones de la Caña de Azúcar Centro Villa Clara. Autopista Nacional Km 246, Ranchuelo, C.P. 53100, Villa Clara, Cuba. Correo electrónico: directora@dcitma.vcl.cu.
IIInstituto de Investigaciones de la Caña de Azúcar (INICA). Carretera CUJAE Km 1 1/2. Boyeros, C.P. 19390. La Habana. Cuba. E-mail: mesa@inica.azcuba.cu; lao@inica.zcuba.cu.
IIIEstación Territorial de Investigaciones de la Caña de Azúcar Oriente-Sur.
IVEstación Provincial de Investigaciones de la Caña de Azúcar Mayabeque.
RESUMEN
El presente estudio tuvo el propósito de determinar la distribución del Virus de la hoja amarilla (Sugarcane yellow leaf virus, SCYLV), en diferentes provincias del país y las variantes genéticas presentes. Se analizaron 35 campos (348 ha), de los cuales se tomaron 525 muestras de hojas en 13 cultivares comerciales. La detección del virus se realizó por Inmunoimpresión de Tejidos y por Reacción en Cadena de la Polimerasa con Transcripción Reversa (RT-PCR). Las muestras que resultaron positivas fueron utilizadas para identificar las variantes genéticas por RT-PCR con cebadores específicos. En la mayoría de las provincias fue alta la incidencia de síntomas asociados a esta enfermedad (82,29%). Se detectó, por primera vez, la presencia de este patógeno en áreas comerciales de todas las provincias que cultivan la caña de azúcar en Cuba. En la mayoría de los cultivares infectados, se observó el mayor grado de severidad de los síntomas, asociada a esta enfermedad. Se determinó la presencia de tres variantes genéticas de SCYLV: BRA-PER, CUB y REU. También se observaron infecciones mixtas de los genotipos BRA-PER + CUB y BRA-PER + REU.
Palabras clave: hoja amarilla de la caña de azúcar, SCYLV, distribución geográfica, variantes genéticas.
ABSTRACT
The present study had the purpose of determining the distribution of the Sugarcane yellow leaf virus (SCYLV) and its genetic variants occurring in different provinces of the country. Thirty five fields were examined (348 ha), and 525 samples of leaves were taken in 13 commercial cultivars. Detection of the virus was carried out by Tissue Blot Immunoassay (TBIA) and the molecular analysis by Reverse Transcription- Polymerase Chain Reaction (RT-PCR). The positive samples were used to identify the genetic variants by RT-PCR with specific primers. In most of the provinces, the incidence of symptoms associated with this disease (82.29%) was high. This is the first time that this pathogen has been detected in commercial areas of all the provinces where sugarcane is cultivated in Cuba. In most of the infected cultivars, the highest severity grade of the symptoms associated with this disease was observed. The presence of three genetic variants of SCYLV was determined: BRA-PER, CUB and REU. Mixed infections of the genotypes, BRA-PER + CUB and BRA-PER + REU, were also observed.
Key words: Sugarcane yellow leaf virus, SCYLV, geographical distribution, genetic variants.
INTRODUCCIÓN
La enfermedad «hoja amarilla de la caña de azúcar» es causada por el Virus de la hoja amarilla de la caña de azúcar (Sugarcane yellow leaf virus, SCYLV), miembro de la familia Luteoviridae, género Polerovirus (1, 2). Se considera una de las principales causas de origen biótico que pueden reducir sustancialmente el rendimiento de la caña de azúcar, junto al raquitismo de los retoños [Leifsonia xyli subsp. xyli (Davis et al.) Evtushenko]; carbón [Sporisirium scitamineum (Sydow)], escaldadura foliar [Xanthomonas albilineans (Ashby) Dowson] y roya parda [Puccinia melanocephala H. Sydow y P. Sydow) (3).
El síntoma más característico de SCYLV es el amarillamiento del raquis o nervadura central de la hoja por el envés y de la lámina foliar; necrosis desde el ápice de la hoja; expansión gradual del amarillamiento desde la nervadura hacia los bordes y acortamiento de los entrenudos terminales (4). En algunos cultivares se produce también una coloración rojiza sobre el haz de la nervadura central de las hojas y, a su vez, presentan amarillamiento en el envés. Estos síntomas no son específicos y pueden tener origen biótico y abiótico (5). Los grados Brix en jugo de la nervadura central de hojas infectadas pueden ser de dos a tres veces más altos en plantas con síntomas que en plantas sanas (6).
Dos patógenos están asociados a los síntomas de amarillamiento, un virus y un fitoplasma (1), cuyos síntomas son idénticos (7). El fitoplasma del amarillo de la caña de azúcar (Sugarcane yellows phytoplasma o SCYP) también está asociado con los síntomas similares de amarillamiento en caña de azúcar (1, 8), estos incrementan su severidad en el período seco de maduración y cosecha de la caña (9).
El virus se propaga por la plantación de trozos de semilla infectada y se transmite por los áfidos Melanaphis sacchari Zehntner, Rhopalosiphum maidis Fitch, Rhopalosiphum rufiabdominalis Sasaki y Sipha flava Forbes (Homoptera: Aphididae) (1, 10, 11).
Están informadas diez variantes genéticas principales, descritas y nombradas de acuerdo al origen geográfico donde fueron determinados por primera vez (BRA, CUB, COL, IND, HAW, PER, REU, CHN1, CHN2, CHN3) (12, 13, 14, 15, 16, 17).
SCYLV está presente en más de 35 países del mundo (18) y su presencia en Cuba se estableció en 1999 a través del análisis de muestras de cañas mantenidas en la colección de germoplasma ubicada en la provincia de Matanzas que expresaron síntomas (19). Posteriormente, en muestras de esa colección, se determinó también la presencia del virus (12, 20, 21); sin embargo, no se ha realizado, hasta la fecha, ninguna prospección en áreas comerciales.
El presente estudio tuvo el propósito de determinar la distribución del virus de la hoja amarilla en diferentes provincias del país y las variantes genéticas presentes.
MATERIALES Y MÉTODOS
La prospección se realizó durante los meses de enero y febrero de 2011, período durante el cual se manifiestan los síntomas con mayor intensidad (9) y abarcó 13 provincias (Artemisa, Mayabeque, Matanzas, Villa Clara, Cienfuegos, Sancti Spíritus, Ciego de Ávila, Las Tunas, Camagüey, Holguín, Granma, Santiago de Cuba y Guantánamo). Se muestrearon al azar 35 campos de caña (de dos a tres campos por provincia), entre 10 y 12 meses, lo que permitió evaluar 13 cultivares comerciales.
Las muestras (con síntomas o asintomáticas) fueron colectadas al azar en cinco puntos de cada campo (Noroeste, Noreste, Medio, Suroeste, Sureste). Se seleccionaron al azar tres tallos de plantas diferentes en cada punto de muestreo (15 por campo), para un total de 525 muestras, y se realizó el diagnóstico visual y la lectura del brix refractométrico del jugo extraído del tercio basal de la primera hoja con cuello visible (hoja +1).
Para establecer el grado de la enfermedad, se utilizó la escala de 4 valores (Tabla 1) propuesta por Chinea et al. (22).
A partir de las 525 muestras de campo, para la detección del virus se seleccionaron al azar muestras de las diferentes provincias, 89 hojas +1 en total, estas fueron conservadas a 4°C hasta su análisis en el laboratorio. La detección del patógeno se realizó por Inmunoimpresión de Tejidos (Tissue Blot Immunoassay o TBIA). Se agregó a la membrana un anticuerpo específico policlonal de conejo anti-SCYLV, gentilmente donado por el Dr. Jean-Claude Girard, CIRAD, Francia y se siguió el protocolo de Schenk et al. (23).
La detección molecular se realizó a 18 muestras positivas por TBIA (seleccionadas al azar), mediante Reacción en Cadena de la Polimerasa con Transcripción Reversa (RT-PCR), con cebadores genéricos para SCYLV: ScYLVf1 (GACAGACTCGGCCAGTGGTC GTG) y ScYLVr1 (GTAAGCCATTGTTGAACGCTGCG) (2). Las muestras que resultaron positivas con cebadores genéricos para SCYLV, se utilizaron para identificar tres variantes genéticas del virus por RT-PCR con los cebadores específicos descritos por Ahmad et al. (12). Los cebadores específicos fueron: BRA-PERF: AACTGCTGCGTCAGGCCCA y R: GACGAGCTTG CGTTGTTTTTCT); CUB: F: GTGCTTCTCCCGG CGGTTCACT y R: ATTCGAGAACAACCTCCGCCTC); REU: F: CAAGCTTCTAGCGGGAATC y R: CAGTT GCTCAATGCTCCACG). El tamaño de los productos esperados de la amplificación de SCYLV fue de 362, 450 y 905 pb, para las variantes genéticas BRA-PER, CUB y REU, respectivamente.
La incidencia de los síntomas (expresado en porcentaje), en muestras de campos con diferente número de cosechas, se determinó por la siguiente fórmula:

Para determinar las diferencias entre el número de cortes o cosechas, en cuanto a la incidencia de síntomas de la hoja amarilla en las muestras evaluadas, se empleó la prueba de Kruskal-Wallis. Para establecer la asociación entre la presencia de síntomas de amarillamiento y los resultados de la prueba TBIA para SCYLV, se elaboró una tabla de frecuencias (2 x 2) y se realizaron las pruebas de Chi-cuadrado y Fisher exacta.
RESULTADOS
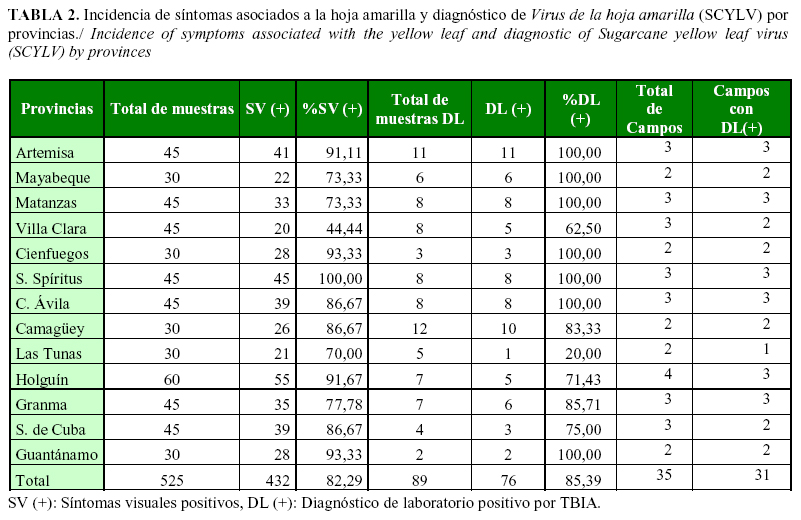
Los síntomas relacionados con la presencia de la enfermedad causada por SCYLV, se observaron en todas las provincias y campos evaluados en 12 de las 13 provincias evaluadas. Sólo en Villa Clara, un campo del cultivar C87-252 resultó asintomático, no obstante, se determinó la presencia del virus en el mismo por RT- PCR. Los resultados de laboratorio por TBIA revelaron la presencia del virus en el 100% de los campos con diagnóstico visual de las provincias Artemisa, Mayabeque, Matanzas, Villa Clara, Cienfuegos, Sancti Spíritus, Ciego de Ávila, Camagüey, Granma, Santiago de Cuba y Guantánamo. En las provincias Las Tunas y Holguín, la presencia del virus se detectó en el 50% y 75% de los campos evaluados, respectivamente.
Con la excepción de Villa Clara, la incidencia de síntomas asociados a la hoja amarilla fue superior al 70% en la mayoría de las provincias (Tabla 2). Se observó alta incidencia de los síntomas asociados a la hoja amarilla (SCYLV) desde caña planta, sin diferencias significativas en relación con otros campos con diferente número de cosechas (Tabla 3).

El 85,39% de las muestras analizadas en el laboratorio, para determinar la presencia del virus por TBIA, resultaron positivas para SCYLV (Tabla 2). En diez de las provincias muestreadas se detectó el virus en todos los campos (Artemisa, Mayabeque, Matanzas, Cienfuegos, Sancti Spíritus, Ciego de Ávila, Camagüey, Granma, Santiago de Cuba y Guantánamo).
Se constató la presencia del virus en 12 muestras asintomáticas (13,48%), mientras otras 11 (12,36%) presentaron síntomas de la enfermedad; sin embargo, resultaron negativas por TBIA.
En 12 de los 13 cultivares evaluados, se observaron síntomas de la enfermedad (Tabla 4) y en la mayoría de los casos se confirmó la presencia del SCYLV. En el cultivar C87-252 no se observaron síntomas; no obstante, las muestras sometidas a TBIA revelaron la presencia del virus. Contrariamente en muestras con síntomas del cultivar C88-380, el diagnóstico serológico no detectó la presencia del virus.
No se encontró asociación estadística entre la presencia de síntomas de amarillamiento y los resultados de la prueba TBIA para SCYLV, de acuerdo a las pruebas de Chi-cuadrado (p=0,7204) y Fisher exacta (p=0,4864).
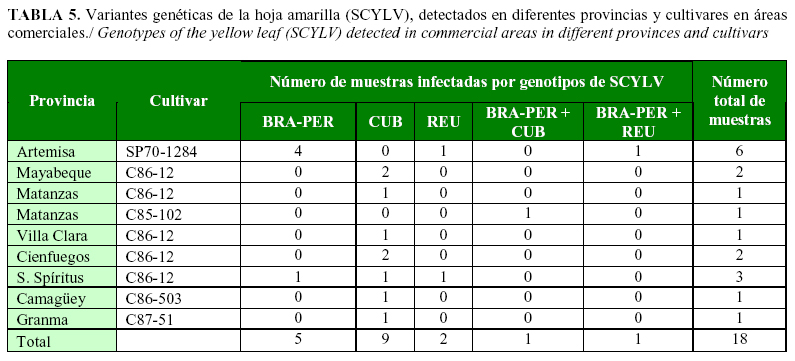
Los cultivares que presentaron mayor grado de severidad de los síntomas, atendiendo a la escala de evaluación y a la frecuencia observada, fueron C132-81, C85-102, C86-12, C87-51, C88-380, C90-469 y SP70-1284. En los cultivares con síntomas se observó el mayor grado de severidad de los síntomas. La presencia de SCYLV fue confirmada, además, por RT-PCR en 18 muestras de los cultivares C85-102, C86-12, C86-503, C87-51 y SP70-1284 (Tabla 4).
Las variantes genéticas BRA-PER, CUB y REU del virus fueron encontrados en 7 (38,8%), 10 (55,5%) y 3 (20%) de las 18 muestras procesadas, respectivamente. En la provincia de Sancti Spíritus se determinaron las tres variantes. Se encontraron infecciones mixtas de BRA-PER + CUB y BRA-PER + REU en las provincias de Matanzas y Artemisa, respectivamente (Tabla 5).
DISCUSIÓN
Se determinó, por primera vez, la presencia del virus de la hoja amarilla (SCYLV) en todas las provincias que cultivan la caña de azúcar en Cuba. La alta incidencia de SCYLV en las muestras analizadas en este estudio, se puede atribuir a la existencia de cultivares comerciales susceptibles, proximidad de fuentes de inóculo, dinámica del vector (Melanaphis sacchari Zehntner) y la propagación a través de material vegetal infectado. En la mayoría de los cultivares comerciales evaluados, se observó el mayor grado de severidad de los síntomas.
Desde la pasada década, se publicaron en Cuba varios artículos sobre la alta incidencia de síntomas asociados a SCYLV y SCYP, lo que indica la posible existencia de fuentes de inóculo importantes de uno o de ambos patógenos.
En el 2005, se informó una incidencia de 16,8 a 39% de infección por SCYP en nueve provincias de Cuba (8). Posteriormente, en Villa Clara se determinó una incidencia de los síntomas en el 47,09 y 32,6% de los campos evaluados en los años 2005 y 2006, respectivamente; mientras que en Cienfuegos, en el año 2005 fue del 38,68%. Se registró la presencia de estos síntomas en el 73,66 y 83,07% del área evaluada en Villa Clara en los años 2005 y 2006, respectivamente; y en el 91,45% del área evaluada en Cienfuegos en el año 2005. La provincia de Cienfuegos presentó mayor incidencia y severidad de los síntomas, en un amplio rango de cultivares, principalmente en C1051-73, C87-51, C120-78, SP70-1284, C86-12, Co997 y C132-8 (24).
La presencia de muestras con síntomas asociados a SCYLV y negativas por TBIA, podría estar relacionada con la baja concentración del virus en muestras con síntomas y falta de sensibilidad para su detección por esta técnica de diagnóstico. También puede ser atribuido a otras causas, entre ellas a la presencia del fitoplasma que causa el «amarillamiento de la hoja» (SCYP), aspecto informado anteriormente (8).
Los resultados sugieren además, que es preciso el diagnóstico de laboratorio para confirmar la presencia o no del virus y que la escala de evaluación visual solo es aplicable para estimar la severidad de los síntomas, que pueden estar asociados tanto al virus SCYLV, como al fitoplasma SCYP.
Las técnicas de diagnóstico moleculares tienen mayor sensibilidad que los métodos inmunoquímicos, pero estos últimos son más económicos, debido a la posibilidad de procesar un número mayor de muestras diariamente, con sensibilidad y especificidad aceptable (21). La combinación de diferentes métodos de laboratorio, como los empleados en este estudio, otros como el diagnóstico molecular fluorescente en tiempo real [Real-time fluorescent (TaqMan) RT-PCR assay] y qRT-PCR para determinación y cuantificación del virus (quantitative real-time RT-PCR assay (qRT-PCR) (25, 26), es la mejor manera de conseguir el diagnóstico exitoso de enfermedades en los campos comerciales para desarrollar las estrategias de manejo adecuadas.
Estos resultados revelan la distribución en el país de tres variantes genéticas del virus: BRA-PER, CUB y REU; esta última se detecta por primera vez en Cuba, por lo que constituye un nuevo informe sobre la distribución mundial de esta variante.
La distribución en el mundo de BRA-PER sugiere que la hoja amarilla fue causada originalmente por esta variante genética, la que se propagó por todo el mundo a través de material de plantas infectadas, incluyendo varias introducciones del virus, cuando el agente causal aún no había sido identificado y detectado por cuarentena vegetal (12). La presencia en Cuba del virus y de las tres variantes, puede deberse a estas causas. En el país, los métodos de diagnóstico no estuvieron disponibles hasta el año 2009, lo que indica la posible incorporación de material genético infectado de otros países a la colección de germoplasma. A su vez, la propagación del virus y sus variantes, año tras año, en áreas de semilla y plantaciones comerciales, al menos desde 1999 cuando se informó de su presencia por primera vez (19), fue favorecida por la presencia del vector M. sacchari y la inexistencia de métodos de control eficientes para la erradicación del virus a través del sistema de producción de semilla certificada.
Anteriormente, fue informada la presencia de dos variantes genéticas del virus, nombrados BRA-PER y CUB, en muestras de los cultivares C132-81 y Ja64-11 procedentes de Cuba, enviadas a Montpellier, Francia, durante una pesquisa internacional (12). BRA-PER es la de mayor distribución en el mundo (1, 12, 27) y CUB fue encontrado además en Brasil, Colombia y Guadalupe; mientras que hasta la fecha, REU solo fue detectada en Brasil, Guadalupe, Mauricio, Barbados y en Isla Reunión (1, 12). La variante genética CUB posee alta incidencia en los cultivares localmente obtenidos en Colombia (12). En Guadalupe, REU es el más común en cultivares locales obtenidos por el programa de mejora de ese país (28). La distribución geográfica limitada de CUB y REU es atribuida a la interacción patógeno - hospedante - vector - ambiente, y por consiguiente, las diferentes evoluciones históricas del virus (12). Estudios posteriores deberán realizarse para determinar la existencia o no de otras variantes genéticas informadas en otras localidades del mundo y el impacto de estas en la producción de caña de azúcar en Cuba.
Esta enfermedad puede causar afectaciones importantes y los daños dependen del cultivar y de la época. En Hawai se determinó que SCYLV reduce en 30% el número de tallos, la biomasa en el 29% y en 26% las toneladas de azúcar por hectárea en el momento de la cosecha, después de los 11 meses (29).
REFERENCIAS
1. Rott P, Mirkov TE, Schenck S, Girard JC. Recent advances in research on Sugarcane yellow leaf virus, the causal agent of sugarcane yellow leaf. Sugarcane International. 2008;26(3):18-22.
2. Girard JC, Fernandez E, Daugrois JH, Roques D, Roumagnac P, Rott P. Genetic diversity of Sugarcane yellow leaf virus in a sugarcane selection plot in Guadeloupe (FWI). International Sugar Journal. 2012;114(358):96-100.
3. You-Xiong Q, Jian-Wei L, Xian-Xian S, Li-Ping X, Ru-Kai Ch. Differential Gene Expression in Sugarcane in Response to Challenge by Fungal Pathogen Ustilago scitaminea Revealed by cDNA-AFLP. Journal of Biomedicine and Biotechnology. 2011.
4. Grisham MP, Eggleston G, Hoy JW, Viatorl RP. The effect of Sugarcane yellow leaf virus infection on yield of sugarcane in Louisiana. Sugar Cane International. 2009;27(3):91-94.
5. Lockhart BE, Cronje CPR. Yellow leaf syndrome. In: Rott P, Bailey R, Comstock JC, Croft B and Saumtally S. (Eds). A guide to sugarcane diseases. CIRAD/ISSCT, La Librairie du Cirad, Montpellier, France. 2000: p 291-295.
6. Izaguirre-Mayoral ML, Carballo O, Alceste C, Romano M, Nass HA. Physiological performance of asymptomatic and yellow leaf syndrome-affected sugarcanes in Venezuela. Journal of Phytopathology. 2002;150:13-19.
7. Smith GR, Braithwaite KS, Cronjé CPR. The viral and phytoplasma forms of yellow leaf syndrome of sugarcane. Proceedings International Society Sugar Cane Technologists. 2001;24:614-617.
8. Arocha Y, Almeida R, Peralta EL, Carvajal O, Jones P. Update data on distribution of yellow leaf síndrome (YLS) in Cuba. Rev Protección Veg. 2005;20(2):1-5.
9. Aday O, Mesa JM, La O-Hechavarría M, Díaz FR, Delgado I, Morales M. Distribución temporal de síntomas del amarillamiento de la hoja en la caña de azúcar en Cuba. Agronomía Mesoamericana. 2012;23(1):1-10.
10.Schenck S, Lehrer AT. Factors affecting the transmission and spread of Sugarcane yellow leaf virus. Plant Disease. 2000;84:1085-1088.
11.Rassaby L, Girard JC, Lemaire O, Costet L, Irey MS, Kodja H, et al. Spread of Sugarcane yellow leaf virus in sugarcane plants and fields on the island of Réunion. Plant Pathology. 2004;53:117-125.
12.Abu Ahmad Y, Royer M, Daugrois JH, Costet L, Lett JM, Victoria JI, et al. Geographical distribution of four Sugarcane yellow leaf virus genotypes. Plant Disease. 2006;90:1156-1160.
13.Viswanathan R, Balamuralikrishnan M, Karuppaiah R. Identification of three genotypes of Sugarcane yellow leaf virus causing yellow leaf disease from India and their molecular characterization. Virus Genes. 2008;37(3):368-379.
14.ElSayed A, Weig A, Komor E. Molecular characterization of Hawaiian Sugarcane yellow leaf virus genotypes and their phylogenetic relationship to strains from other sugarcane-growing countries. European Journal Plant Pathology. 2011;129(3):399-412.
15.Wang MQ, Zhou GH. A near complete genome sequence of a distinct isolate of Sugarcane yellow leaf virus from China, representing a sixth new genotype. Virus Genes. 2010;41:268-272.
16.Wang MQ, Xu DL, Li R, Zhou GH. Genotype identification and genetic diversity of Sugarcane yellow leaf virus in China. Plant Pathology. 2012;61(5):986-993.
17.Lin YH, Gao SJ, Damaj MB, Fu HY, Chen RK, Mirkov TE. Genome characterization of sugarcane yellow leaf virus from China reveals a novel recombinant genotype. Archives of Virology. 2014; SpringerLink, http://link.springer.com/journal705, Published online, 07 January 2014: 1 p.
18.Komor E, EL Sayed A, Lehrer T. Sugarcane yellow leaf virus introduction and spread in Hawaiian sugarcane industry: Retrospective epidemiological study of an unnoticed, mostly asymptomatic plant disease. European Journal Plant Pathology. 2010;127:207-217.
19.Arocha Y, González L, Peralta EL, Jones P. First report of virus and phytoplasma pathogens associated with Yelow Leaf Syndrome of sugarcane in Cuba. Plant Disease. 1999;83:1171.
20.Aday O, Chinea A, Mesa JM, La O M, Zardón M, Díaz FR, et al. Fitoplasmas y virus de la hoja amarilla en el germoplasma y colecciones de caña de azúcar en la región central de Cuba. Fitosanidad. 2011;15(4):195-204.
21.Zardón MA, Gallo A, Mesa JM, Arencibia A, Zamora L, Martínez Y, et al. Detección de infecciones mixtas en genotipos de caña de azúcar en Cuba. Rev Protección Veg. 2012;27(2):77-84.
22.Chinea MA, Pérez G, Aday O, Cabrera L, Pérez JR, Carvajal O. Comportamiento del germoplasma de la caña de azúcar ante el síndrome de la hoja amarilla en Cuba. Memorias Congreso Científico del Instituto Nacional de Ciencias Agrícolas. Mayabeque, Cuba. Noviembre 24 al 28 del 2008: 10p.
23.Schenck S, Hu JS, Lockhart BEL. Use of a tissue blot immunoassay to determine the distribution of Sugarcane yellow leaf virus in Hawaii. Sugar Cane. 1997;4:5-8.
24.Aday O, Chinea A, Díaz FR, Gil Y, Carmenate L, Morales M. Incidencia del síndrome de la hoja amarilla de la caña de azúcar (YLS) en plantaciones comerciales de Villa Clara y Cienfuegos. Centro Agricola. 2012;39(1):17-24.
25.Yun J, Steven TS, Schenck S, Arcinas A, Komor E. RT-PCR and quantitative real-time RT-PCR detection of Sugarcane Yellow Leaf Virus (SCYLV) in symptomatic and asymptomatic plants of Hawaiian sugarcane cultivars and the correlation of SCYLV titre to yield. European Journal of Plant Pathology. 2010;127(2):263-273.
26.Chinnaraja C, Viswanathan R, Sathyabhama M, Parameswari B, Bagyalakshmi K, et al. Quantification of Sugarcane yellow leaf virus in in vitro plantlets and asymptomatic plants of sugarcane by RT-qPCR. Current Science. 2014;106:1-6.
27.Joomun N, Parmessur Y, Antoine M, Dookun-Saumtally A. Mauritius Sugarcane Industry Research Institute, Screening for Sugarcane yellow leaf virus in quarantine in Mauritius. 10th ISSCT Plant Pathology Workshop, Nanning, China, 7-11 May, 2012: 27p.
28.Abu Ahmad Y, Rassaby L, Royer M, Borg Z, Braithwaite KS, Mirkov TE, et al. Yellow leaf of sugarcane is caused by at least three different genotypes of Sugarcane yellow leaf virus, one of which predominates on the Island of Réunion. Archives of Virology. 2006;151:1355-1371.
29.Lehrer A, Wu KK, Komor E. Impact of Sugarcane yellow leaf virus on growth and sugar yield of sugarcane. Journal of General Plant Pathology. 2009;75:288-296.
Recibido: 21-11-2013.
Aceptado: 12-9-2014.

{kind=link}
{kind=link}
{kind=link}
{kind=link}