










Meu SciELO
 Serviços customizados
Serviços customizadosServiços Personalizados
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Protección Vegetal
versão On-line ISSN 2224-4697
Rev. Protección Veg. vol.31 no.1 La Habana jan.-abr. 2016
COMUNICACIÓN CORTA
Variabilidad molecular de aislados de Xanthomonas axonopodis pv. phaseoli procedentes de la provincia Mayabeque, Cuba
Molecular variability of Xanthomonas axonopodis pv. phaseoli isolates from Mayabeque Province, Cuba
Mylene Corzo López, Odaylin Plasencia Márquez, Benedicto Martínez Coca, Madelaine L. Quiñones Pantoja
Grupo de Fitopatología, Dirección de Sanidad Vegetal, Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, Mayabeque, Cuba. Correo electrónico: mylene@censa.edu.cu.
RESUMEN
El objetivo del trabajo fue caracterizar molecularmente, mediante rep-PCR, aislados de Xanthomonas axonopodis pv. phaseoli procedentes de diferentes áreas productoras de frijol común (Phaseolus vulgaris L.) en la provincia Mayabeque. Se obtuvieron 29 aislados a partir de 30 muestras sintomáticas colectadas, a los que se le determinó, previamente, las características morfológicas, bioquímicas-fisiológicas y patogénicas. Se detectó una elevada variabilidad genética entre los aislados analizados, pues se formaron cuatro grandes grupos genéticos con un coeficiente de similitud entre ellos de 73% y un gran número de haplotipos o subgrupos (22 haplotipos). Los cuatro grupos génicos estuvieron integrados por aislados que causan síntomas diferentes. No se observó una relación directa entre la variabilidad genética presente entre los aislados y las características patogénicas presentes en condiciones semicontroladas. Se demostró la amplia variabilidad genética que existe en los aislados de X. axonopodis pv. phaseoli asociados a diferentes cultivares de frijol común en condiciones de campo.
Palabras clave: rep-PCR, Tizón común bacteriano, Phaseolus vulgaris.
ABSTRACT
The aim of this work was the molecular characterization by rep-PCR of isolates of Xanthomonas axonopodis pv. phaseoli from different common bean (Phaseolus vulgaris L.) producing areas in the province Mayabeque. Twenty-nine isolates were obtained from thirty symptomatic samples collected, to which the biochemical, physiological, morphological, and pathogenic characteristics were previously determined. A high genetic variability was detected among the analyzed isolates, which formed four major genetic groups with a coefficient of similarity of 73% between them and a high number of haplotypes or subgroups (22 haplotypes). The four gene groups were composed of isolates that caused different symptoms. No direct relationships were observed between this genetic variability among isolates and the pathogenic characteristics present under controlled conditions. The wide genetic variability that exists in isolates of Xanthomonas axonopodis pv phaseoli associated with different cultivars of common bean under field conditions was shown.
Key words: rep-PCR, common bacteria bligth (CBB), Phaseolus vulgaris.
El Tizón común o Bacteriosis común es una de las enfermedades más destructivas en el cultivo del frijol (Phaseolus vulgaris L.) en zonas de climas templados y tropicales (1). Esta enfermedad es provocada por las bacterias Xanthomonas axonopodis pv. phaseoli (Xap) y su variante Xanthomonas axonopodis pv. phaseoli var. fuscans (Xapf), denominada también como Xanthomonas fuscans subsp. fuscans (Xff) (2).
Las bacterias X. axonopodis pv. phaseoli y X. fuscans subsp. fuscans se encuentran ampliamente distribuidas por el mundo, ya que la trasmisión por semillas es la principal fuente de infección (4), independiente de la resistencia de los cultivares a la enfermedad (3). Indudablemente, el manejo integrado de la enfermedad en campo tiene como base el uso de semillas libres del patógeno, de cultivares resistentes, la rotación del cultivo y la eliminación de restos de cosechas (5).
La variabilidad genética de estas bacterias se estudió por diversos investigadores debido a la importancia económica que presentan estos fitopatógenos. Disímiles son las técnicas moleculares utilizadas para ello: la Amplificación Aleatoria de ADN Polimórfico (RAPD, del inglés Random Amplified Polymorphic ADN Fragment), el Polimorfismo de la Longitud del Fragmento de Restricción (RFLP, del Inglés Restriction Fragment Length Polymorphism), Polimorfismo de la Longitud del Fragmento Amplificado (AFLP, del inglés Amplified Fragment Length Polymorphism), así como la Reacción en Cadena de la Polimerasa de Elementos Repetitivos (rep-PCR, de inglés repetitive element-PCR), entre otras (6,7,8).
El rep-PCR es una técnica que se usa para identificar, diferenciar y determinar la variabilidad genética entre bacterias patógenas. Este método utiliza cebadores que reconocen secuencias altamente conservadas, repetidas y dispersas por todo el genoma bacteriano; con la técnica del PCR se amplifica la secuencia de ADN que se encuentra entre estos elementos repetidos (9). Con esta técnica, Weingart et al. y Vauterin et al. diferenciaron patovares de Pseudomonas syringae (10) y Xanthomonas campestris (11), respectivamente. Mkandawire et al. (7) identificaron tres aislados de X. axonopodis pv. phaseoli; dos de ellos se agruparon con aislados colectados en el Este de África, mientras que el otro se agrupó con cepas de otras localidades. Posteriormente, en estudios realizados en España con esta misma técnica, López et al. (6) revelaron la existencia de una ligera variabilidad genética entre aislados de X. axonopodis pv. phaseoli y la similitud de estos con aislados del Nuevo Mundo.
En Cuba, Rodríguez et al. (12), utilizando la técnica de rep-PCR con los cebadores Eric y Box, diferenciaron aislados de Xap y de Xff; también analizaron la variabilidad genética presente entre los aislados, los que manifestaron una baja variabilidad genotípica entre ellos. Recientemente, Corzo et al. (13) obtuvieron nuevos aislados de Xap, que mostraron diferencias en los síntomas de la enfermedad expresados en el cultivar de frijol BAT-304, bajo condiciones semicontroladas. La variabilidad patogénica arrojó como resultados la agrupación de dichos aislados en cuatro grupos sintomatológicos (G1, G2, G3 y G4), lo que pudiera estar relacionado con posibles diferencias genotípicas en los mismos.
El objetivo del trabajo fue caracterizar molecularmente, mediante rep-PCR, aislados de Xanthomonas axonopodis pv. phaseoli procedentes de diferentes áreas productoras de frijol común en la provincia Mayabeque, Cuba.
Los 29 aislados utilizados provienen de muestras foliares (trifolios ubicados en la parte media de la planta) y vainas con síntomas similares a los de Bacteriosis común, colectadas en las fases de prefloración, floración y formación de la vaina, en el periodo comprendido de septiembre de 2013 a marzo de 2014, en los municipios San José de las Lajas, Quivicán y Güines, pertenecientes a la provincia Mayabeque, Cuba (Tabla 1).
Para la caracterización molecular de los aislados se procedió, primeramente, a la obtención de una suspensión bacteriana de cada aislado, con una concentración de 108UFCx ml-1 a una DO de 600nm (Espectrofotómetro T60 UV PG Instruments). Para lo cual se tomaron 2 ml que se inocularon en 5 ml de Caldo Nutriente (CN) y se incubaron a 28oC de 12 a 18h. Del medio inoculado, se tomaron 2 ml y se centrifugaron a 8 000 rpm por 5 min en una centrífuga Eppendorf. El precipitado celular se lavó tres veces con 1 ml de agua destilada estéril y se centrifugó a 8 000 rpm. La extracción del ADN total del cultivo bacteriano se realizó según lo descrito por Zamari et al. (14). Los ADN se conservaron a -200C hasta su posterior uso en la PCR. Los ADN obtenidos se analizaron en el ensayo de rep-PCR (11) para lo cual se utilizaron los cebadores ERIC, BOX y REP (Tabla 2).
El ensayo de amplificación se realizó en una mezcla de reacción de 25 ml de volumen final, la cual contenía 1,25m GoTaqÒ G2 Green Master Mix (Promega): 1X Green GoTaqÒ G2 Reacción Buffer (pH 8,5), 200 mM de dNTP y 1,5 mM de MgCl2, 0,2 mM de cada cebador y 2 ml de ADN molde. La reacción de amplificación se realizó según el protocolo descrito por la EPPO en 2010 (11). Como control negativo de la reacción se usó agua libre de RNAsa. Los fragmentos de ADN amplificado se visualizaron por irradiación con luz ultravioleta en un transiluminador (Syngene InGenius L, Syngene, EE.UU) sobre un gel de agarosa al 2% en TBE al 0,5X, teñido con bromuro de etidio. Se usó el marcador de peso molecular de 100 pb (Ladder, Promega). El ensayo se realizó por triplicado.
Los patrones de bandas, alcanzados por el cebador que aportó mayor cantidad de bandas polimórficas, se observaron directamente en la imagen del gel y se transformaron en una matriz binaria (1 presencia de la banda, 0 ausencia de la banda). Con los datos de esta matriz se calculó la similitud genética entre los aislados utilizando el coeficiente de Dice con el programa NTSys pc 2.11x (16). La matriz de similitud se usó para construir el dendrograma por el método de la Media Aritmética No Ponderada (UPGMA, Unweighted Pair -Group Method Aritmetic Average) mediante el algoritmo SAHN (Sequential Agglomerative Hierarchical and Nested) del programa anteriormente mencionado.
Para determinar la posible asociación entre los grupos sintomatológicos (G1, G2, G3 y G4) y los grupos genéticos creados a partir del análisis del dendrograma con los datos arrojados por la técnica rep-PCR, se realizó una prueba de Chi cuadrado independiente, utilizando el programa InfoStat versión 2.0 (17). Se consideraron valores significativos de p£0,05.
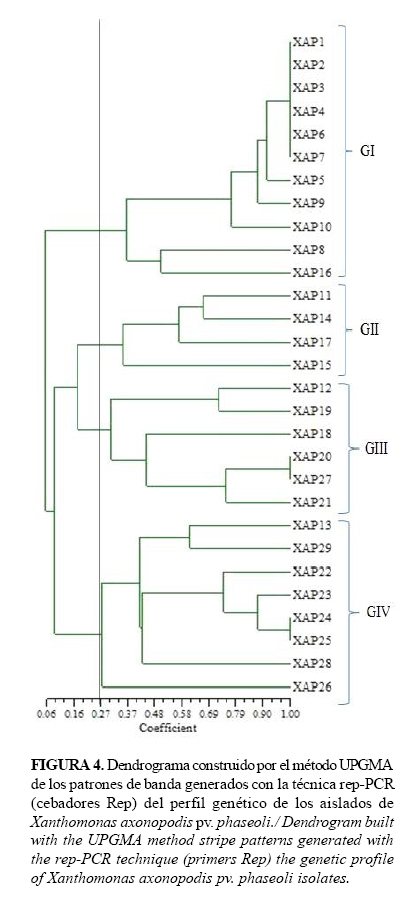
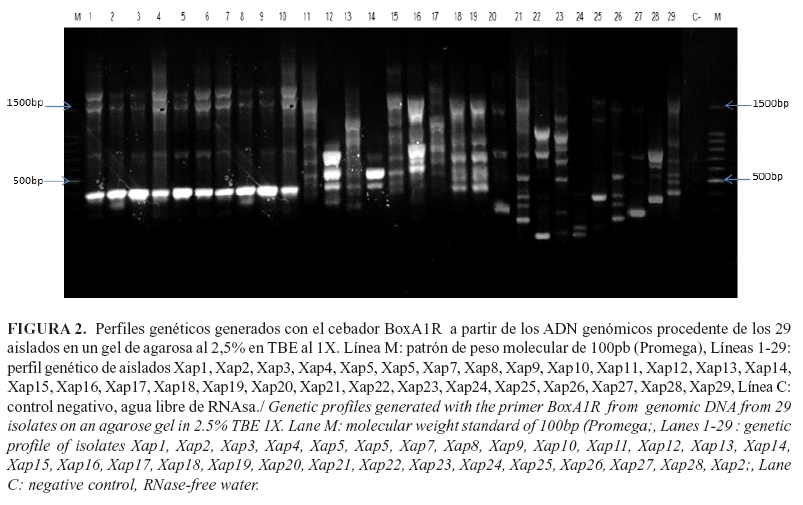
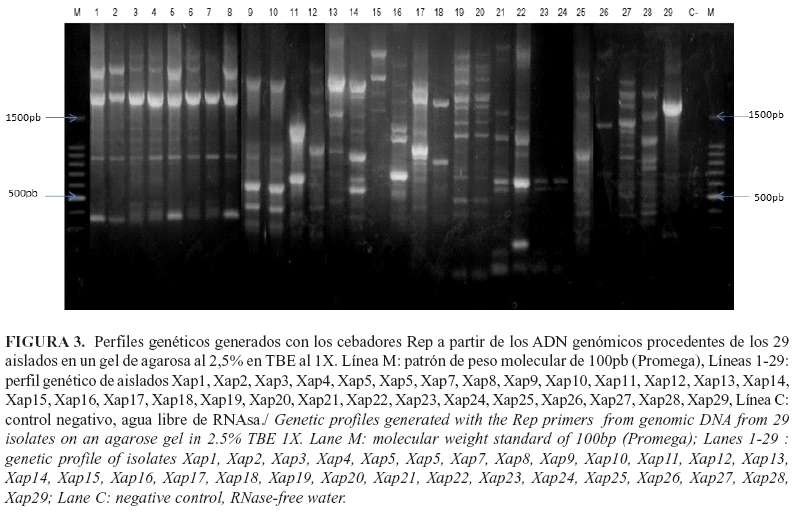
Con el uso de los cebadores Eric, Box y Rep y la técnica rep-PCR, se amplificaron secuencias repetidas del genoma de los aislados de Xap, las cuales mostraron un elevado número de bandas polimórficas con tallas entre 150 pb y 2 000 pb. Con los cebadores Eric y Box se obtuvieron 33 bandas polimórficas, respectivamente (Fig.1, Fig.2). Sin embargo, con los cebadores Rep se visualizaron 44 bandas polimórficas (Fig.3), los que aportaron el mayor polimorfismo entre los aislados.
Los valores del coeficiente de similitud de Dice, correspondiente al análisis del patrón de bandas obtenido con los cebadores Rep, estuvo en el rango de 0,06 y 1. Estos resultados muestran la elevada variabilidad genética presente entre los aislados de los diferentes municipios de una misma provincia y coinciden con los resultados de Mkandawire et al. (7) y Bett et al. (17), quienes observaron alta variabilidad genética entre aislados de Xap.
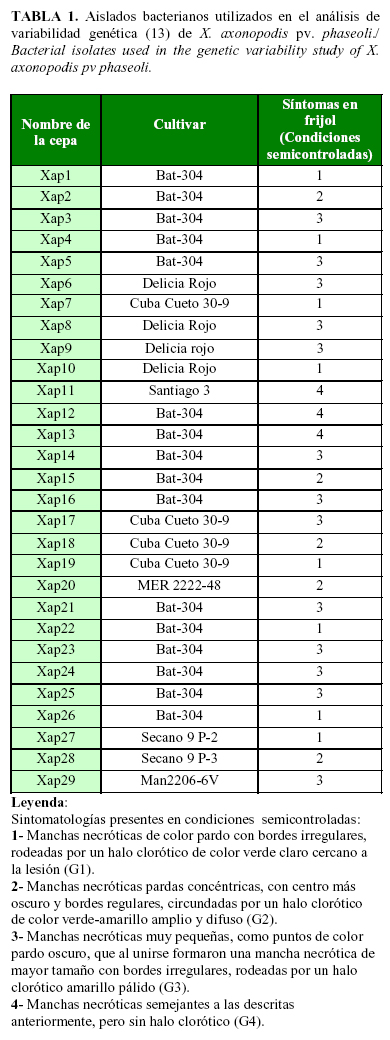
El análisis del dendrograma (Fig. 4) mostró la formación de cuatro grandes grupos genéticos con un coeficiente de similitud (CS) de 73% entre ellos; en cada grupo se ubicaron aislados de diferentes municipios de la provincia Mayabeque. El primer grupo (GI) representa 11 aislados de los municipios San José de las Lajas, Quivicán y Güines, en los cuales se concentran seis haplotipos o subgrupos con un CS de 63%. El segundo (GII) agrupa cuatro aislados de los municipios San José de las Lajas y Güines, y cada uno representa un haplotipo con un CS de 65%. El tercer grupo (GIII) está formado por seis asilados de los municipios San José de las Lajas y Güines, lo integran cinco haplotipos con un CS de 70%. El cuarto grupo (GIV), está constituido por ocho aislados de los municipios San José de las Lajas y Güines y presentan 7 haplotipos con CS de 71%. Estos resultados indican que el patovar phasoli es altamente heterogéneo y presenta una elevada variabilidad genética en las plantaciones de frijol en la provincia Mayabeque.
Los resultados expuestos se corresponden con los informados por Mkandawire et al. (7) entre aislados procedentes de diferentes regiones del Este de África, así como los de López et al. (6) con aislados originarios de diferentes territorios de España y Bett et al. (16) con aislados procedentes de diferentes regiones de Canadá, los cuales informaron la elevada heterogeneidad genética presente entre los aislados de Xap.
En los cuatro grupos genéticos (GI, GII, GII, GIV) se distribuyen todos los grupos sintomatológicos (G1, G2, G3 y G4), lo que no permitió detectar una relación directa entre la variabilidad genética y la diversidad de síntomas de la enfermedad observada en condiciones semicontroladas. Esto se corroboró con los resultados en la prueba Chi cuadrado independiente, donde los valores de p no fueron significativos (p=0,4547), lo que demostró que no existe una asociación entre dichos grupos.
La técnica de rep- PCR sirvió para caracterizar molecularmente los aislados de Xap utilizados; sin embargo, no fue lo suficientemente resolutiva para determinar la correlación entre la variabilidad genética presente entre los aislados y los diferentes síntomas expresados en condiciones semicontroladas en el cultivar Bat 304. Por tanto, se recomienda el uso de otras técnicas moleculares como AFLP (18) y MLST (Tipificación por Secuencias de Múltiples genes) (19, 20), que permitan explorar otras zonas del genoma bacteriano, las cuales pudieran estar relacionadas con la diversidad de síntomas observados.
El conocimiento de la variabilidad genética de los patógenos que circulan en las áreas productoras de frijol común del país es un elemento a tener en cuenta para perfeccionar, tanto el programa mejoramiento como el manejo integrado de este cultivo (1,14).
REFERENCIAS
1. Mederos Y. Revisión bibliográfica: Indicadores de la calidad en el grano de frijol (Phaseolus vulgaris L.). Instituto Nacional de Ciencias Agrícolas. 2013;27:55-62.
2. Ah-You N, Gagnevin L, Grimont PAD, Brisse S, Nesme X, Chiroleu F, et al. Polyphasic characterization of Xanthomonas pathogenic to members of the Anacardiacae and their relatedness to the species of Xanthomonas. Int J Syst Evol Microbiol. 2009;59:306-318.
3. Popoviæ T, Balaž J, Gavriloviæ V, Aleksiæ G. Distribution and characterization of phytopathogenic bacteria on commercial bean crop in Vojvodina Serbia. Plant Protection. 2009;60:101-125.
4. Darrasse A, Bureau C, Samson R, Morris CE, Jacques MA. Contamination of bean seeds by Xanthomonas axonopodis pv. phaseoli associated with low bacterial densities in the phyllosphere under field and greenhouse conditions. Eur J Plant Pathol. 2007;119:203-215.
5. Arayas M. Guía de identificación y manejo integrado de enfermedades del frijol en América Central. 3th ed. Tegucigalpa. Honduras. 2012. p.1-37.
6. Lopez R, Asensio C, Gilbertson RL. Phenotypic and genetic diversity in strains of common blight bacteria (Xanthomonas campestris pv. phaseoli and X. campestris pv. phaseoli var. fuscans) in a secondary center of diversity of the common bean host suggests multiple introduction events. Phytopathology. 2006;96:1204-1213.
7. Mkandawire BC, Mabagala RB, Guzman P, Gepts P, Gilbertson RL. Genetic diversity and pathogenic variation of common blight bacteria (Xanthomonas campestris pv. phaseoli and X. campestris pv. phaseoli var. fuscans) suggests pathogen coevolution with the common bean. Phytopathology. 2004;94:593-603.
8. Mahuku GS, Jara C, Henriquez MA, Castellanos G, Cuasquer J. Genotypic characterization of the common bean bacterial blight pathogens Xanthomonas axonopodis pv. phaseoli and X. axonopodis pv. phaseoli var. fuscans by rep-PCR and PCR-RFLP of the ribosomal genes. J Phytopathol. 2006;154:35-44.
9. OEPP/EPPO. Rep-PCR tests for identification of bacteria. Bulletin OEPP/EPPO Bulletin. 2010;40:365-368.
10.Weingart H, Volksch B. Genetic fingerprinting of Pseudomonas syringae pathovars using ERIC-, REP-, and IS50-PCR. J Phytopathol. 1997;145:339-345.
11.Vauterin L, Rademaker J, Swings J. Synopsis on the taxonomy of the genus Xanthomonas. Phytopathology. 2000;90:677-682.
12.Rodríguez MO. Evaluación de la reacción de cultivares y líneas de frijol ( Phaseolus vulgaris L.) a Bacteriosis cumún ( Xanthomonas axonopodis pv. pahseoli) e identificación de marcadores de interés para este carácter. Tesis de Doctor en Ciencias Agrícolas. Universidad Agraria de La Habana. 2011: 103 pp.
13.Corzo LM, Rivero GD, Zamora GL, Martínez ZY, Martínez CB. Detección e identificación de nuevos aislados de Xanthomonas axonopodis pv. phaseoli en cultivares de frijol común (Phaseolus vulgaris L.) en la provincia Mayabeque, Cuba. Rev Protección Veg. 2015;30(2):97-103.
14.Zamani Z, Bahar M, Jacques MA, Lak MR, Akhava A. Genetic diversity of the common bacterial blight pathogen of bean, Xanthomonas axonopodis pv. phaseoli, in Iran revealed by rep-PCR and PCR-RFLP analyses. World J Microbiol Biotechnol. 2011;27(10):2371-2378.
15.Versalovic J, Schneider M, De Bruijn FJ, Lupski JR. Genomic fingerprinting of bacteria using repetitive sequence based polymerase chain reaction. Methods in Molecular Cell Biology. 1994;5:25-40.
16.NTSYSpc. Numerical Taxonomy and Multivariate Analysis System User's Guide. 2005 Exeter Software, Setauket, NY.
17. Di Rienzo JA, Casanoves F, Balzarini MG, Gonzalez L, Tablada M, Robledo CW. 2010. InfoStat versión 2.0. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina.
18.Bett KE, Banniza S. Population study of Xanthomonas spp. from bean growing regions of Canada and response of bean cultivars to pathogen inoculation. Can J Plant Pathol. 2014;36(3):341-353.
19.Mayer L, Padilha da SW, Bittencourt MA, Tondo VC. AFLP analysis of Xanthomonas axonopodis and X. arboricola strains used in Xanthan production studies reveal high levels of polymorphism. Brazilian Journal of Microbiology. 2010;41:741-748.
20.Young JM, Park DC, Shearman HM, Fargierc E. A multilocus sequence analysis of the genus Xanthomonas. Systematic and Applied Microbiology. 2008;31:366-377.
21.Fargierc E, Fischer-Le Saux M, Manceau C. A multilocus sequence analysis of Xanthomonas campestris reveals a complex structure within crucifer-attacking pathovars of this species. Systematic and Applied Microbiology. 2011;34:156-165.
Recibido: 14-12-2015.
Aceptado: 24-3-2016.


{kind=link}
{kind=link}
{kind=link}
{kind=link}