










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión impresa ISSN 1010-2752versión On-line ISSN 2224-4697
Rev. Protección Veg. vol.31 no.2 La Habana mayo-ago. 2016
ARTÍCULO ORIGINAL
Caracterización molecular de la especie invasiva Middle East-Asia minor one (MEAM1) de Bemisia tabaci (Gennadius), presente en Cuba
Molecular characterization of Bemisia tabaci (Gennadius), Middle East-Asia minor one (MEAM1) invasive species in Cuba
Yamila Martínez-ZubiaurI*, Yenne Marrero AlvarezI, Carlos Pupo FeriaII, Ileana Miranda CabreraI, Iván Galindo-CastroIII
ICentro Nacional de Sanidad Agropecuaria, CENSA, San José de las Lajas, Mayabeque, Cuba.
IICentro Universitario de las Tunas.
IIIInstituto de Estudios Avanzados, IDEA, Venezuela.
RESUMEN
En este trabajo se utilizaron marcadores microsatélites y la secuencia del gen mtCOI para realizar una caracterización molecular de la especie Middle East-Asia minor one (MEAM1) de Bemisia tabaci (Gennadius) (Hemiptera, Aleyrodidae), en Cuba. Se seleccionaron individuos recolectados en pepino (Cucumis sativa L.), remolacha (Beta vulgaris L.), tomate (Solanum lycopersicum L.) y calabaza (Curcubita pepo L.); en frutales de interés económico como fruta bomba (Carica papaya L.) y en dos especies de arvenses hospedantes de Bemisia como hierba mora (Solanum americanum Mill) y salvia (Salvia officinalis L), en ecosistemas no productivos. Los resultados permitieron confirmar la presencia de la especie MEAM1 en todas las especies de plantas estudiadas, con baja divergencia molecular no asociada al hospedante. La secuenciación evidenció la presencia de las MEAM1 coexistiendo con la especie New World (NW) en Salvia officinalis L.
Palabras clave: Bemisia tabaci, MEAM, marcadores microsátelites, gen mtCOI.
ABSTRACT
Microsatellite markers and the mitochondrial sequence of mtCOI gene were used for the molecular characterization of the MEAM1 species of Bemisia tabaci (Gennadius) (Hemiptera, Aleyrodidae) in Cuba. Whitefly specimens collected from cucumber (Cucumis sativa L.), sugar beet (Beta vulgaris L.), tomato (Solanum lycopersicum L.), squash (Cucurbita pepo L), fruits of economic interest like papaya (Carica papaya L.), and two species of Bemisia weed hosts like American black nightshade (Solanum americanum Mill) and sage (Salvia officinalis L) were selected in non-productive ecosystems The results made evidente the presence of MEAM1 population with low molecular divergence in all hosts examined. DNA sequencing revealed the coexistence of MEAM1 with the New World (NW) species in S. officinalis L in natural habitat.
Key words: Bemisia tabaci, MEAM1, microsatellites marker, mtCOI gene.
INTRODUCCIÓN
La mosca blanca (Bemisia tabaci Genn.) es una especie haplodiploide, descrita por primera vez como plaga en la agricultura hace 120 años en Grecia y desde entonces es una de la más dañinas, pues afecta múltiples especies de plantas, tanto arvenses como cultivos de interés económico. Es una plaga cosmopolita ampliamente distribuida por las regiones tropicales y subtropicales que causa daños directos a las plantas y es vector de más de 110 virus de plantas (1).
En la literatura se refieren varios métodos moleculares que se utilizan para la caracterización de las poblaciones de mosca blanca; todos revelan una amplia variabilidad de biotipos y grupos filogenéticos: desde los patrones de esterasas, la amplificación aleatoria de ADN polimórfico (RAPD del inglés Random Amplification of Polymorphic DNA), el análisis de polimorfismos en la longitud de fragmentos amplificados (AFLP, del inglés Amplified fragment length polymorphism) hasta la secuenciación de la región mitocondrial y ribosomal. El estudio de diferentes locis microsatélites ha brindado una mayor resolución a las investigaciones acerca de las estructuras de las poblaciones y ofrece las primeras evidencias de que al menos 10 especies indistinguibles morfológicamente se encuentran en la región de Asia-Pacífico (1, 2).
Posteriormente, con la secuenciación del gen mitocondrial citocromo oxidasa (mtCO1) se concluyó que B. tabaci es un complejo formado por, al menos, 24 especies crípticas imposibles de distinguir morfológicamente, entre los que están incluidos 11 grupos genéticos de mayor nivel y diferenciación evolutiva; estos grupos los asociaron a los diferentes biotipos empleados, lo que confiere mayor valor genético y propone la eliminación de la denominación de biotipos (3, 4). A continuación, en cursiva se describen las especies propuestas y los biotipos asociados entre paréntesis: Mediterranean (Q, J, L, sub-Saharan Africa silverleaf); Middle EastAsia minor 1 (B, B2); Middle East-Asia minor 2; Indian Ocean (MS); Asia I (H, M, NA); Australia/Indonesia; Australia (AN); China 1 (ZHJ3); China 2; Asia II 1 (K, P, ZHJ2); Asia II 2; Asia II 3 (ZHJ1); Asia II 4; Asia II 5 (G); Asia II 6; Asia II 7 (Cv); Asia II 8; Italy (T); sub-Saharan Africa 1; sub-Saharan Africa 2 (S); sub-Saharan Africa 3; sub-Saharan Africa 4; New World (A, C, D, F, Jatropha, N, R, Sida); y Uganda.
En Cuba se informó, en 1995, la presencia de B. argentifolli (5); más tarde la caracterización molecular de poblaciones cubanas de mosca blanca, fundamentalmente en tomate, permitió determinar la presencia del biotipo B de B. tabaci, aunque no se encontró el biotipo A o especie nativa (6) en las plantas analizadas. En 2011 (7) se informó la especie MEAM1 (previamente biotipo B) de forma predominante en cultivos hortícolas y en Malva sp., asociada al cultivo del tomate.
El presente trabajo tuvo como objetivos conocer la distribución de la especie MEAM1 en diferentes hospedantes y ecosistemas y determinar, molecularmente, su variabilidad genética.
MATERIALES Y MÉTODOS
Se recolectaron 284 adultos de mosca blanca de pepino (Cucumis sativa L.), remolacha (Beta vulgaris L), tomate (Solanum lycopersicum L.) y calabaza (Curcubita pepo L.), además de un cultivo no hortícola, que fue fruta bomba (Carica papaya L.). También se recolectaron insectos en dos especies de arvenses hospedantes de Bemisia sp., presentes en ecosistemas no productivos; estas fueron: salvia (Salvia officinalis L.) y hierba mora (Solanum americanum Mill).
El ADN se extrajo de adultos de B. tabaci, según el protocolo propuesto por Delatte et al. (8), colocando una sola mosca en el tubo eppendorf de 1,5ml y macerando fuertemente con una varilla de cristal. El tubo se calentó a 95oC por 5 minutos y se puso rápidamente a frío. Posteriormente se centrifugó a 8000 g por 30 segundos y el sobrenadante se transfirió a un tubo limpio y almacenado a -80oC hasta su uso.
Con el objetivo de descartar la presencia de la especie Mediterránea (MED, anteriormente biotipo Q) y de agrupar los ADN correspondientes a la especie MEAM1 (anteriormente biotipos B) y separarlos de la especies no MEAM1. Se analizó cada ADN procedente de los adultos mediante la amplificación de dos locis microsatélites, Bem23 y Ms145 (7). Los fragmentos se separaron por electroforesis en geles de agarosa al 2%.
Los ADN positivos para el grupo MEAM1 se utilizaron para estudiar el polimorfismo genético de los individuos colectados en diferentes hospedantes, mediante el análisis de tres locis microsatélites (Bem6, Bem15 y Bem25) descritos por De Barro et al. (2).
Las amplificaciones se realizaron en una mezcla de volumen final 20 µl que contiene 100 µM dNTP, 0.1µM de cada cebador, 4 mM MgCl2, 1 U de Taq polimerasa y 2µl de ADN extraído. Todos los locis microsatélites se amplificaron con el siguiente programa 35 ciclos de 1 min a 94 °C, 1 min a 55 °C, 1 min a 73 °C, 10 µl de cada producto amplificado fue visualizado en un gel de agarosa al 2%. Los productos obtenidos de los cebadores de los locis Bem 6, Bem15 y Bem25 se visualizaron en geles de poliacrilamida al 7%, en condiciones nativas.
Con los resultados de las amplificaciones se construyó una matriz de datos binarios sobre la cual se realizó el cálculo de las distancias mediante índice de similaridad de Jaccard y el análisis de conglomerados utilizando el método de UPGMA (programa InfoStart) (9). Los dendrogramas permitieron evaluar la variabilidad genotípica entre los individuos seleccionados, correspondientes a la especie MEAM1.
Para comprobar los resultados con el estudio de los marcadores se seleccionaron cinco ADN genómicos, dos que amplificaron el patrón MEAM1 y tres que no mostraron patrón MEAM1. Estos ADN se amplificaron con los cebadores C1-J2195/L2-N3014 (10) del gen mitocondrial citocromo oxidasa (mtCO1), según el protocolo de amplificación propuesto por Muñiz et al. (7). Los fragmentos obtenidos se secuenciaron y se analizaron con el software BLAST (http://www.ncbi.nlm.nih.gov).
RESULTADOS Y DISCUSIÓN
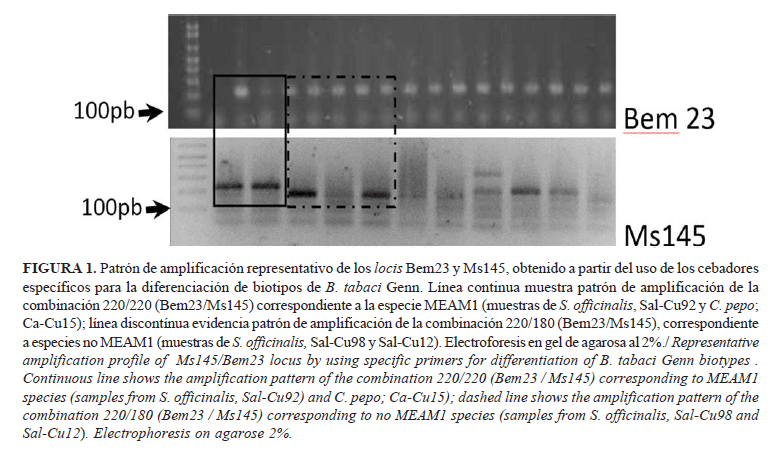
En la Figura 1 se observan los patrones representativos amplificados para los marcadores Ms145 y Bem 23 que demostraron dos tipos de combinaciones: 1) Patrón de 220pb para ambos marcadores y 2) Patrón de 220 pb para Bem 23 y 180pb para Ms145. La combinación de alelos de los locis Bem23/Ms145 con el patrón 220/220 coincide con el informado para el grupo MEAM1, referido a la presencia del alelo 220 en homocigosis. La amplificación de individuos con patrones 220/180 observados coincide con los referidos a especies no MEAM1 (7). En las muestras analizadas no se detectó la presencia de MED (biotipo Q) con la combinación 410/180.
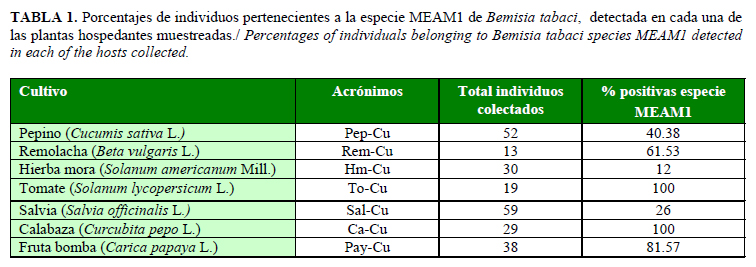
En la Tabla 1 se aprecian los resultados del análisis general de la amplificación de todas las muestras con los marcadores de los locus Bem23 y Ms145. Se demuestra el porcentaje de individuos que amplificaron la combinación 220/220 correspondiente a la presencia de la especie MEAM1. Se puede observar que las dos arvenses analizadas mostraron los menores porcentajes de presencia de la especie MEAM1, lo cual puede estar asociado a que estas plantas se recolectaron en ecosistemas no productivos, menos perturbados por las producciones de hortalizas y por altos niveles de incidencia de la especie MEAM1. Esta puede ser una evidencia de que las arvenses que se encuentran en ecosistemas no productivos son nichos de contacto genético y ecológico entre especies nativas e invasivas. Resultados similares se informaron en Isla Reunión para el biotipo B invasivo y el Ms nativo (11). Por otra parte, tanto en calabaza como tomate, el 100% de los individuos analizados correspondieron con la especie MEAM1 y esto confirma los resultados previamente obtenidos (6, 7).
Los ADN positivos para la especie MEAM1, seleccionada a partir del análisis de la combinación de los locus Ms145/Bem23, amplificaron un único fragmento de, aproximadamente, 220pb con el marcador Bem6, correspondiente al alelo 220 del locus Bem6. Estos resultados coincidieron con lo descrito por McKenzie et al. (12) para la especie MEAM 1 o biotipo B.
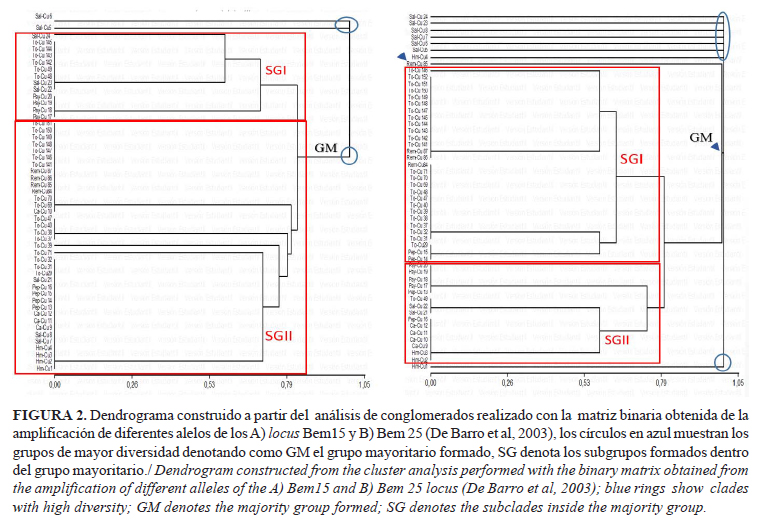
En cada uno de los casos, se obtuvo polimorfismo para la combinación de Bem15 y Bem25 con alelos de tallas desde 100-250pb. El análisis de conglomerados (Fig. 2), para ambos marcadores, demostró que la variabilidad fue, aproximadamente, del 1%.
En ambos locis, sobre la base de 1% de divergencia, se separaron del agrupamiento mayoritario (GM) dos individuos colectados de Salvia officinalis L., en el locus Bem25 se agruparon independientes la mayoría de los aislados de S. officinalis y dos individuos colectados en Solanum americanum Mill. Los grupos mayoritarios formados (GM) se subdividieron para formar dos subgrupos (SGI y SGII), donde en el locus Bem15 los individuos colectados en los diferentes hospedantes se agruparon de forma heterogénea con una variabilidad menor del 1%; sin embargo, en el locus Bem25 el subgrupo SGI incluyó a todos los individuos procedentes de tomate y remolacha, mientras que el SGII incluyó los individuos procedentes del resto de los hospedantes. Estos resultados indican que existe aún una baja variabilidad de la especie MEAM1 en el país, tal como fue informado para el resto de las Américas (13).
Es posible que los bajos porcentajes de divergencias entre los subgrupos estén asociados a los disturbios y la heterogeneidad de los ecosistemas muestreados y que constituya una evidencia de que las interacciones ocurridas y la inmigración de las especies no son aún suficientemente fuertes para la ocurrencia de flujo de genes entre las poblaciones y subpoblaciones genéticas presentes, similar a los descrito por varios autores, debido, posiblemente, a la reciente introducción en este hemisferio (4, 13).
Las secuencias de ADN permitieron confirmar los resultados del estudio con los marcadores microsatélites. Se obtuvieron cuatro secuencias de los fragmentos amplificados de la mtCO1. Dos generadas por individuos colectados de Salvia (S. officinalis, Sal-Cu92) y calabaza (C. pepo; Ca-Cu15) y fueron 99% idénticas a secuencias de la especie MEAM1 de B. tabaci e identificada en otros países de este hemisferio como especie introducida; tal ha sido el caso de Estados Unidos (Genebank Acc. No. HQ877599) y Argentina (Genebank Acc. No. KM100359).
Las dos secuencias restantes se obtuvieron de plantas de Salvia (S. officinalis) con el patrón no MEAM1 (aislados Sal-Cu98, aislados Sal-Cu12) y que correspondieron, con 98% de identidad, con la especie del Nuevo Mundo (NW), informada previamente en Cuba (5) como biotipo nativo (Genebank Acc No. FN821787); estos aislados mostraron identidad con secuencias de otros países del hemisferio como Chile y Honduras (Genebank Acc. No. AY057132, AF34770).
Los resultados se corresponden con los planteado por otros autores (5, 6, 7) sobre la presencia en Cuba de la especies MEAM1 y NW de B. tabaci y constituyen una evidencia de las relaciones simpátricas que pueden estar ocurriendo, inclusive en un mismo hospedante. Ejemplo de esto es el resultado en S. officinalis, sobre la cual se determinó la presencia de más de una especie (MEAM1, NW).
Con estos resultados se confirma que la especie MEAM1 posee habilidad para utilizar múltiples hospedantes y, en algunos casos, compartido con la especie NW; tal como se informó para B. tabaci por otros autores (4, 5, 6, 7, 8). Estos estudios deben continuar en el futuro, a fin de conocer la influencia de los hospedantes en las poblaciones de B. tabaci en el país y poder correlacionarla con el tipo de ecosistemas.
El biotipo Q, endémico del Mediterráneo y ampliamente distribuido por varios países (12, 14, 15), no se encontró dentro de las poblaciones cubanas de B. tabaci. La coexistencia de especies introducidas (MEAM1) y nativa (NW) de Bemisia, tanto en ecosistemas agrícolas como en no productivos, ejercen una fuerza importante en la distribución y aparición de begomovirus en el país. El monitoreo continuo de la distribución de Bemisia tabaci y la dilucidación de las características de las especies presentes son aspectos de crucial importancia para el manejo de esta plaga y para obtener un efectivo control, por la información que ofrece sobre la relación con los hospedantes, la resistencia a insectidas, la asociación con especies de bacterias simbiontes y el flujo de genes en su estructura genética (16).
AGRADECIMIENTOS
Los autores agradecen a la M. Sc. Iris Palenzuela por la identificación de las arvenses colectadas. El trabajo fue financiado por el proyecto ¨Estudio y caracterización de la variabilidad genética de plagas emergentes en los ecosistemas agrícolas¨ del Convenio integral de colaboración Cuba-Venezuela.
REFERENCIAS
1. De Barro PJ, Rueman JWH, Frohlich DR. Bemisia argentifolii is a race of B. tabaci (Hemiptera: Aleyrodidae): the molecular genetic differentiation of B.tabaci populations around the world. Bull Entomol Res. 2005;95:193-203.
2. De Barro PJ, Scott KD, Graham GC, Lange CL, Schutze MK. Isolation and characterization of microsatellite loci in Bemisia tabaci. Mol Ecol. 2003,Notes3: 40-43.
3. Dinsdale A, Cook L, Riginos C, Buckley YM, De Barro PJ. Refined Global Analysis of Bemisia tabaci (Hemiptera: Sternorrhyncha: Aleyrodoidea: Aleyrodidae) Mitochondrial Cytochrome Oxidase 1 to Identify Species Level Genetic Boundaries Ann Entomol Soc Am. 2010;103(2):196-208.
4. De Barro PJ, Liu SS, Boykin LM, Dinsdale AB. Bemisia tabaci: a Statement of Species Status. Annu Rev Entomol. 2011;56:1-19.
5. Vázquez LL. Sistema de diagnóstico, inventario y plantas hospederas de mosca blanca en Cuba. Tesis presentada en opción al grado científico de Doctor en Ciencias Agrícolas. (1995). Instituto de Investigaciones de Sanidad Vegetal. C. Habana, Cuba. 121pp.
6. Muñiz Y, Martínez Y, Martínez MA, Fonseca D, Granier M, Peterschmitt M. Caracterización molecular de poblaciones cubanas de Bemisia tabaci (Gennadius). Rev Protección Veg. 2006;21(3):163-169.
7. Muñiz Y, Granier M, Caruth C, Umaharan P, Marchal C, et al. Extensive Settlement of the Invasive MEAM1 Population of Bemisia tabaci Hemiptera: Aleyrodidae) in the Caribbean and Rare Detection of Indigenous Populations. Environ Entomol, 2011;40(5):989-998.
8. Delatte H, Reynaud B, Granier M, Thornary L, Lett JM, e al. A new silverleaf-inducing biotype Ms of Bemisia tabaci (Hemiptera:Aleyrodidae) indigenous to the islands of the south-west Indian Ocean. Bull Entomol. Res. 2005;95:29-35.
9. Di Rienzo JA, Casanoves F, Balzarini MG, Gonzalez L, Tablada M, Robledo CW. InfoStat versión 2010. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina.
10.Frohlich DR, Torres-Jerez ID, Bedford PG, Markham, Brown JK. A phylogeographical analysis of the Bemisia tabaci species complex based on mitochondrial DNA markers. Mol Ecol. 1999;8:168-1691.
11.Delatte H, David P, Granier M, Lett JM, Goldbach R, et al. Microsatellites reveal extensive geographical, ecological and genetic contacts between invasive and indigenous whitefly biotypes in an insular environment. Genet Res, Camb. 2006;87:109-124.
12.McKenzie CL, Bethke JA, Byrne FJ, Chamberlin JR, Dennehy TJ, Dickey AM, et al. Distribution of Bemisia tabaci (Hemiptera: Aleyrodidae) Biotypes in North America after the Q Invasion. J Econ Entomol. 2012;105(3):753-766.
13. Calvert L, Villarreal N, Frohlich D. Using Molecular Techniques to. Analyse Whitefly Species and Biotypes in Latin America. En: Biotypes of Bemisia tabaci. 2000;CHAPTER 3.13;251-262.
14.Esterhuizen LL, Mabasa KG, van Heerden SW, Czosnek H, Brown J K, et al. Genetic identification of members of the Bemisia tabaci cryptic species complex from South Africa reveals native and introduced haplotypes. J Appl Entomol. 2013;137:122-135.
15.Dong C, Fang HW, Zhang YJ, Brown JK. Change in the Biotype Composition of Bemisia tabaci in Shandong Province of China from 2005 to 2008. Environ Entomol. 2010;39(3):1028-1036.
16.Henria H, Terraza G, Gnankinéb O, Fleurya F, Laurence M. Molecular characterization of genetic diversity within the Africa/Middle East/Asia Minor and Sub-Saharan African groups of the Bemisia tabaci species complex. International Journal of Pest Management 2013;59 (4):329-338.
17.Bing XL, Yanga J, Zchori-Feinb E, Wang XW, Liu SS. Characterization of a Newly Discovered Symbiont of the Whitefly Bemisia tabaci (Hemiptera: Aleyrodidae). Appl Environ Microbiol. 2013;79(2):569-575.
Recibido: 12-10-2015.
Aceptado: 1-8-2016.
*Autor para correspondencia: Yamila Martínez-Zubiaur. Correo electrónico: yamila@censa.edu.cu


{kind=link}
{kind=link}
{kind=link}