










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión On-line ISSN 2224-4697
Rev. Protección Veg. vol.30 no.1 La Habana ene.-abr. 2015
ORIGINAL ARTICLE
Biology and population parameters of Tuta absoluta (Meyrick) under laboratory conditions
Biología y parámetros poblacionales de Tuta absoluta (Meyrick) bajo condiciones de laboratorio
Leticia DuarteI, María de los Ángeles MartínezI, Vanda Helena Paes BuenoII
IDirección de Sanidad Vegetal. Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, Mayabeque, Cuba. Correo electrónico: leticia@censa.edu.cu.
IILaboratorio de Control Biológico. Departamento do Entomología, Universidad Federal de Lavras, Lavras, Minas Gerais, Brasil.
ABSTRACT
The aim of this work was to determine the temperature effect on Tuta absoluta (Meyrick) biological development. Its biological and population parameters were evaluated on the Cuban tomato (Solanum lycopersicum L.) cultivar Vyta at temperatures of 25±1oC and 20/30±1oC under controlled conditions. The duration (in days) of the egg, larval and pupal stages and reproductive periods, as well as the sex ratio, the viability of eggs and pupae (in per cent), total fecundity/female and longevity obtained at different temperature regimens were compared between treatments by ANOVA with Mann-Whitney test (p<0.05). The larval survival (in per cent) was compared between treatments by ANOVA with Bonferroni test (p<0.05).Only the egg stage and the pre-oviposition period were longer with 20/30oC (p <.0001). Viability of the pupae was higher at (25oC) (p=0.0121).The larval survival was affected negatively with 32.14 and 20% at 25oC and 20/30oC, respectively (p=0.0206). The survival curves for larvae and female adults in both conditions were of type II. The survival of females subjected to (20/30oC) was three days longer, but without significant differences between treatments. The total fecundity/female and longevity were not affected. The specific fecundity (mx) in both temperatures was more stable in a range of 0.8-1.0 until the 10 days of age. The population growth parameters of T. absoluta Ro, T, rm, TD and (l) were 9.36, 8.90, 5.52, 5.75, 0.02, 0.01, 34.64, 117.01, and 1.02, 1.01 at 25oC and 20/30oC, respectively. The biological and population parameters of T. absoluta on the tomato Vyta cultivar were influenced by the temperature in the test, and, as a consequence, the development of the pest was limited under the laboratory conditions. In addition, indications of Vyta as a possible resistant cultivar versus the pest were shown under the temperature regimen studied.
Key words: leaf miner, host plant resistant, Solanum lycopersicum.
RESUMEN
El objetivo de este trabajo fue determinar el efecto de la temperatura sobre el desarrollo biológico de Tuta absoluta (Meyrick). Los parámetros biológicos y poblacionales fueron evaluados a temperaturas de 25±1oC y 20/30±1oC sobre el cultivar de tomate (Solanum lycopersicum L.) Vyta en condiciones controladas. La duración (en días) de las fases de huevo, larva, pupa y períodos reproductivos, así como razón sexual, viabilidad de los huevos y pupas (en por ciento), fecundidad total/hembra y la longevidad obtenida en los diferentes regímenes de temperatura por ANOVA con prueba de Mann-Whitney (p<0.05), se compararon entre tratamientos. La supervivencia larval (en por ciento) se comparó entre los tratamientos por ANOVA, con una prueba de Bonferroni (p<0.05). Solo la fase de huevo y el periodo de pre-oviposición fueron más largos con 20/30oC (p <.0001). La viabilidad pupal fue superior a 25oC (p=0,0121). La supervivencia larval se afectó negativamente con 32,14 y 20% a 25oC y 20/30oC, respectivamente (p=0,0206). En ambas condiciones, las curvas de supervivencia para larvas y hembras adultas fueron de tipo II. La supervivencia de las hembras sometidas a 20/30oC fue tres días más larga, pero sin diferencias significativas entre los tratamientos. La fecundidad total/hembra y la longevidad no se afectaron. La fecundidad específica (mx) fue más estable, en un rango de 0.8-1.0 hasta los 10 días de edad en ambas temperaturas. Los parámetros de crecimiento poblacional de T. absoluta Ro, T, rm, TD y (l) fueron 9,36, 8,90; 5,52, 5,75; 0,02, 0,01; 34,64; 117,01 y 1,02, 1,01 a (25oC) y (20/30oC), respectivamente. El desarrollo biológico y parámetros poblacionales de T. absoluta sobre el cultivar Vyta, estuvieron influenciados en el estudio por la temperatura, y como consecuencia, el desarrollo de la plaga se limitó en condiciones de laboratorio. Además, se mostraron indicios de Vyta como un posible cultivar resistente, frente a la plaga bajo el régimen de temperatura estudiado.
Palabras clave: minador de hojas, resistencia de planta hospedante, Solanum lycopersicum.
INTRODUCTION
Tomato (Solanum lycopersicum L.) is original from Andean countries and is one of the vegetables most cultivated in the world (1). The present world production is about 152.1 million tons of fresh fruit (2). Despite of this, the crop is attacked from planting to harvest by numerous pests and diseases (3).
The tomato leaf miner Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae), is an extremely devastating pest that may decrease fruit quality, or even, cause 100% losses in open field and greenhouse crops, mainly if control methods are not applied (4). It was first described in Peru in 1917 and is considered one of the most devastating pests for the tomato crop in South America and a serious threat to tomato production in the Mediterranean region (5). Invasion of the Mediterranean Basin was rapid; T. absoluta traveled about 4,000 km in five years and has become now a major threat to tomato production in three continents (Europe, Africa, and Asia) (6). It extends North-South from The Netherlands to Sudan, and West-East from Portugal to Iran (7). The presence of this pest was also reported in Panama (8).
This species is a multivoltine pest that mines leaves, fruits, flowers, buds and stems. The damage is produced when the larvae feed on the leaf mesophyll expanding mines, thus affecting the photosynthetic capacity of the crop with the subsequent yield reduction. T. absoluta is an oligophagous insect that feeds on solanaceous species, where, besides tomato, is associated with other cultivated and non-cultivated host-plants (9). The duration of the life cycle depends on the environmental conditions, in particular the temperature, ranging from 76.3 days at 14°C to 23.8 days at 27°C (10). Also there may be an overlap in generations and all the stages of the cycle are found in field conditions (11). Temperature variations directly influence the development rate of insects and indirectly influence their development through effects on the host plant, and so, the plant quality as food resource (12).
It is known that temperature has an important influence on duration of the different development stages of the insects allowing a higher or lower exposure time of them to the natural enemies (13). Additionally, negative effects of ecological factors on the multiplication of the population could be considered, and consequently, a higher reduction of the number of progenies in natural conditions. van Lenteren (14) reports that the research involving knowledge of plant resistance to pests is a challenge that should be displayed for the sustainable growth of plant production systems. Understanding the relationship between temperature variation and tomato cultivars concerning pest development is important to evaluate the control option of host-plant resistance and to determine whether a combination of host-plant resistance with biological control is a realistic option.
The tomato leaf miner is included in the Official list of quarantine of the Cuban Republic; its introduction constitutes a risk for the country, and the resistance level of the Cuban cultivars versus this species has not been studied. The phytogenetic resources are known to contain an important quantity of genes for being managed and explored in a rational form,mainly those genera of plants related to resistance to biotic and abiotic factors that affect cultivated species by the man (15).
In Cuba, the introgressions of the Ty-1 gene from native species of tomato (Solanum chilense) to cultivated plants (S. esculentum) allowed to obtain the resistance lines in the whitefly-geminivirus complex, until the generalization of the Vyta commercial cultivar for the best productive areas in the country (16). In this paper, it is reported a study on the development and populational growth of T. absoluta on the Cuban tomato cultivar Vyta under constant and alternating temperature regimes.
MATERIALS AND METHODS
The experiments were carried out at the Laboratory of Biological Control, Department of Entomology, Universidade Federal de Lavras (UFLA), Lavras, Minas Gerais, Brazil.
The tomato seeds of the Cuban cultivar Vyta were supplied by the Horticultural Research Institute «Liliana Dimitrova», located in Quivicán, Mayabeque, Cuba. The seedlings were produced in root ball trays with substrate UFLA in a greenhouse. After 20 days, they were transferred to 5L pots. The pots were provided with a mixture of soil, sand, calcium, a complete formula of nitrogen, phosphorum and potassium (NPK) and substrate UFLA for obtaining a higher nutritional value and a consequent best pest population development. Each pot was supplied with 0.83, 0.42, 0.02, 0.13 and 0.42Kg of each component, respectively. The plants were fertilized weekly with 100 ml per pot of a dissolution of KNO3. The plants were subjected to 23±7,6oC of temperature, 64±15.4% of relative humidity and 12 hours of photoperiod. The plants were irrigated by aspersion with a frequency of one hour by space of five minutes.
The regime of constant and alternating temperatures used in the experiment was defined according to studies on T. absoluta development conducted at different temperatures reported in the literature (9,17). Moreover, this study considered the temperature average from regions of the country where the largest Cuban tomato productions have been achieved. The constant and alternating temperature at 25°C and 20/30°C, were chosen, respectively. According to the temperature fluctuations, the values of alternating temperature were considered minimum and maximum according to night or daytime. The experiments were carried out in climatic chambers. Two temperature treatments were used: 25°C and 20/30°C, with a relative humidity of 70±10% and photoperiod of 12 hours.
Rearing of Tuta absoluta on plant of the tomato cultivar Vyta under laboratory conditions.
Adults of T. absoluta of 48-72 hours of emerged were collected from the pest rearing established in Santa Clara at the Biological Control Laboratory in UFLA. Plants of the Vyta cultivar were placed into acrylic cages (60x30x30 cm) with 100 adults of the pest. After 48 hours, the rearing was started with the eggs oviposited by the adults. The laboratory conditions used for the rearing were 25±2°C of temperature, 70%±10% of relative humidity and 12 hours of photoperiod. Eggs, larvae and adults of T. absoluta from individuals of the second generation were used for starting the tests.
Development and survival of T. absoluta
Egg stage
To determine the duration of the egg stage and their viability, T. absoluta couples were placed into a transparent acrylic glasses (6.5 cm, 8.0 cm diameter) for 24 hours. Each glass was fixed to a Petri dish (10 cm diameter) with an adhesive film. Each dish contained a leaf of the Vyta cultivar inserted into an Eppendorf tube with distilled water and supported to a polystyrene cube. Tomato leaves were used as oviposition substrate. The adults were fed with drops of honey. A few portion of this was placed inside of the glasses with the help of an entomological brush. After 24 hours, the excess of eggs present on the leaves were removed with a fine-tipped brush. Only, fifteen eggs were left per leaf. Ten replicates per each treatment were included (n= 150) The larvae hatched were counted daily (Figure 1).

Larval stage
Newly hatched larvae from the pest reared at the laboratory were used in the test. Stems of tomato leaflets were wrapped with cotton moistened with distilled water and placed into the Petri dishes. Five larvae of T. absoluta were placed on each leaf using a fine brush under a stereoscopic microscope (Zeiss Stemi 2000-C), with a magnification of 1x10. Twenty-eight Petri dishes (Ø 8 cm) containing a leaf with five larvae per each was considered, for a total of 140 larvae per treatment. The leaves were changed every two days to allow the development of the larvae until pupae. A disk of filter paper was placed in the bottom of each Petri dish for absorbing the excess moisture. Then, the dishes were sealed with PVC film. The number of live larvae was counted daily.
Pupal stage
When the larvae reached the pupal stage, they were individually placed into a glass tube (8.5 x 2.5). The pupae were observed daily until adult emergence. Considering larvae survival until reaching the pupal stage, the number of pupae evaluated was 45 and 38 at 25 and 20/30oC, respectively. The emerged adults were sexed according to the methodology proposed by Coelho and Franca (18), and the presence of possible deformations was observed. The sex ratio was evaluated in both treatments.
Effects of the Vyta tomato cultivar and temperature on the reproduction and population parameters of T. absoluta.
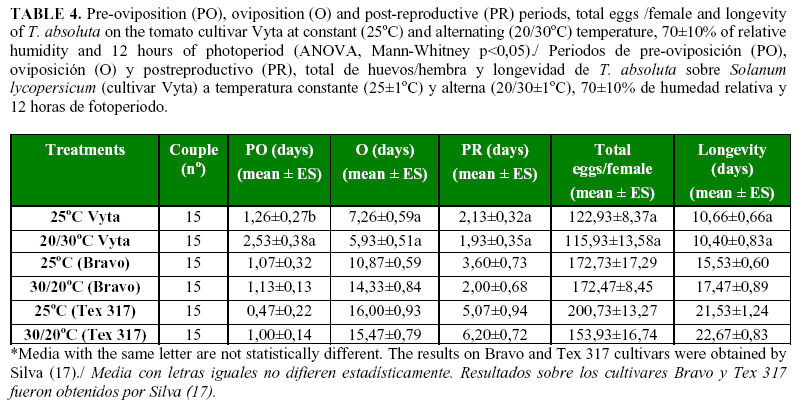
To determine the reproductive potential and survival of adults, the newly emerged couples of T. absoluta at constant and alternating temperature from the pupae stage test were placed into a transparent acrylic glass (6.5 cm, 8.0 cm diameter) until death. The glass was supported with a Petri dish (9cm diameter) with the same conditions above mentioned. The leaves of the tomato cultivar were used as substrate of oviposition for adults of T. absoluta (Figure 1).
Fifteen couples (1 couple per glass) per each treatment were placed on the leaves in the experimental arena. A few drops of honey were placed for feeding the insects. The small leaves were removed daily and the number of eggs per couple counted and death of the adults recorded. In addition, the pre-oviposition, oviposition and post-reproductive periods, as well as the longevity of the adults were evaluated according to the methodology described by Borgoni & Carvalho (19) and Silva (17).
Fertility life table
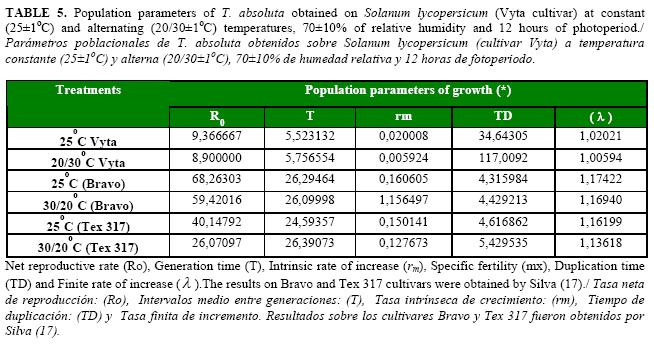
To estimate the population parameters of T. absoluta on the Vyta tomato cultivar under both temperature conditions, the fertility life table was constructed per each treatment. The rate of survival (lx), specific fertility (mx), net reproductive rate (Ro), intrinsic rate of increase (rm), generation time (T), finite rate of increase () and duplication time (TD) were determined in the test. Some definitions (Table 1) and calculations of these parameters were as follows:

Data analysis
A Kolmogorov Smirnov test was used to know the normal distribution of the data in the experiment. The duration (in days) of the egg, larval and pupal stages and the reproductive periods, as well as the sex ratio, the viability of eggs and pupae (in per cent), total fecundity/female and longevity obtained at different temperature regimens were subjected to an ANOVA with the Mann-Whitney test (p<0.05) for the comparison between treatments. The larval survival (in per cent) subjected to an ANOVA with the Bonferroni test (p<0.05) for the comparison between treatments. The SAS program, version 9.0 was used for the statistical analyses.
RESULTS
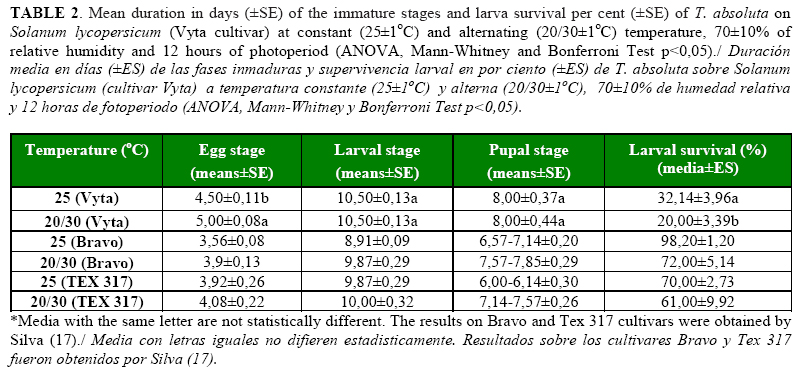
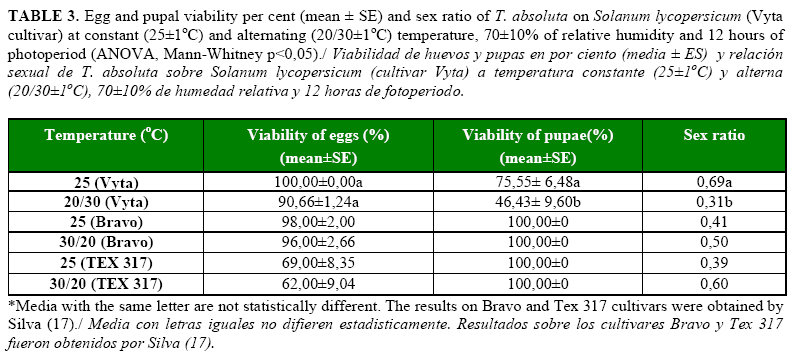
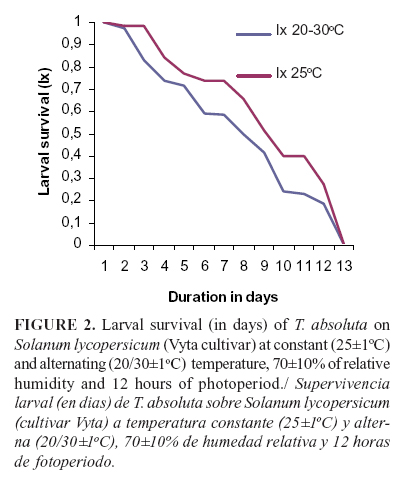
The duration of the larval and pupal stages of T. absoluta did not show significant differences under both temperature conditions. However, the duration of the egg stage of the pest was significantly longer (p <.0001) under the alternating (20/30oC) than under the constant temperature (25oC). Despite of this, the immature stages of the insect on the Vyta cultivar were longer than on the tomato cultivars evaluated by Silva (17) under the same temperature regimens (Table 2). The highest effect of both temperatures on T. absoluta development was showed by the viability of the pupae, with significant differences between treatments (p=0.0121) (Table 3), and larvae survival was affected negatively. The average percentage of survival showed statistical differences (p=0.0206) between treatments and the highest effects were obtained under the alternating temperature (Table 3). The larval survival described a curve of type II under both treatments, where a constant number of individuals died by time unit (Figure 2). The sex ratio of the adults were affected by the temperature with statistical differences between treatments (p=0.0035); the highest effect was obtained when the Vyta cultivar was combined with the alternating temperature (Table 3).
The duration of the reproductive periods of the adults, the fecundity of the females and longevity were not affected notably by the temperature conditions in the test; only the pre-oviposition period was longer with alternating temperature than with the constant temperature (p <.0001) (Table 4). However, the duration of these periods, total eggs/female and longevity of T. absoluta were lower than what was obtained by the pest on the Bravo and Tex 317 cultivars at the same temperature conditions (17). The low oviposition period limited the pest oviposition time, and as consecuence, an lower quantity of eggs was laid by T. absoluta females on the Vyta cultivar.
Moreover, the survival of the female adults showed a curve of the type II, where a constant number of individuals died by time unit. The females subjected to alternating (20/30oC) temperature reached three more days of life than the females under constant temperature (25oC), for a survival of 16 days. The highest population mortality started in a range of 5-7 days of age, in both conditions. In addition, the specific fecundity (mx) was better established in a range of values of 0.8-1.0 until the 10 days of age of the population despite the progressive death of individuals by time unit (Figure 3).
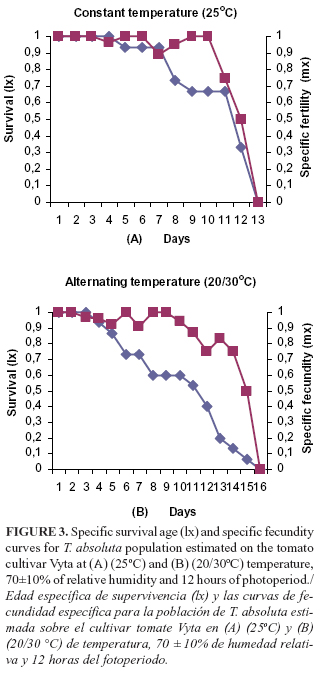
The lowest value of the finite rate of increase (l) of the pest was obtained with the alternating (20/30oC) temperature as a consequence of the increase of the time necessary for the duplication of individuals in the population (TD). The values of the intrinsic rate of increase (rm), were lower than the values of the (l), but, in this case, the higher value of rm was reached by the pest with the constant (25oC) temperature. Under both temperature conditions, these values were positive indicating the populational growth of T. absoluta under laboratory conditions (Table 5).
The net reproductive rate (Ro) of T. absoluta was higher at 25oC than at 20/30oC, allowing a better population increase (rm) of the pest under constant temperature. The generation time (T) was higher under the alternating temperature in the laboratory conditions (Table 5), indicating a longer exposure of the population to natural enemies or ecological factors by a higher interval between generations; this results indicated that Ro and rm of the pest could be affected in natural conditions.
DISCUSSION
The results of the biological and population parameters of T. absoluta on the tomato cultivar Vyta in the present study differed from those obtained by Silva (17) on Brazilian tomato cultivars under similar temperature conditions. In comparing these results, it was allowed making evident the potential of the Cuban cultivar since the resistance of the commercially extended Brazilian cultivars to T. absoluta is quite known (20). It was the first time that the Cuban cultivar was tested against this pest based on its resistance to whitefly-begomovirus complex (21).
In this work, the results indicated that the combination of temperatures with the Vyta cultivar could be an important tool to combat the pest. Besides, the longer duration of the egg, larval and pupal stages obtained on Vyta cultivar could allow a higher exposure time of the pest to the natural enemies in natural conditions. Moreover, it is known the potential use of predators, parasitoids and entomopathogens as biological control agents of T. absoluta (22). Some of these control agents, mainly indigenous mirid predators, have already been successfully used in integrated pest management programs (23).
The response of T. abosoluta to the alternating temperature could be attributed to changes in the insect causing disorders on its metabolism and development of immature phases. Due to being poikilotermic organims, insects do not present mechanisms to regulate their body temperature and, therefore, their physiology is strongly influenced by this factor (24). Thus, it is possible that the alterations produced within a phase may influence the development in the next phase.
Regarding insect sex ratio, is a parameter that can be influenced by several factors. It is documented that the patterns defining sex are the population mating structure and the environmental conditions (25). About this latter, it is known that temperature can aversely influence on sperm viability (26), an element that could explain why the sex ratio was in favor of males at the alternating temperature in the present study.
The sex rate values of T. absoluta observed in the present work were contrary to those obtained by Silva (17), so that the numbers of males or females in the population could be attributed to the combined effect of the temperature with the tomato variety on which the insects were developed.
These results suggest that T. absoluta has a limited development of the Vyta variety. The low larval survival reached by the insect under both temperature conditions are considered a result of great importance, taking into account that the crop damages are caused by the larvae.
When the results on larval survival and pupa viability herein obtained were compared with those obtained by Pereyra and Sánchez (9) and Moreira et al. (27) on other tomato varieties under similar temperature conditions, the highest values of these parameters obtained by these authors suggest a negative effect of the Vyta variety on T. absoluta development.
On the other hand, although no significant differences among treatments were found in the insect oviposition period, this period was shorter than that obtained by Moreira et al. (27), who determined the resistance of the tomato wild lines S. hirsutum f. glabratum (PI134417), S. pimpinellifolium (PI 126931), and S. peruvianum at 25 oC , and they concluded that the short oviposition period could be related to the presence of toxic substances in the leaves which reduced oviposition quickly after the insect got in touch with them.
Likewise, although no significant differences among treatment were observed in the species fecundity, the values obtained in the present study were far below those reached by Thomazini et al., (28) y Silva (17).
In this respect, Ecole et al. (29) and Gonçalves-Neto et al. (30) state that the resistance of tomato varieties to T. absoluta is attributed to the presence of allelochemicals in the glandular thricomes of the leaves and the methyl-ketona 2 tridecanona is the major secondary metabolite responsible for resistance.
The glandular thricomes of Solanum species release toxic exudates or resins when they are touched by phytophagous organisms. These irritating substances can potentially cause death. Research on the roll of thricomes in the resistance of the tomato crop as a host has been mainly focused on S. hirsutum, S. hirsutum f. glabratum and S. pennellii, which are resistant to Keiferia lycopersicella (Walsingham) (Lepidoptera: Gelechiidae) because they get fewer insect larvae and the s on the leaves are smaller when they are compared with S. lycopersicum (31).
Different hybrid combinations between genotypes with high and low levels of aminoacids showed excelent levels of resistance to T. absoluta. These allelochemicals can act preventing the oviposition, feeding, and producing a lethal effect on the development of some insects phases (20).
In turn, Leite (32) defines that tomato varieties show resistance by antibiosis when a longer duration of the insect development phases and a low pupa viability, larval survival and female fecundity are observed. These criteria agree with the results attained in the present study, suggesting that the Vyta variety can exert antibiosis as a resistance mechanism to T. absoluta.
In this sense, Ecole et al. (29) consider that a longer duration and a lower larval survival of T. absoluta on tomato wild accessions of S. hirsutum f. typicum reinforce the hypothesis that antibiosis and antixenosis are the mechanisms of these genotypes involved in the resistance to tomato moth, since they cause a higher larval mortality by inanition or antibiotic action.
This type of mechanims were referred to by Saldúa and Castro (33) when they evaluated the resistance of different comercial wheat varieties against Sipha maydis Passerini (Hemiptera: Aphididae), and they concluded that the antixenosis exerts a strong selection pressure on the insect population due to volatile compounds inducing the not preference, whereas the antibiosis, based on anti-nutitive principles afects the multiple metabolic proccesses of the aphid. These criterion could be associated with the limited development of the immature phase of T. absoluta affecting, consequently, the insect population increase in the present study.
On the other hand, Gilardón et al. (34) evaluated the resistance of 24 tomato lines from crosses between S. lycopersicum y S. hirsutum f. glabratum against T. absoluta, and concluded that those lines manifesting resistance exerted antibiosis as the resistance mechanism to the first insect larval stages, which was correlated with a low average weight of fruits, a result that corroborated the difficulties in selecting lines with a high resistance to tomato moth and big fruits.
CONCLUSION
The biological and population parameters of T. absoluta maintained on the Vyta tomato cultivar were influenced by temperature, and development of the pest was consequently limited. The individuals subjected to alternating (20/30oC) temperature showed lower values of the survival of the larval stage and adult, viability of pupae (%), sex ratio and total female fecundity. These results suggest the possible resistance of Vyta cultivar to T. absoluta, allowing the highest population reduction when the cultivar was subjected to alternating temperature.
These results are of great importance because it is the first time that the biological and population parameters of an exotic species are evaluated on a Cuban tomato variety previous to its introduction into the country. They must be included in the risk analysis of this phytophagous insect.
Considering the importance of this pest to Cuba and the implications of its introduction into the island, the tomato cultivar Vyta, with those characteristics suggesting its possible resistance to T. absoluta, constitutes an important arm to combat this quarantine pest in Cuba. Despite the characteristics of the Vyta cultivar related to size, texture, organoleptic properties and others not preferred by farmers, it has been used as a resistant control (TYLCV) and in genetic improvement programs for the incorporation of Ty-1 gene varieties of commercial interest.
REFERENCES
1. Filgueira FAR. Viçosa, MG: Ed. Novo manual de olericultura: agrotecnologia moderna na produção e comercialização de hortaliças. 2008. UFV, 3ed. 194-241 p.
2. FAOSTAT Database. World ranking: Tomatoes, by Production (tonnes), última actualización 16/febrero/2012. Disponible en: http://faostat.fao.org/faostat. Consultado: enero, 2015.
3. Coopercitrus. Tomaticultura lidera crescimento e lucratividade no setor de hortaliças. Revista Agropecuária, 2010. Disponível em: http://www.revistacoopercitrus.com.br/ ?pag =materia& codigo =5908. Acesso em: 23 out. 2012.
4. Bueno VH, van Lenteren JC, Lins Jr, Calixto AM, Montes F, Silva D, et al. New records of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) predation by Brazilian Hemipteran predatory bugs. J Appl Entomol. 2013;137(1-2):29-34.
5. Desneux N, Luna MG, Guillemaud T, Urbaneja A. The invasive South American tomato pinworm, Tuta absoluta, continues to spread in Afro-Eurasia and beyond: the new threat to tomato world production. Journal of Pest Science. 2011;84(4):403-408.
6. Tropea Garzia G, Siscaro G, Biondi A, Zappalà L. Tuta absoluta, a South American pest of tomato now in the EPPO region: Biology, distribution and damage. EPPO Bull. 2012;42:205-210.
7. Ponti L, Gutierrez AP, Altieri MA. Holistic management of invasive species: The case study of Tuta absoluta (Meyrick) (Lepidoptera, Gelechiidae). Atti Accademia Nazionale Italiana di Entomologia. 2013;125-136.
8. CABI Head Office, Wallingford, UK. Distribution Maps of Plant Pests, Tuta absoluta. Distribution map. 2011. Map 723 (1st revision).
9. Pereira PC, Sanchez NE. Effects of two solanaceus plants on developmental and populational parameters of the tomato leaf miner, Tuta absoluta (Lepidoptera: Gelechiidae). Neotropical Entomology. 2006;35:671-676.
10.Barrientos ZR, Apablaza HJ, Norero SA, Estay PP. Temperatura básica y constante térmica de desarrollo de la Polilla del tomate Tuta absoluta (Lepidoptera: Gelechiidae). Ciencia e Investigacion Agraria. 1998;251:33-137.
11.Souza JC, Reis PR. Traça do tomateiro, històrico, reconhecimento, biologia, prejuizios e controle. Belo Horizonte: EPAMIG. 1992. Boletin 32. 19p.
12.Karban R, Baldwin IT. Induced responses to herbivory. 2007. University of Chicago Press, 319p.
13.Lara FM. Princípios de resistência de plantas a insetos. São Paulo, Icone. 1991: 336p.
14.Lenteren JC. Biological control of pests and diseases in greenhouses: an unexpected success. In: Vincent, C., Goettel, M.S. and Lazarovits, G. (eds.). Biological Control: a Global Perspective, CABI, Wallingford. 2007:105-117.
15.Tsutomu A. Tomato as a model plant for plantpathogen interactions. Plant Biotechnology. 2007;24:135-147.
16.Gomez O, et al. Breeding for resistence to begomovirus in tropic-adapted tomato genotypes. Plant Breeding. 2004;123(3):275-279.
17.Silva D. Desenvolvimento e tabela de vida de fertilidade de Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) em regime de temperaturas constante e alternante em dois cultivares de tomate. UFLA. 2012. 75p (tese mestrado).
18.Coelho MCF, Franca FH. Biologia, quetotaxia da larva e descripçao da pupa e adulto da traça do tomateiro. Pesquisa Agropecuaria Brasileira. 1987;2(22):129-135.
19.Bogorni PC, Carvalho GS. Biologia de Tuta absoluta Meyrick (Lepidoptera: Gelechiidae) em diferentes cultivares de Lycopersicum escullentum Mill. Bioikos Campinas. 2006;20(2):49-61.
20.Maciel GM, Maluf WR, Silva VF, Gonçalves neto AC, Gomes LAA. Híbridos pré-comerciais resistentes a Tuta absoluta obtidos de linhagem de tomateiro rica em acilaçúcares. Horticultura Brasileira. 2011;29:151-156.
21.Alvares M, Moya C, Florido M, Plana D. Resultados de la mejora genética del tomate (Lycopersicon esculentum Mill.) y su incidencia en la producción hortícola de cuba. Cultivos Tropicales. 2003;24(2):63-70.
22.Mollá O, González-Cabrera J, Urbaneja A. The combined use of Bacillus thuringiensis and Nesidiocoris tenuis against the tomato borer Tuta absoluta.; Springer, Amsterdam, Netherlands, BioControl. 2011;56(6):883-891.
23.Mollá O, Biondi A, Alonso-Valiente M, Urbaneja A. A comparative life history study of two mirid bugs preying on Tuta absoluta and Ephestia kuehniella eggs on tomato crops: implications for biological control. BioControl. 2014 doi:10.1007/s10526-013-9553-8.
24.Resh VH, Carde RT. Encyclopedia of Insects. 2nd edition. Academic Press is an imprint of Elsevier. 2009. 1132p. ISBN: 978-0-12-374144-8.
25.Wikipedia, 2015. Selección sexual. Disponible en: https://es.wikipedia.org/wiki/Selecci%C3%B3n_sexual Consultado: 10/11/2014.
26.Boscan N. Cultivo masivo de insectos entomófagos con fines de control biológico. Fonaiap divulga. 1988;28. Disponible en: http://sian.inia.gob.ve/repositorio/revistas_tec /FonaiapDivulga/fd28/texto/cultivo.htm. Consultado: 10/11/2014.
27.Moreira LA, Picanço MC, Silva GA, Semeão AA, Casali VWD, Campos MR, et al. Antibiosis of eight Lycopersicon genotypes to Tuta absoluta (Lepidoptera: Gelechiidae). Revista Ceres. 2009;56(3):283-287.
28.Thomazini AP, Vendramim JD, Brunherotto R, Lopes MT. Efeito de Genótipos de Tomateiro sobre a Biologia e Oviposição de Tuta absoluta (Meyrick) (Lep.:Gelechiidae). Neotropical Entomology. 2001;30(2):283-288.
29.Ecole C, Picanço M, Moreira MD, Magalhães STV. Componentes Químicos Associados à Resistência de Lycopersicon hirsutum f. typicum a Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). An Soc Entomol Brasil. 2000;29(2):327-337.
30.Gonçalves Neto AC, Silva VF, Maluf WR, Maciel GM, Nizio DAC, et al. Resistência à traça-do-tomateiro em plantas com altos teores de acilaçúcares nas folhas. Horticultura Brasileira. 2010;28:203-208.
31.Simmons AT, Gurr GM. Trichomes of Lycopersicon species and their hybrids: effects on pests and natural enemies. Agricultural and Forest Entomology. 2005;8:1-11.
32.Leite GLD. Resistencia de tomates a pragas. Unimontes Cientifica. Montes Claros. 2004;2(6):130-140.
33.Saldúa VL, Castro AM. Antibiosis and antixenosis against the black cereal aphid (Sipha maydis) in commercial wheat cultivars. Rev Fac Agron. 2011;110(1):1-11.
34.Gilardón E, Gorustovich M, Collavino G, Hernández C, Pocoví M, et al. Resistencia de líneas de tomate a la polilla del tomate (Tuta absoluta Meyr.) en laboratorio y a campo. Invest. Agr: Prod Prot Veg. 2002;17(1):35-42.
Recibido: 25-10-2014.
Aceptado: 25-2-2015.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}