










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión On-line ISSN 2224-4697
Rev. Protección Veg. vol.31 no.1 La Habana ene.-abr. 2016
ARTÍCULO ORIGINAL
Detección y caracterización molecular del Potato virus Y (PVY) en cultivos de papa (Solanum tuberosum L.) del norte de Antioquia, Colombia
Detection and molecular characterization of Potato virus Y (PVY) in potato (Solanum tuberosum L.) crops in northern Antioquia, Colombia
Daniel Muñoz Escudero, Pablo Andrés Gutiérrez Sánchez, Mauricio Marín Montoya*
Universidad Nacional de Colombia - Sede Medellín, Facultad de Ciencias, Escuela de Biociencias, Laboratorios de Microbiología Industrial y Biología Celular y Molecular, Calle 59A No 63-20, Medellín, Colombia. Correo electrónico: dmunoze@unal.edu.co, paguties@unal.edu.co.
RESUMEN
El objetivo de este trabajo fue evaluar las técnicas de detección del Potato virus Y (PVY):TAS-ELISA e Inmunocaptura-RT-PCR en tiempo real (IC-RT-qPCR), en tejidos foliares de papa (Solanum tuberosum L.) cv. Diacol-Capiro procedentes de tres lotes del Norte de Antioquia, Colombia. Adicionalmente, se secuenció el genoma de este virus mediante secuenciación masiva de nueva generación (NGS). Se detectó el PVY con ambas técnicas en los tres lotes, aunque estas difirieron significativamente en su sensibilidad; con IC-RT-qPCR se encontró el virus en el 88,8% de las muestras, mientras que con TAS-ELISA en el 33,3%. La naturaleza de los amplicones se confirmó por secuenciación Sanger (identidad=98-100%) y comparación con los valores de temperaturas de fusión (Tm) del control positivo (77,5°C±1°C). Los análisis bioinformáticos permitieron caracterizar el genoma completo de la cepa PVY_Yarumal, con una profundidad de 716x y un tamaño de 9701nt. Presenta un marco de lectura abierto (ORF) que codifica para una poliproteína de 3061 aminoácidos, flanqueada por regiones no traducidas (UTR) 5' y 3' de 187 y 328nt, respectivamente. Los análisis filogenéticos, tanto de la poliproteína completa como de la secuencia de la cápside, indican su identidad con genotipos de la raza necrosante PVYNTN; se identificaron, además, los motivos K400E419 y QELA asociados a dicho linaje. Estos resultados plantean la necesidad de incorporar en los programas de manejo de enfermedades virales, en la agroindustria de papa en Colombia, el empleo de técnicas de detección altamente sensibles como las basadas en RT-PCR en tiempo real.
Palabras clave: ELISA, NGS, Potyvirus, RT-qPCR, Solanaceae.
ABSTRACT
The aim of this work was to evaluate the techniques TAS-ELISA and IC-RT-qPCR as detection tools for Potato virus Y (PVY) in foliar tissues of potato (Solanum tuberosum L.). Leaf samples of the potato variety Diacol-Capiro from three different plots in northern Antioquia (Colombia) were used. Additionally, the complete genome sequence of this virus was obtained using Next-generation sequencing (NGS). PVY detection using both techniques differed significantly in sensitivity, as 88.8% of the samples tested positive for PVY using IC-RT-qPCR in contrast to only 33.3% with TAS-ELISA. Sanger sequencing of the RT-qPCR amplicons and the comparison with the melting temperatures of the positive control (77.5°C±1°C) were used to confirm the presence of PVY. Bioinformatics analysis resulted in a contig of 9701 nt with an average sequence depth of 716x. The PVY_Yarumal contains an ORF coding for a 3061 residue protein flanked by 5' and 3' untranslated regions of 187 and 328 nt, respectively. Phylogenetic analysis using the complete polyprotein CDS and CP regions suggests phylogenetic identity with necrotic strain PVYNTN; sequence motifs K400E419and QELA typical of PVYNTN were also found in PVY_Yarumal. These results suggest that urgent measures are required to strengthen current integrated virus disease management in the potato agroindustry in Colombia by using highly sensitive detection techniques based on real-time RT-PCR.
Key words: ELISA, NGS, Potyvirus, RT-qPCR, Solanaceae.
INTRODUCCIÓN
El Potato virus Y (PVY), la especie tipo del género Potyvirus (Familia Potyviridae), es uno de los virus que mayormente limita la producción de papa en el mundo, al ocasionar pérdidas hasta del 80% en los rendimientos de los cultivos y afectar la calidad de los tubérculos cuando se presentan infecciones por cepas necrosantes como PVYNTN y PVYWi (1, 2). Su infección causa un amplio rango de síntomas que incluyen mosaicos de suaves a severos, rugosidad de hojas, defoliación, enanismo, necrosis foliar y de tubérculos y, finalmente, reducción en el rendimiento de las plantas (3). Este virus se transmite, de manera no persistente, por al menos 100 especies de áfidos donde Myzus persicae Sulzer es la más eficiente, al igual que por medios mecánicos y por tubérculos-semilla infectados (4). Es por esto que el establecimiento de los cultivos con material de siembra certificado libre de virus constituye un aspecto fundamental para el manejo de este y otros virus que infectan la papa (2, 5).
Los viriones de PVY se caracterizan por presentar partículas filamentosas de 730 nm de longitud y 11 nm de diámetro, con una molécula de ARN de cadena sencilla de, aproximadamente, 9700 nucleótidos (nt), con una cola de poli-A en el extremo 3´ y una proteína VPg unida, covalentemente, al extremo 5´ (4). Su genoma se expresa a partir de la traducción de una poliproteína que se procesa por tres proteasas de origen viral (NIa, HC-Pro y P1) y da origen a 10 proteínas funcionales denominadas: P1, HC-Pro, P3, 6K1, CI, 6K2, NIa-VPg, NIa-Pro, NIb y CP; las dos últimas correspondientes a la replicasa viral RdRp (RNA-dependent RNA polymerase) de 59,7 kDa y a la proteína de la cápside (CP) de 30kDa (6).
El PVY presenta altas tasas de mutación y recombinación genética, por lo que esta especie es muy variable a nivel biológico y molecular; tradicionalmente, se reconocen cinco razas principales denominadas: PVYO (raza ordinaria), PVYN (raza necrosante), PVYC (raza que causa estriados puntiformes), PVYZ (raza no necrosante) y PVYNP (no patogénica a papa), con base en sus diferencias en patogenicidad sobre papa, tabaco y pimentón (7). En la última década, a través de técnicas moleculares, se identificaron diferentes linajes genéticos asociados a cada una de esta razas; los más característicos fueron aquellos denominados PVYNTN y PVYN:O /Wi (3, 7), que corresponden a cepas que causan necrosis en tubérculos de papa y, en particular, la enfermedad denominada como «Anillo Necrótico de los Tubérculos» (Tuber Necrotic Ringspot Disease of Potato - PTNRD, por sus siglas en inglés) (7, 8).
Para la detección específica de PVY se utilizan diferentes metodologías biológicas, serológicas y moleculares. Entre las primeras se destacan la inspección visual de síntomas y pruebas de inoculación en plantas indicadoras; este proceso requiere entre seis y ocho semanas y obedece a criterios subjetivos del evaluador (9). De otra parte, las metodologías serológicas como ELISA (ensayo por inmunoabsorción ligado a enzimas), se han utilizado ampliamente para la detección del PVY al ofrecerse comercialmente anticuerpos, tanto monoclonales como policlonales; sin embargo, estas metodologías tienen restricciones de sensibilidad y diferenciación de variantes, pues son especialmente limitantes para la detección de este virus en tejidos con bajo título, como los tubérculos en latencia (5, 10). En contraste, los métodos de detección viral basados en ácidos nucleicos, como RT-PCR (Reacción en cadena de la polimerasa con retrotranscripción previa) (8, 9) y RT-PCR en tiempo real (RT-qPCR) (9,11),son altamente sensibles, de rápida ejecución y permiten el diagnóstico simultáneo de diferentes virus en formatos de detección múltiple (6).
En los últimos años se incorporaron a las herramientas de diagnóstico y caracterización molecular de virus, las de secuenciación masiva de nueva generación (NGS); los más usados, en la actualidad, son las plataformas 454 GS FLX+ (Roche), SOLiD (ABI) y HiSeq2000/2500 (Illumina) (12, 13). Con la utilización de sistemas NGS, en los últimos años se descubrieron al menos 49 virus de plantas con genomas, tanto de ARN como de ADN; 36 de los cuales se clasificaron en 16 familias virales y nueve se constituyeron en las especies tipo de igual número de nuevos géneros (13). En Colombia, el uso de metodologías NGS también permitió detectar y caracterizar molecularmente diferentes especies de virus, como son Andean potato latent virus (APLV) (14), Potato virus S (PVS) (15) y PVY (16).
Con el fin de ofrecer alternativas de detección que apoyen los programas de manejo integrado de enfermedades virales y de producción de tubérculos certificados de papa en Colombia y otros países, en este trabajo se evaluaron las técnicas para detección del Potato virus Y (PVY) de TAS-ELISA e Inmunocaptura-RT-PCR en tiempo real (IC-RT-qPCR) en tejido foliar de papa (Solanum tuberosum L.) cv. Diacol-Capiro obtenido de tres lotes del Norte de Antioquia, Colombia. Adicionalmente, se secuenció el genoma de este virus mediante secuenciación masiva de nueva generación (NGS) con el sistema NGS IlluminaHiSeq 2000, como base para su caracterización molecular en Colombia.
MATERIALES Y MÉTODOS
Muestras
En dos lotes de cultivo de S. tuberosum cv. Diacol-Capiro del municipio Santa Rosa de Osos (06°382 513 N, 75°272 373 W, altitud 2550 msnm) y en un lote ubicado en Yarumal (6°572 03 N, 75°252 13 W, altitud 2100 msnm) en el departamento de Antioquia, Colombia, se obtuvieron de manera aleatoria 15 muestras de folíolos de plantas de papa en estado de floración, con el fin de evaluar la presencia de PVY mediante pruebas de TAS-ELISA (ELISA del tipo sandwich de triple anticuerpo) e IC-RT-qPCR (RT-PCR en tiempo real con inmunocaptura previa). Adicionalmente, en el lote de Yarumal se obtuvo una muestra con síntomas típicos de la infección por PVY (mosaico severo, enanismo y deformación de brotes) con el fin de ser utilizada para la secuenciación del transcriptoma foliar por NGS.
Pruebas de TAS-ELISA e IC-RT-qPCR
Las 45 muestras foliares de folíolos de papa se evaluaron para detectar la presencia de PVY utilizando pruebas de TAS-ELISA (SRA 20001/0500) de la compañía Agdia (Indiana, EE.UU.), que emplean anticuerpos policlonales para la captura del PVY y monoclonales conjugados a fosfatasa alcalina para su revelado, siguiendo las instrucciones del fabricante. Los resultados colorimétricos se cuantificaron en un equipo Multiskan RC/MS/EX (Thermo, EEUU); en cada prueba se incluyó un control positivo y un control negativo, consistentes en tejidos vegetales suministrados en forma liofilizada por el proveedor comercial (Agdia). La definición de las muestras positivas, para la infección de PVY, se basó en la fórmula del valor de corte (Cut-off) reportada por Bioreba (http://www.bioreba.com), [Cut-off = (promedio + 3 desviaciones estándar) x 1,1]. La naturaleza viral del control positivo de PVY se confirmó previamente por RT-PCR convencional y secuenciación por el método de Sanger, con los cebadores PVYCPF (5´ACC ATC AAG SAA ATG ACA CA 3´) y PVYCPR (5´ CGG AGA GAC ACT ACA TCA CA 3´) que amplifican el gen completo (801 pb) de CP (17), siguiendo el protocolo reportado por Henao-Díaz et al. (18).
Una vez finalizadas las pruebas de TAS-ELISA, se procedió a la liberación en, todas las muestras (tanto las positivas como negativas), de las partículas virales que, putativamente, se reconocieron por los anticuerpos específicos para PVY a través del procedimiento descrito por Wetzel et al. (19), en el que se utilizan 70 µl de buffer de liberación (Tris-HCl 10mM pH 8.0, 1% Triton X-100) e incubación a 70°C durante 10 min. Estos productos de Post-ELISA se utilizaron como molde para evaluar la presencia del PVY mediante RT-qPCR con SYBR Green I. Para la síntesis del ADN copia (ADNc) se utilizaron 200 U de la enzima Maxima Reverse Transcriptase (Thermo), 1X de buffer RT, 0,5 mM de dNTPs, 100 pmol del cebador Oligo-dT, 20 U de inhibidor de ARNasas y 12,5 µl del producto de liberación post-ELISA. Las reacciones se incubaron en un termociclador T3 (Biometra, Alemania) a 65°C por 5 min, seguido de 50°C por 30 min y 85°C por 5 min. El qPCR se realizó con el kit Maxima SYBR Green/ROX qPCR Master Mix (2X) (Thermo), en 25 µl de reacción con 12,5 µl del kit, 10 µl de agua estéril libre de nucleasas, de 20 a 100 ng de ADNc y 0,3 µM de los cebadores PVY-1 FP (5´CCA ATC GTT GAG AAT GCA AAA C 3´) y PVY-1 RP (5´ATA TAC GCT TCT GCA ACA TCT GAG A 3´) (11) que amplifican una porción de 74 pb de la región CP de PVY. Todas las reacciones de RT-qPCR incluyeron un control negativo libre de cDNA viral y un control positivo de PVY, proveído por la compañía Agdia. La amplificación se realizó en un equipo Rotor-Gene Q-5plex Platform (Qiagen, Alemania) con adquisición de fluorescencia después de cada ciclo; como muestras positivas se definieron aquellas que presentaron valores de Ciclo umbral (threshold Cycle - Ct) menores de 35 y amplicones con temperatura de fusión (Tm) de ±1°C con respecto al control positivo. Todas las reacciones de RT-qPCR incluyeron un control negativo libre de ADNc viral.
Adicionalmente, la naturaleza viral de cinco de los amplicones que cumplieron con estas condiciones y del control positivo se confirmó por secuenciación Sanger en la compañía Macrogen (Corea del Sur), previa purificación con el kit QIAquick Gel Extraction (Qiagen). Las secuencias obtenidas se editaron con el programa Mega 6.0 (20) y se compararon con las bases de datos moleculares mediante el programa BLASTN (http://www.ncbi.nlm.nih.gov/BLAST).La concordancia y las diferencias significativas entre los resultados ofrecidos por las pruebas de TAS-ELISA y de IC-RT-qPCR se evaluaron mediante el índice de Cohen´s Kappa y la prueba de McNemar, respectivamente, utilizando el programa Graphpad (http://graphpad.com/quickcalcs).
Pruebas de RT-qPCR en bulks
Con el fin de evaluar la utilidad de la técnica de RT-qPCR en dos pasos para la detección de PVY directamente a partir de ARN total de tejido foliar de papa, se establecieron tres bulks de muestras por cada lote de cultivo (S1 a S5; S6 a S10 y S11 a S15) y se les realizó su maceración con nitrógeno líquido para proceder a la extracción del ARN total con el kit Gene JET Plant RNA Purification (Thermo), a partir de 100 mg de muestra y siguiendo las instrucciones del fabricante. El ARN obtenido fue eluído en 50 µl de agua tratada con DEPC y se determinaron su concentración y su pureza por lecturas de absorbancia a 260nm y 280nm en un equipo Nanodrop 2000C (Thermo), para su uso en pruebas confirmatorias de la presencia de PVY mediante RT-qPCR. Las condiciones de amplificación y de análisis de resultados fueron similares a las descritas anteriormente, así como la confirmación de la naturaleza viral de los amplicones, que en este caso se realizó para dos de las muestras.
Secuenciación NGS
Para la secuenciación del genoma completo de PVY, se partió del ARN total de una muestra foliar sintomática colectada en el municipio de Yarumal. Posteriormente, la muestra se trató con el kit TruSeq Stranded Total RNA with Ribo-Zero Plant (Illumina, EEUU) para eliminar el ARN ribosomal y luego se evalúo el número de integridad del ARN (RIN - RNA Integrity Number) a partir de lecturas en un equipo 2100 Bioanalyzer (Agilent Technologies, EEUU). Las librerías de ADNc se construyeron con el kit TruSeq RNA Simple Preparation (Illumina) y la secuenciación masiva se realizó en un equipo HiSeq 2000 (Illumina) de la compañía Macrogen. Una vez logradas las secuencias, se removieron las bases con baja calidad (Phred<30) utilizando el programa SeqTK (https://github.com/lh3/seqtk); los reads asociados a secuencias de PVY se identificaron por análisis de mapeo con respecto a genomas de referencia, a través de los programas Bowtie2 y BLASTX.
Análisis filogenéticos
Las secuencias obtenidas por NGS, para el genoma completo de PVY y por Sanger para la región CP del control positivo, se utilizaron como base para realizar análisis filogenéticos, a partir de su alineamiento con respecto a secuencias disponibles en GenBank para cepas de PVY de referencia de las razas PVYO, PVYN/NTN, PVYN:O/Wi, PVYC y PVYZ, así como con respecto a secuencias de CP de aislamientos de este virus obtenidos previamente en cultivos de papa y tomate de árbol de Colombia. Los alineamientos se realizaron con el programa Clustal W y los análisis de agrupamiento con el algoritmo de Máxima verosimilitud y el modelo GTR utilizando el programa Mega 6.0 (26). El soporte de la topología interna de las ramas del árbol se determinó por análisis de bootstrap con 500 remuestreos.
RESULTADOS Y DISCUSIÓN
Pruebas de TAS-ELISA e IC-RT-qPCR
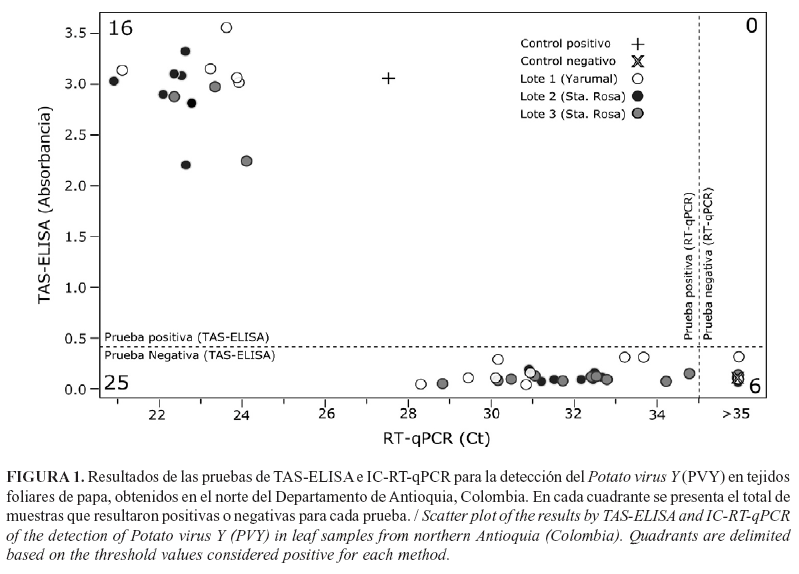
El PVY se detectó mediante pruebas de TAS-ELISA en los tres lotes de cultivos de papa, con una incidencia del 33,3% para las 45 muestras evaluadas (Fig. 1). En estas evaluaciones se definió 0,41 como el valor de corte del ensayo en el histograma de frecuencias de absorbancia a 405nm; los valores promedio alcanzados para los controles negativos fueron 0,10 (SD=0,032) y 2,9 (SD=0,42) para el control positivo comercial.
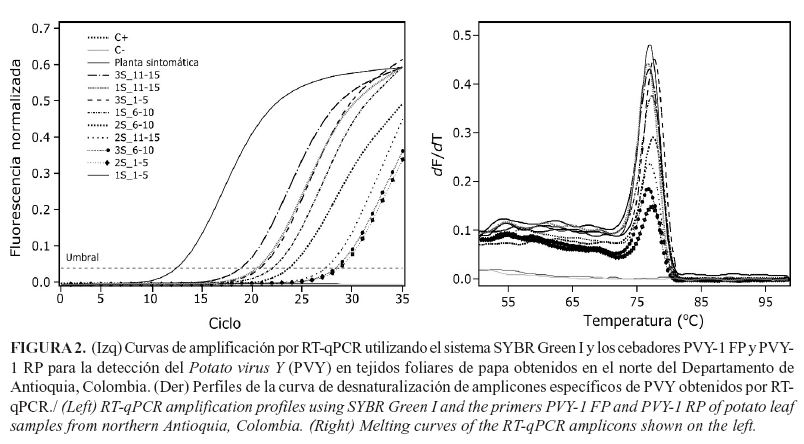
Las pruebas de IC-RT-qPCR, utilizando como molde los productos de la Post-ELISA, resultaron positivas para el 88,8% de las muestras evaluadas, con valores de Ct de 20,94 a 34,81 (promedio 28,11) para las muestras bajo análisis y de Ct=27,55 para el control positivo (Fig. 1). El análisis de las curvas de desnaturalización arrojó un valor de Tm=77,38°C para el control positivo y de Tm=77,5±1°C para las muestras (Fig.2). Estos valores son similares a los encontrados por Medina et al. (21) en un estudio tendiente a evaluar la presencia de PVY en tubérculos de papa de las variedades Diacol-Capiro y Criolla-Colombia en el departamento de Antioquia, pues en ese caso los controles positivos presentaron valores de Tm de 77,5°C +/- 0,5°C. Adicionalmente, la naturaleza viral de los amplicones generados en las reacciones de IC-RT-qPCR se confirmó por secuenciación para cinco muestras y se encontraron niveles de identidad del 98% -100% con respecto a secuencias de aislamientos de PVY reportados en GenBank (ej. KJ946936, KJ865800 y KJ847048). Asimismo, la secuencia del amplicón del control positivo presentó 100% de identidad con accesiones de PVY como KJ946936 y KJ865800.
Estos resultados señalan diferencias evidentes entre los niveles de sensibilidad ofrecidos por ambas técnicas para la detección de virus de plantas, al presentarse 25 disparidades (53,2% de observaciones) entre estas y muy bajo índice de concordancia de Cohen´s Kappa (k= 0,14; SE=0,06; intervalo de confianza al 95%: 0,023 a 0.258);se encontraron, además, diferencias significativas en la capacidad de detección de PVY por parte de ambas pruebas (McNemart=23,04, P<0,0001). Diversos reportes en virología vegetal han registrado tales diferencias; es frecuente encontrar que las metodologías basadas en RT-qPCR, con respecto a pruebas de ELISA, ofrecen niveles superiores de sensibilidad en el orden de 104 a 108 veces (9, 22). Específicamente para PVY, Kogovsek et al. (1), en un estudio sobre el diseño de pruebas de RT-qPCR para diferenciar aislamientos recombinantes de PVYNTN, registran que las pruebas en tiempo real presentaban sensibilidades superiores entre 105 y 107 veces con respecto a las pruebas de ELISA comerciales; mientras que Medina et al. (21), al evaluar la capacidad de las pruebas de RT-PCR en tiempo real para el diagnóstico de este virus en tubérculos-semilla, encontraron que esta prueba detectó el PVY en el 80,6% de las muestras, en comparación con el 37,5% detectado con pruebas de TAS-ELISA.
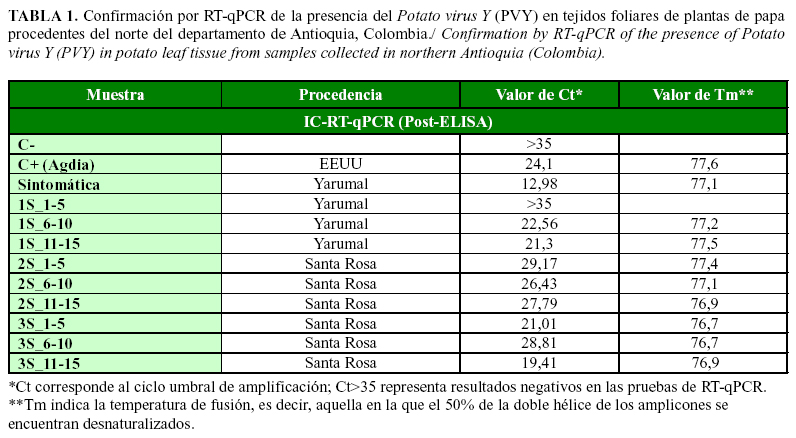
Pruebas de RT-qPCR en bulks
Las pruebas de RT-qPCR, a partir de ARN total de tejido foliar de papa, detectaron el PVY en ocho de los nueve bulks de muestras evaluadas, con valores de Ct de 19,41 a 29,17 (promedio=24,56) y de Tm de 77,1°C±0,5°C; mientras que para el control positivo y para la muestra con síntomas aparentes de infección por PVY, los valores fueron de Ct=24,1(Tm=77,6) y Ct=12,98 (Tm=77,1), respectivamente (Tabla 1). Las diferencias notables entre los valores de Ct alcanzados en las pruebas de RT-qPCR, a partir de ARN total en comparación con IC-RT-qPCR, indican la presencia de un mayor título viral en el procedimiento que utiliza extracción de ácidos nucleicos en comparación con la cantidad de partículas virales retenidas por los anticuerpos en los pozos de las placas de ELISA, por lo que se sugiere el empleo de esta primera metodología para confirmar la presencia de PVY en muestras de mayor relevancia, como por ejemplo aquellas procedentes de bancos madres de plantas de papa, en material in vitro y/o de minitubérculos, en muestras de importancia cuarentenaria o en materiales avanzados de mejoramiento genético por resistencia a virus; mientras que se sugiere emplear la IC-RT-qPCR en forma rutinaria como complemento o sustitución a las pruebas de ELISA que se utilizan tradicionalmente en los esquemas de certificación de tubérculo-semilla de papa, así como para apoyar estudios epidemiológicos y de control de este virus.
La efectividad del uso de la práctica de siembra de semilla certificada de papa por su sanidad viral se ha demostrado plenamente en países como Canadá y Estados Unidos, en los que operan estrictos sistemas estatales y privados de certificación de semilla, acompañados por el empleo de técnicas de detección altamente sensibles como RT-PCR múltiple y RT-PCR en tiempo real (5, 23); en EEUU se estima que un nivel del 10% de incidencia de PVY en material de siembra de papa representa pérdidas económicas significativas para los agricultores (24).Por su parte, Fageria et al. (9), evaluando la incidencia de PVY en lotes de cultivos de papa de New Brunswick (Canadá), reportaron que los niveles de infección en tejido foliar del 0,6% (en 2009) y 2% (en 2010) condujeron al incremento en la incidencia de este virus hasta en 20,3% (2009) y 21,9% (2010), en los tubérculos obtenidos al momento de la cosecha. Halterman et al. (5) indicaron que el aumento en la detección de cepas necrosantes de PVY en los cultivos de papa de EEUU, y la consiguiente disminución de variantes de la raza ordinaria, han puesto nuevamente a este virus como uno de los más limitantes para la agroindustria de producción de papa. Esto se debe a que las cepas necrosantes generalmente inducen síntomas foliares atenuados y difíciles de detectar visualmente, pero causan daños considerables en la calidad de los tubérculos.
Secuenciación NGS
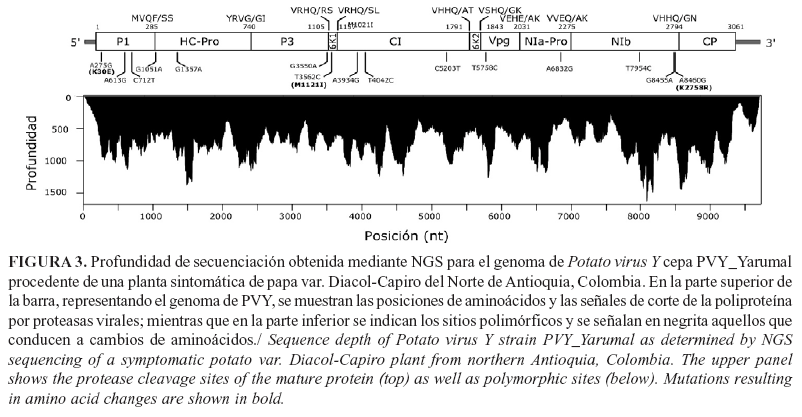
En la planta de papa var. Diacol-Capiro, con síntomas de mosaico rugoso, se confirmó la infección por PVY, tanto con las pruebas de TAS-ELISA (Absorbancia a 405 nm=2,74) como mediante RT-qPCR (Ct=12,98; Tm=77,1°C). La secuenciación NGS del transcriptoma de tejido foliar generó una librería pareada de 6.980.917reads, para un total de 1.396.183.400 nt (Fig. 3). Los análisis de mapeo contra genomas de referencia permitieron el ensamblaje con una profundidad promedio de 716x del genoma completo de PVY de 9701 nt, con PVY_Yarumal como nombre del aislamiento y KT336551 como código de accesión de Genbank.
Este genoma presenta un marco de lectura abierto (ORF) que codifica para una poliproteína de 3061 a.a., flanqueado por regiones no traducidas (UTR) 5' y 3' de 187 y 328nt (excluyendo la cola de poli-A), respectivamente. Las proteínas P1, proteinasa HC-Pro, P3, PIPO, 6K1, proteína de inclusión cilíndrica (CI), 6K2, proteína de unión al genoma (VPg), proteinasa de inclusión nuclear (NIa-Pro), replicasa viral (NIb) y proteína de la cápside (CP) tienen 285, 455, 365,247, 52, 634, 52, 188, 244, 519 y 287a.a., respectivamente; los sitios de corte proteolíticos se identificaron en los residuos 285, 740, 1105, 1157, 1791, 1843, 2031, 2275 y 2794 (Fig. 3) y son similares a los reportados por Adams et al. (25) en su análisis del clivaje de la poliproteína potyviral. El análisis de variación interna realizado entre los reads generados para cada segmento del aislado PVY_Yarumal, identificó la ocurrencia de 15 mutaciones puntuales, tres de las cuales conducen a cambios de aminoácidos en la poliproteína (K30E en P1, M1121I en 6K1 y K2758R en NIb), lo que da cuenta de las altas tasas de mutación que se han reportado en este virus en múltiples estudios moleculares (3, 4,8).
Los motivos y las regiones que caracterizan el genoma de los potyvirus se identificaron en el genoma de la cepa PVY_Yarumal e incluyeron las regiones denominadas por Turpen (26) como potyboxes A (TCAATACAACAT) y B (TCAAACAA) entre las posiciones 16-27 y 68-76 del extremo 5' UTR; los motivos conservados en HC-Pro FRNK (residuos de a.a. 463-466) e IGN (residuos 532-534 a.a.) son asociados a la supresión de silenciamiento de genes y replicación viral, respectivamente (27); la secuencia nucleotídica GA7T en la posición (2918-2926) de P3, que da origen a la proteína P3N-PIPO (247 a.a.) resultante de un cambio en el marco de lectura, fue identificado previamente por Cheng et al. (28); los motivos GSGKSTGLP (residuos 1245-1253) y DECH (residuos 1331-1334) en CI, que indican la homología de esta proteína con la superfamilia de helicasas SF2 (29); los sitios de corte de la cisteín proteasa NIa-Pro en las uniones P3/6K1, 6K1/CI, CI/6K2, 6K2/VPg, VPg/NIa-Pro, NIa-Pro/NIb, NIb/CP con secuencia de reconocimiento V-X-(HE)-(QE) (25); el motivo GDD conservado en las RdRp virales (residuos 2626-2628) y la secuencia de localización nuclear II (NSLII) TPISTPDGTIVKKFRGNNSGQPSTV (residuos 2567-2591) en NIb; finalmente, el motivo DAG (residuos 2800-2802) en la región CP, que está asociado a la transmisión por áfidos de PVY (29).
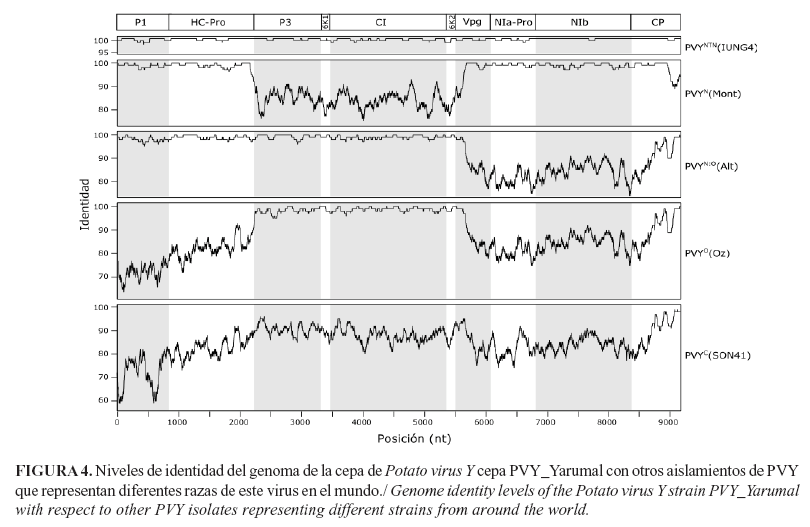
Los análisis de variación realizados entre la secuencia de PVY_Yarumal y las cepas representativas de diferentes razas de PVY que se reportan en el mundo, indican niveles de identidad de nt del 99,6% (99,8% en a.a.) y 99,2% (99,4% en a.a.) con respecto a aislamientos de PVYNTN de Polonia (IUNG-4) y Alemania (Linda), respectivamente; estas fueron las únicas regiones que presentaron valores inferiores a 99% de identidad, la 6K2 (98,7%) (con IUNG-4) y P1 (98,5%) y 6K1 (98,07%) (con Linda) (Fig. 4). Adicionalmente, la identidad del aislamiento PVY_Yarumal, como miembro de la raza N de PVY y específicamente del linaje PVYNTN, se confirmó por el hallazgo en la región HC-Pro de los dos residuos K400E419 (MKIFYPDVHDAELPRILVDHET) reportados por Tribodet et al. (30) como determinantes de los síntomas de necrosis de venas inducidos por aislamientos PVYN, así como por la secuencia de P1 RLRRQELATVRT reportada por Shubert et al. (3) en aislamientos del linaje PVYNTN.
Por otra parte, los niveles promedio de identidad entre la cepa PVY_Yarumal y los aislamientos representativos de PVYC fueron del 85,1% (92,1% en a.a.) y del 89,1% (93,4% en a.a.) con relación a PVYO; las regiones que codifican para P1 y para las proteinasas Hc-Pro y NIa-Pro fueron las más disímiles, con niveles de identidad de 72 a 82,5% (72% a 93,4% en a.a.) y 73 a 82,1% (81% a 92% en a.a.), con respecto a PVYC y PVYO, respectivamente (Fig. 4).
Análisis filogenéticos
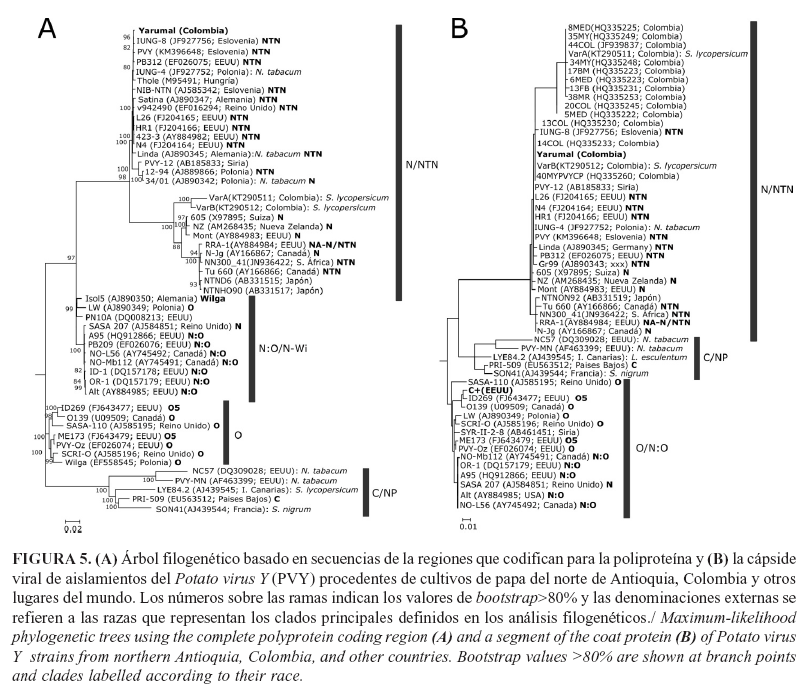
El árbol filogenético generado a partir de la secuencia completa, que codifica para la poliproteína de PVY, presentó cuatro clados principales que corresponden a los linajes que representan las razas PVYN/NTN, PVYN:O/N-Wi, PVYO y PVYC/NP, cuyos representantes provienen de diferentes países del mundo (Fig. 5A). El clado de PVYN/NTN incluyó cepas del tipo principal N y de la variante NTN y, a su vez, se presentó subdividido en dos grupos con 100% de soporte de bootstrap; el primero de ellos albergó la secuencia del aislamiento PVY_Yarumal obtenida en este trabajo, en conjunto con diferentes aislamientos de PVY procedentes de cultivos de papa de Eurasia y Norteamérica. De forma interesante, los otros dos aislamientos de PVY, completamente secuenciados en Colombia y obtenidos en plantas de tomate de cultivos de Antioquia, se ubicaron en el otro subclado de PVYN/NTN; se confirmó así la ocurrencia, en este país, de al menos dos linajes de cepas necrosantes que infectan plantas solanáceas en Colombia. Situación que infirieron Henao-Díaz et al. (18) en estudios basados en secuencias de CP de aislamientos de papa y tomate de árbol.
Por otra parte, el árbol filogenético realizado para CP presentó una topología similar al anterior, aunque las cepas de PVYO y PVYN:O se ubicaron en un solo clado, dado que las regiones de recombinación entre estas se ubican en otras regiones del genoma diferentes a CP (4). En este análisis filogenético el aislamiento PVY_Yarumal se ubicó en un subclado que incluye cepas de este virus de Colombia y de otros países del mundo, mientras que nuevamente el aislamiento de PVY de tomate reportado en GenBank (KT290511) se agrupó solo con aislamientos de PVY de Colombia, lo que reconfirma la ocurrencia de los dos genotipos necrosantes de PVY en la zona alto andina de Antioquia (Fig. 5B). Finalmente, la secuencia del control positivo comercial (Agdia) empleado en los análisis de TAS-ELISA y RT-qPCR se ubicó en el clado PVYO, lo que coincide con su origen en EEUU y confirma su utilidad en los análisis aquí realizados.
Las topologías de los árboles filogenéticos encontradas en este trabajo coinciden con los reportes de Ogawa et al. (8), en los que se utilizaron, como base de análisis, 149 secuencias de poliproteínas de PVY y donde se incluyen 20 de Japón; es recurrente en sus análisis la división del linaje PVYN/NTN en dos sublinajes que los autores denominan N-1 (N-Europa) y N-2, (N-North America); sin embargo, la presencia en este trabajo y en los de Gil et al. (31) y Henao-Díaz et al. (18), sobre un tercer linaje conformado solo por aislamientos colombianos de PVY, confirman la ocurrencia de variantes únicas de PVY en la región Andina de Colombia y sugieren la posible patogenicidad cruzada de estos aislamientos entre diferentes hospederos de plantas solanáceas, como son la papa, el tomate y el tomate de árbol; por lo que se requiere que se evalúe experimentalmente mediante pruebas de inoculación mecánica de virus en ambiente controlado.
Los resultados de la presente investigación, donde se detectó el PVY en 88,8% de 45 muestras foliares de plantas de papa de cultivos del Norte de Antioquia y aquellos recientemente reportados por Medina et al. (21), en los que se encontró que este virus utiliza RT-qPCR en 100% de 16 muestras de tubérculos de Diacol-Capiro y en 66,6% de Criolla-Colombia procedentes del departamento de Antioquia, ponen de manifiesto la urgencia de revisar los programas de manejo de enfermedades virales en la agroindustria de papa en Colombia y, muy especialmente, reforzar los programas de certificación de tubérculo-semilla con el empleo de técnicas de detección altamente sensibles, como las basadas en RT-PCR en tiempo real.
En el futuro resultará de gran interés la realización de pruebas biológicas que evalúen los efectos de las cepas necrosantes de PVY sobre los rendimientos y la calidad de los tubérculos de las variedades locales de papa que se cultivan en Colombia y en otros países andinos. Lo anterior se debe a que, hasta el momento, no está definido si la enfermedad PTNRD, causada por algunas variantes de PVYNTN, se presenta en los cultivos de esta región, pues es posible que la ocurrencia de síntomas necróticos en los tubérculos se les atribuya, equivocadamente, a otros patógenos como son Tobacco rattle virus (TRV), Potato mop-top virus (PMTV) o, incluso, a Pectobacterium solanacearum y Phytophthora infestans.
AGRADECIMIENTOS
Esta investigación se realizó con la financiación de los proyectos 19438 y 26737 de la Vicerrectoría de Investigaciones de la Universidad Nacional de Colombia y por International Foundation for Science (IFS) GrantC/4634-2.
REFERENCIAS
1. Kogovsek P, Gow L, Pompe-Novak M, Gruden K, Foster GD, et al. Single-step RT real-time PCR for sensitive detection and discrimination of Potato virus Y isolates. J Virol Methods. 2008;149:1-11.
2. Kerlan C. Potato viruses. En: Mahy BW, van Regenmorte lMH (editors). Desk encyclopedia of plant and fungal virology. Academic Press, Oxford, ReinoUnido. 2008;458-471.
3. Schubert J, Fomitcheva V, Sztangret-Wisniewska J. Diferentiation of Potato virus Y strains using improved sets of diagnostic PCR primers. J Virol Methods. 2007;140:66-74.
4. Karasev AV, Gray SM. Continuous and emerging challenges of Potato virus Y in potato. Annu Rev Phytopathol. 2013;51:571-586.
5. Halterman D, Charkowski A, Verchot J. Potato, viruses, and seed certification in the USA to provide healthy propagated tubers. Pest Tech. 2012;6:1-14.
6. Crosslin JM, Hamlin LL. Standardized RT-PCR conditions for detection and identification of eleven viruses of potato and Potato spindle tuber viroid. Am J Potato Res. 2011;88:333-338.
7. Singh RP, Valkonen JPT, Gray SM, Boonham N, Jones RAC, et al. Brief review: The naming of Potato virus Y strains infecting potato. Arch Virol. 2008;153:1-13.
8. Ogawa T, Nakagawa A, Hataya T, Ohshima K. The genetic structure of populations of Potato virus Y in Japan; based on the Analysis of 20 full genomic sequences. J Phytopathol. 2012;160:661-673.
9. Fageria MS, Singh M, Nanayakkara U, Pelletier Y, Nie X, Wattie D. Monitoring current-season spread of Potato virus Y in potato fields using ELISA and real-time RT-PCR. Plant Dis. 2013;97:641-644.
10.Crosslin JM. PVY: An old enemy and a continuing challenge. Am J Potato Res. 2013;90:2-6.
11.Singh M, Singh RP, Fageria MS, Nie X, Coffin R, Hawkins G. Optimization of a Real-Time RT-PCR assay and its comparison with ELISA, conventional RT-PCR and the grow-out test for large scale diagnosis of Potato virus Y in dormant potato tubers. Am J Potato Res. 2013;90:43-50.
12.Roossinck MJ, Martin DP, Roumagnac P. Plant virus metagenomics: Advances in virus discovery. Phytopathology. 2015;105:716-727.
13.Wu Q, Ding S, Zhang Y, Zhu S. Identification of viruses and viroids by Next-Generation Sequencing and homology dependent and homology independent algorithms. Annu Rev Phytopathol. 2015;53:1-20.
14.Kreuze J, Koenig R, De Souza J, Vetten HJ, Muller G, Flores B, et al. The complete genome sequences of a Peruvian and a Colombian isolate of Andean potato latent virus and partial sequences of further isolates suggest the existence of two distinct potato-infecting tymovirus species. Virus Res. 2013;173:431-435.
15.Gutiérrez PA, Alzate JF, Marín M. Complete genome sequence of a novel Potato virus S strain infecting Solanum phureja in Colombia. Arch Virol. 2013;158:2205-2208.
16.Villamil-Garzón A, Cuellar WJ, Guzmán-Barney M. Natural co-infection of Solanum tuberosum crops by the Potato yellow vein virus and potyvirus in Colombia. Agron Colomb. 2014;32:213-223.
17.Glais L, Tribodet M, Kerlan C. Genomic variability in Potato potyvirus Y (PVY): evidence that PVYNW and PVYNTN variants are single to multiple recombinants between PVYO and PVYN isolates. Arch Virol. 2002;147:363-378.
18.Henao-Díaz E, Gutiérrez-Sánchez P, Marín-Montoya M. Análisis filogenético de aislamientos del Potato virus Y (PVY) obtenidos en cultivos de papa (Solanum tuberosum) y tomate de árbol (Solanum betaceum) en Colombia. Actual Biol. 2013;35:219-232.
19.Wetzel T, Candresse T, Macquaire G, Ravelonandro M, Dunez J. A highly sensitive immunocapture polymerase chain reaction method for plum pox potyvirus detection. J Virol Methods. 1992;39:27-37.
20.Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: Molecular evolutionary genetics analysis. Mol Biol Evol. 2013;30:2725-2729.
21.Medina HC, Gutiérrez PA, Marín M. Detección del Potato virus Y (PVY) en tubérculos de papa mediante TAS-ELISA y qRT-PCR en Antioquia, Colombia. Bioagro. 2015;27:83-92.
22.Mumford R, Walsh K, Barker I, Boonham N. Detection of Potato mop top virus and Tobacco rattle virus using a multiplex realtime fluorescent reverse-transcription polymerase chain reaction assay. Phytopathology. 2000;90:448-453.
23.Frost K, Groves RL, Charkowski AO. Integrated control of potato pathogens through seed potato certification and provision of clean seed potatoes. Plant Dis. 2013;97:1268-1280.
24.Nolte P, Whitworth J, Thornton MK, McIntosh CS. Effect of seed borne Potato virus Y on performance of Russet Burbank, Russet Norkotah, and Shepody potato. Plant Dis. 2004;88:248-252.
25.Adams MJ, Antoniw JF, Beaudoin F. Overview and analysis of the polyprotein cleavage sites in the family Potyviridae. Mol Plant Pathol. 2005;6:471-487.
26.Turpen T. Molecular cloning of a Potato virus Y genome: nucleotide sequence homology in non-coding regions of potyviruses. J Gen Virol. 1989;70:1951-1960.
27.Shiboleth YM, Haronsky E, Leibman D, Arazi T, Wassenegger M, Whitham SA, et al. The conserved FRNK box in HC-Pro, a plant viral suppressor of gene silencing, is required for small RNA binding and mediates symptom development. J Virol. 2007;81:13135-13148.
28.Chung BY, Miller WA, Atkins JF, Firth AE. An overlapping essential gene in the Potyviridae. Proc Natl Acad Sci USA. 2008;105:5897-5902.
29.Revers F, García JA. Molecular biology of potyviruses. Adv Virus Res. 2015;92:101-199.
30.Tribodet M, Glais L, Kerlan C, Jacquot E. Characterization of Potato virus Y (PVY) molecular determinants involved in the vein necrosis symptom induced by PVYN isolates in infected Nicotiana tabacum cv. xanthi. J Gen Virol. 2005;86:2101-2105.
31.Gil JF, Cotes JM, Marín M. Incidencia de potyvirus y caracterización molecular de PVY en regiones productoras de papa (Solanum tuberosum L.) de Colombia. Rev Col Biotecnol. 2011;85-93.
Recibido: 28-7-2015.
Aceptado: 6-2-2016.
* Autor para correspondencia: Mauricio Marín Montoya. Correo electrónico: mamarinm@unal.edu.co.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}