










Meu SciELO
 Serviços customizados
Serviços customizadosServiços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Protección Vegetal
versão impressa ISSN 1010-2752
Rev. Protección Veg. vol.29 no.3 La Habana set.-dez. 2014
COMUNICACIÓN CORTA
Detección del virus de la poliedrosis nuclear (BmNPV) en el gusano de seda (Bombyx mori L.), mediante hibridación de ácidos nucleicos
Detection of nuclear polyhedrosis virus (BmNPV) in silkworm (Bombyx mori L.) by nucleic acid hybridization
Lidia Chang-SidorchukI, Heidy González-AlvarezI, Bassanio A. GraneauII, Yamila Martínez-ZubiaurI
ILaboratorio de Virología Vegetal y Molecular. Dpto de Fitopatología. Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de Las Lajas, Mayabeque, Cuba. Correo electrónico: lchang@censa.edu.cu.
IIFacultad de Biología, Universidad de La Habana, Cuba.
RESUMEN
El gusano de seda de la morera, Bombyx mori (Lepidoptera: Bombycidae), es afectado por numerosas enfermedades como la Grasserie, provocada por el virus de la poliedrosis nuclear (BmNPV). Su manejo y control se dirigen principalmente a la prevención y diagnóstico temprano. El objetivo de este trabajo estuvo encaminado al diagnóstico del virus BmNPV asociado a los síntomas de Grasserie, observados en las crías de gusano de seda en Cuba. Se desarrollaron los métodos de diagnóstico molecular basados en la hibridación radiactiva y no radiactiva de ácidos nucleicos, a partir de extracciones de poliedros de larvas del tercer y cuarto instar. En ambos métodos de hibridación se pudo detectar la presencia de BmNPV. En el caso de la hibridación radiactiva, se obtuvieron resultados positivos del 5,5% de las muestras analizadas del tercer instar, lo que coincide con los resultados de la hibridación no radiactiva. Sin embargo, en el cuarto instar se detectó el 28,8% de muestras positivas por hibridación radiactiva y 22,2% por la no radiactiva, del total de muestras analizadas. Aunque la hibridación radiactiva mostró más sensibilidad, ambos métodos permitieron confirmar que es posible detectar BmNPV a partir del tercer instar, lo cual permite establecer las medidas de tratamiento preventivo y manejo de las crías para minimizar las pérdidas productivas.
Palabras clave: Bombyx mori, hibridación de ácidos nucleicos, virus de la poliedrosis nuclear, sericultura.
ABSTRACT
The mulberry silkworm, Bombyx mori L. (Lepidoptera: Bombycidae) is affected by numerous diseases like Grassery, caused by the nuclear polyhedrosis virus (BmNPV). The management and control of these diseases mainly consist of prevention and early diagnosis. The objective of this work was aimed at the diagnosis of the virus BmNPV associated with the symptoms of Grassery observed in the production areas of the Mulberry silkworm in Cuba. Molecular diagnostic methods for BmNPV based on radioactive and non-radioactive hybridization of nucleic acids (HNA), from larvae's polyhedra extractions in the 3rd and 4th instars, were developed. In both hybridization methods, BmNPV presence was detected. Positive results of 5.5% of samples analyzed in the 3rd larval stage were obtained using radioactive hybridization, which agreed with non-radioactive hybridization results. Nevertheless, in the 4th stage, 28.8% of positive samples were detected using radioactive hybridization and 22.2% of total analyzed samples for non-radioactive hybridization. Although radioactive hybridization showed higher sensibility, both methods allowed to confirm that it was possible to detect BmNPV from the 3rd larval stage, which allows to establish the enhancement of preventive treatment and management measures of mulberry litters with the objective of minimizing production loses.
Key words: Bombyx mori, nucleic acid hybridization, nuclear polyhedrosis virus, sericulture.
El gusano de seda de la morera, Bombyx mori L. (Lepidoptera: Bombycidae), es afectado por un gran número de patógenos como son virus, bacterias, hongos y microsporidios, agentes causales de la Grasserie, Flacherie, Pebrina y Muscardina (1, 2, 3). Estas enfermedades son bien conocidas en casi todas las áreas del mundo donde practican la sericultura, y provocan daños considerables a las cosechas de cocones del gusano de seda (4).
La Grasserie, provocada por un virus del género baculovirus (virus de la poliedrosis nuclear de Bombyx mori (BmNPV)), es una de las enfermedades virales de mayor impacto mundial. Puede llegar a provocar entre el 70-80% de las pérdidas en producciones (5), afecta a todos los países productores, principalmente los tropicales, donde se puede manifestar todo el año (6).
En 2011 se iniciaron en Cuba las producciones masivas de este insecto y se obtuvieron los primeros niveles de producción. Sin embargo, a partir del 5to instar del ciclo de vida del insecto, comenzaron a aparecer sintomatologías que sugirieron la presencia de enfermedades. Los resultados de patogenicidad, microscopía electrónica y detección molecular realizados por Martínez et al. (7), confirmaron la presencia en las crías del virus de la poliedrosis nuclear (BmNPV), como agente patógeno causante de los síntomas observados.
Los trabajos de identificación y caracterización de este virus realizados en Cuba, permitieron obtener herramientas para el diagnóstico molecular de estos patógenos mediante la Reacción en Cadena de la Polimerasa (PCR). Sin embargo, nuevos métodos se desarrollan en la actualidad a fin de contar con medios más simples y a la vez con alta sensibilidad y confiabilidad.
Los ensayos de hibridación de ácidos nucleicos constituyen una herramienta muy útil en el diagnóstico e identificación de los virus de la poliedrosis nuclear (NPV). En los últimos años, varios autores informaron el empleo de diferentes variantes de hibridación para la identificación y cuantificación de NPV de varias especies (1, 8, 9).
El objetivo del presente trabajo fue evaluar los métodos de hibridación molecular, con relevado radiactivo y no radiactivo, para la detección del virus de la poliedrosis nuclear de B. mori en el tercer y cuarto instar del ciclo de vida en condiciones de producción.
En ambos métodos de hibridación se aplicó el mismo procedimiento variando el marcaje de la sonda. Se empleó, como sonda, el fragmento del gen de la poliedrina de baculovirus obtenido por PCR con cebadores polheforward: 5´CGTGTACGACAACAAGTAC3´ y polhereverse: 5´AAAGTGAGTTTTTGGTTTTTGCC3´ descritos por Szewczyk et al. (10).
El fragmento fue purificado con el sistema de purificación Wizard SV Gel and PCR Clean-up System (Promega), según el protocolo descrito (https://www.promega.com). Para la hibridación radiactiva se marcaron 25ng de ADN con a32 P-(dATP) según el protocolo descrito en el juego de reactivos Megaprime (Amersham). La sonda no radioactiva se marcó mediante el juego de reactivos «DIG DNA Labeling kit» (Roche), siguiendo el procedimiento indicado en el instructivo de uso adjunto al juego de reactivos.
Para la extracción de cuerpos de poliedros se tomaron 36 larvas del 3er y 4to instar de diferentes camas y seleccionadas al azar, se conformaron dos grupos de 18 larvas. Se maceraron en tampón de homogenización (1% ácido ascórbico, 2% SDS, 0.01MTris pH 7.8 y 0.001M EDTA) y se filtraron a través de tres capas de gasa estéril. El homogeneizado se centrifugó a 10000 rpm, por 12 min en centrífuga Sigma y se resuspendió el precipitado en tampón TE (50 mM tris: 10 mM EDTA) pH 7,8. Esta suspensión se transfirió a tubos limpios y se centrifugó a 12 000 rpm por 12 minutos a 4°C; se eliminó el sobrenadante y el precipitado se resuspendió en tampón TE 1X.
Los grupos de poliedros de cada edad fueron desnaturalizados antes de aplicarlos a las dos membranas, a partir de la centrifugación de un 1ml de suspensión de poliedros a 12000 rpm por 10 min, el precipitado se homogenizó en 200µl de solución Na2CO3 (0,02mM; pH 11). La mezcla se incubó durante 20 minutos a 25°C, con agitación ligera en el equipo Thermo-Shaker (Fisher). A cada tubo se añadieron 30 µl de solución de acetato de sodio (0, 16 M; pH 5) y se incubaron 10 minutos a 94ºC. Se aplicaron 20 µl de cada muestra en ambas membranas de nylon (Hynbond N+, Amersham). Se utilizó una muestra con BmNPV como control positivo y un control de sonda.
Las soluciones de prehibridación e hibridación se prepararon con la siguiente mezcla: 5X de Denhardt (Sigma Chemical Co.), 5X SSC (150 mM NaCl, 15 mM citrato de sodio, pH 7.0), 50% formamida, 250µg/ml de ADN de esperma de salmón, 50 mM de pirofosfato de sodio pH 6.5, y 0.1% SDS.
Las membranas se prehibridaron durante 2 horas a 42ºC. Luego se añadieron las sondas marcadas y se dejaron toda la noche hibridando a 42ºC. Tras la hibridación se realizaron tres lavados de 15 minutos, con una solución 1X SSC y 0.1% SDS, dos veces a temperatura de 55ºC y una vez a 65ºC.
Las membranas se expusieron durante 16 horas en el casete de exposición (Kodak) a temperatura ambiente. Los revelados se realizaron en cuarto oscuro con soluciones reveladoras y fijadoras, según Sambrook et al. (11).
Mediante ambos métodos de hibridación, se pudo detectar la presencia del BmNPV a partir de las dos etapas analizadas (Figura). En el 4to instar se detectó por hibridación radiactiva un ligero aumento en el número de larvas infectadas. Este resultado coincide con otros autores que plantean menor sensibilidad en la detección de patógenos virales con el uso de la detección no radiactiva, lo que pudiera estar determinado por el tipo de marcaje (12). Sin embargo, en nuestro caso, aunque hubo detección del virus desde el 3er instar, no se mostraron diferencias por ambos métodos de detección en cuanto al número de larvas infectadas. La detección de BmNPV desde edades tempranas de las crianzas es de gran importancia, ya que en estas etapas tempranas del ciclo de vida es imprescindible realizar un diagnóstico con el fin de aplicar los tratamientos de manejo para la prevención y dispersión de las enfermedades.
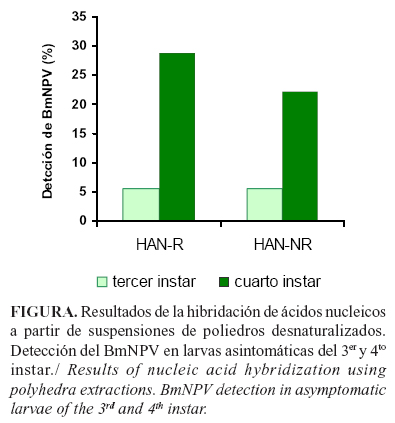
Con ambos métodos se observó un mayor avance de la enfermedad en las larvas a partir del cuarto instar, demostrado por un mayor porcentaje de detección de larvas infectadas, lo que pudiera estar asociado a una producción creciente y exponencial de poliedros en esta etapa que desencadena en la expresión de los síntomas en el quinto instar, tal como ha sido descrito por Cifuentes (6).
Por otra parte, los resultados sugieren que la hibridación de poliedros desnaturalizados puede ser utilizada como método de detección de BmNPV en condiciones de producción, tal como ha ocurrido en la detección de otros virus de lepidópteros (1, 8, 9). Este método permitirá el análisis de gran cantidad de muestras en condiciones extensivas de producción y facilitará el trabajo, pues brinda la posibilidad de una detección a partir de material semipurificado, en este caso poliedros, ya que en la PCR los contaminantes pueden inhibir la acción de la enzima Taq polimerasa, lo que exige necesariamente un ADN purificado.
Los resultados permitieron tomar medidas de tratamiento preventivo y manejo de las crías desde el tercer instar, a fin de minimizar las pérdidas productivas por la incidencia de la enfermedad.
Aunque la hibridación no radiactiva mostró un menor porcentaje de detección, se sugiere la optimización de esta variante de hibridación, con el fin de minimizar los riesgos que conlleva el uso de la radioactividad, tanto para el personal que labora con ella, como para el medio ambiente.
REFERENCIAS
1. Keating ST, Keating JP, Elkinton J. DNA hybridization assay for detection of gypsy moth nuclear polyhedrosis virus in infected gypsy moth (Lymantria dispar L.) larvae. Appl Environ Microbiol. 1989;55(11):2749-2754.
2. Rao S, Nageswara M, Muthulakshmi S, et al. Phylogenetic Relationships of Three New Microsporidian Isolates from the Silkworm, Bombyx mori. J Inver Pathol. 2004;86:87-95.
3. Sakthivel S, Angaleswari C, Mahalingan P. Isolation and Identification of Bacteria Responsible for Flacherie in Silkworms. Advances in Applied Science Research. 2012;3(6):4066-4068.
4. Chakravorty R, Das R, Neog K, Das K, Sahu M. A diagonostic manual for diseases and pest of muga silkworm and their host plants. 2007; 1-47.
5. Babu K, Ramakrishna S, Reddy Y. Metabolic Alterations and Molecular Mechanism in Silkworm Larvae during Viral Infection: A Review. African J Biotech. 2009;8(6):899-907.
6. Cifuentes C, Cesar A, Sohn K. Manual Técnico de Sericultura: Cultivo de la Morera y Cría Del Gusano de Seda en El Trópico. Pereira, Convenio SENA-CDTS. 1998; 438.
7. Martinez Y, Prieto M, Shilher W, Marrero Y, Lobo de Souza M, et al. Detection of Bombyx mori Nucleopolyhedrosis viruses (BmNPV). Resúmenes XVIII Congreso INCA. 2012. http://ediciones.inca.edu.cu/files/congresos/2012 /resumenes_2012.pdf.
8. Eblin P, Smith P, Frankenhuyzen V. DNA Hybridization Assay for Detection of Nucleopolyhedrovirus in Whitemarked Tussock Moth (Orgyia leucostigma) Larvae. Pest Manag. Sci. 2001;57(1):66-71.
9. Thorne C, Imre S, Nicholas C, Levin D. Development and Evaluation of Methods to Detect Nucleopolyhedroviruses in Larvae of the Douglas-fir Tussock Moth Orgyia pseudotsugata (Mc Donnough). Appl Eviron Microbiol. 2007;73(4):1101.
10.Szewczyk B, Piotr B, William S, Lukasz R, Iwona S, Marlinda L. Detection and Identification of Baculovirus Pesticides by Multitemperature Single Strand Conformational Polymorphism. J Environ Sci and Health. 2008;B43(7):539-545.
11.Sambrook J, Fritsch E, Maniatis T.Cold Spring Harbor Laboratory Press. Molecular cloning. 2da. Edición. 2008.
12.Potter J. PCR and DNA Hibridization methods for specific detection and identification of Bean-infecting begomovirus. Tesis de Maestría. Universidad de Wisconsin-Madison. 2001; 221.
Recibido: 6-10-2014.
Aceptado: 5-11-2014.
