










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión On-line ISSN 2224-4697
Rev. Protección Veg. vol.31 no.3 La Habana sep.-dic. 2016
ARTÍCULO ORIGINAL
Coexistencia de Empoasca spp. (Cicadellidae: Typhlocybinae) y tisanópteros en Phaseolus vulgaris L.
Coexistence of Empoasca kraemeri Ross and Moore (Homoptera: Cicadellidae) and Thrips palmi Karny (Thysanoptera: Thripidae) in Phaseolus vulgaris L.
Ileana Miranda Cabrera, Marbely del Toro Benitez, Adayakni Sánchez Castro, Susana Ramírez González, Heyker Lellani Baños Díaz, Moraima Suris Campos, Miriam Fernández Argudín
Grupo Plagas Agrícolas, Dirección de Sanidad Vegetal, Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, Mayabeque, CP 32 700, Cuba
RESUMEN
Generalmente, las densidades de los insectos fitófagos en el campo descienden después de intensas lluvias. Con el objetivo de evaluar este supuesto y demostrar la coexistencia de fitófagos en el cultivo del frijol (Phaseolus vulgaris L.), se monitorearon los insectos vectores presentes en una subpacerla de la variedad BAT-306 en la campaña diciembre 2015-marzo 2016. Las fechas de muestreo se fijaron posteriores a un día con precipitaciones. Se empleó el diseño de cinco cuadrantes fijos. A partir de los 32 días posteriores a la siembra, se monitoreó la densidad poblacional de saltahojas, trips y moscas blancas en cinco plantas tomadas al azar en cada cuadrante. Mediante la ley de potencia de Taylor, se determinó el patrón de disposición espacial de los insectos de mayor incidencia. Se graficó la dispersión por muestreo. Se emplearon modelos matemáticos para evaluar la existencia de competencia interespecífica y se realizó un análisis de componentes principales para determinar la relación de las poblaciones con las condiciones meteorológicas. La densidad promedio de moscas blancas fue inferior a 0,28 individuos por muestreo. Los especímenes de trips y saltahojas dominantes se identificaron como Thrips palmi Karny y Empoasca kraemeri Ross y Moore. Se demostró que estas poblaciones coexisten a partir de la fase reproductiva del cultivo, con un patrón agregado y tasa intrínseca de incremento (rm) de 0,11 para las empoascas y 0,09 para trips. Aun cuando las precipitaciones acumuladas tienen una relación inversa con el nivel poblacional, los valores superiores a los 60 milímetros no lograron densidad cero. La velocidad del viento, el déficit de saturación y la intensidad solar se relacionaron de manera directa con las densidades de los insectos vectores.
Palabras clave: Thrips palmi, Empoasca kraemeri, tasa intrínseca de incremento, disposición espacial, competencia interespecífica.
ABSTRACT
Generally, the densities of phytophagous insects decrease in the field after an intense rain. For evaluating this assumption and demonstrating the coexistence of pests in the bean crop, the insect pests present in the common bean variety BAT 306 were sampled from December 2015 to March 2016. The samples were always taken the day after a rainy day. A five fixed quadrant design was used. Population density was monitored from the day 32 after planting by counting the leafhoppers, thrips, and whiteflies on five random plants in each quadrant. Taylor’s power law was used to determine the spatial pattern distribution of the insects with the highest incidence; population dispersion per sampling was graphed. Inter-specific competition was evaluated using mathematical models, and an analysis of principal components was made to determine the relationships between the populations and some climatic factors. The average density of whiteflies was less than 0.28 individuals by sample. The leafhopper was identified as Empoasca kraemeri and the thrips as Thrips palmi. These two species populations were shown to coexist with an aggregate pattern distribution and an intrinsic rate (rm) of 0.11 and 0.09, respectively, once the crop reached the reproductive stage. Despite the inverse relationship between the accumulated precipitations and the population levels, rainfall values over 60 millimeters did not achieve density zero. Wind speed, deficit of saturation, and sunshine were directly related to the densities of these vector insects.
Key words: Thrips palmi, Empoasca kraemeri, intrinsic increment rate, spatial pattern distribution, inter-specific competence.
INTRODUCCIÓN
El frijol común (Phaseolus vulgaris L.) es un eslabón dentro de la seguridad alimentaria, pues constituye un alimento clave en la canasta básica de la población cubana para suplir las necesidades proteicas (1). En Cuba se incrementa la producción de este rublo al contarse con 129 911 mil hectáreas sembradas (2). Las Cooperativas de Crédito y Servicios (CSS) y los agricultores incrementaron las tierras disponibles para granos y hortalizas (2); el país tiene, entre los lineamientos de la política económica y social, favorecer que la ciencia esté dirigida al incremento de estas producciones (3). Estudiar el comportamiento de los fitófagos presentes, en cada ecosistema en particular, es un inicio dentro de todo el trabajo que se debe acarrear para lograr comprender las dinámicas poblacionales y contribuir a disminuir los fitófagos en este cultivo.
A pesar de que en el frijol se informan diferentes fitófagos de interés (4, 5), se destacan los saltahojas y los trips (6,7) como capaces de producir severos daños por su alimentación directa o como vectores de enfermedades. Además, inciden en una parte estructural de la planta, como es la hoja, con un comportamiento aparentemente similar por el hecho de ser móviles, de pequeño tamaño. Por tanto, se precisa conocer qué tipo de relación se establece entre estas especies en cuanto a su coexistencia.
Conocer cómo inciden ambas poblaciones en una determinada región permitirá establecer medidas de control efectivas (8). Además, es importante identificar la posible coexistencia de las especies en las diferentes condiciones climáticas y características de los agroecosistemas, ya que el cambio climático incide en la proliferación de algunos insectos vectores, cambiando su abundancia relativa y, por consiguiente, las relaciones interespecíficas (9).
Por otra parte, aunque conocemos que son muchos los factores que intervienen en la regulación de las poblaciones (10), algunos autores aseguran que las altas precipitaciones disminuyen marcadamente la densidad de los insectos (11). Si esta disminución es tal que la población del fitófago no causa daños en el cultivo, es algo que hay que probar en las condiciones que se estudien.
Dado que los insectos vectores coinciden en el cultivo y se desconocen las relaciones interespecíficas, el presente estudio tuvo como objetivos: I) conocer la incidencia de insectos vectores en una subparcela de frijol común, II) determinar la coexistencia de los fitófagos que resultaran más abundantes y III) evaluar el decremento de las poblaciones de insectos en un campo de frijol variedad BAT 306, después de intensas lluvias.
MATERIALES Y MÉTODOS
La toma de datos experimentales se realizó en la finca “La Ciruela” (23,09° latitud sur y -82,12° longitud este), perteneciente a la CSS “Manuel Fajardo” Municipio San José de las Lajas, provincia Mayabeque, Cuba. Se estableció una subparcela de 4x16 m2 seleccionada al azar, en una hectárea de frijol variedad BAT-306. La distancia de siembra fue de 0,70 m entre hileras y 0,05 m entre plantas. Se utilizó nutrición orgánica y química, mediante compost, humus de lombriz, otros nutrientes orgánicos y fertilizantes químicos (fórmula completa NPK). El riego fue por aspersión.
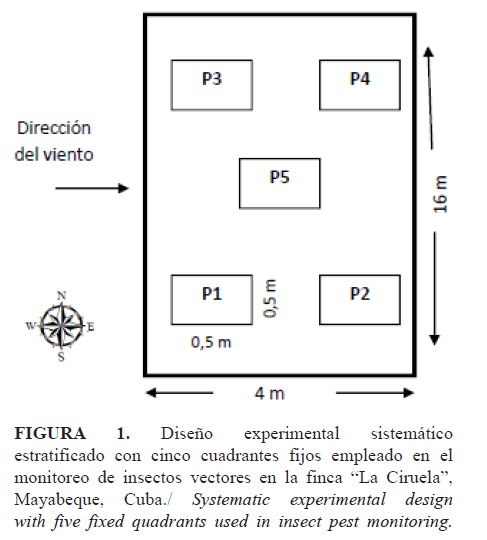
A partir de 32 días después de la siembra, se efectuaron siete muestreos; la fecha de muestra se ubicó un día después de la ocurrencia de lluvias en el municipio San José de las lajas, donde se encuentra la finca. Se contaron insectos vectores (saltahojas, thrips y mosca blanca) presentes en cinco plantas tomadas al azar, alrededor de cinco puntos marcados según el diseño de los cinco cuadrantes (Fig. 1).
En cada muestreo se recolectaron especímenes de los insectos vectores y se llevaron al Laboratorio de Entomología de la Dirección Sanidad Vegetal del Centro Nacional de Sanidad Agropecuaria (CENSA), para su identificación con la ayuda del microscopio estereoscopio marca Stemi SVZeiss. En el caso de los saltahojas, la identificación se realizó con la ayuda de la clave de Zayas (12) y se consideraron solo las ninfas. Los tisanópteros se analizaron como un complejo de larvas que se identificaron al realizar el montaje de los adultos, según la técnica convencional de Mound y Marullo (13) y con la ayuda del microscopio óptico Axioskop 40, Zeiss hasta 400 X; se utilizaron las claves de Mound y Marullo (13) y Mound y Kibby (14).
Se cuantificó el complejo de larvas; se estudió la fluctuación poblacional en correspondencia con la fenología del cultivo y se describió la distribución por puntos.
Se determinó el patrón de disposición espacial a partir de la ley de potencia de Taylor (15). Mediante modelos logísticos de crecimiento (15), se calculó la tasa intrínseca de incremento de los saltahojas y trips. El sistema de competencia interespecífica (9) y el modelo de cuantificación de la competencia (16) se emplearon para demostrar la coexistencia de tisanópteros y saltahojas.
De la estación meteorológica de Tapaste, ubicada a menos de 1 Km de la finca, se tomaron los datos de las variables: temperatura del aire (máxima, media y mínima), temperatura diurna y nocturna, déficit de saturación medio y máximo, lluvias, humedad relativa, evaporación, evapotranspiración, velocidad del viento máxima y media, nubosidad, brillo solar y radiación global. Se calculó el promedio de 10 días anteriores al muestreo de cada variable, excepto las precipitaciones que se tomaron acumuladas. Los datos meteorológicos se relacionaron con la densidad promedio de los fitófagos mediante el análisis de componentes principales empleando el paquete estadístico InfoStat 2016 (17).
RESULTADOS Y DISCUSIÓN
De los insectos evaluados se encontraron, como especies predominantes entre los saltahojas, Empoasca kraemeri Ross y Moore y el tisanóptero Thrips palmi Kanry; lo anterior coincide con lo informado por otros autores que mencionan a estos insectos como plagas claves del cultivo y vectores de enfermedades virales (7, 18). Aunque se deben hacer otros estudios encaminados a identificar un número mayor de especímenes con el uso de la genitalia de los machos y técnicas moleculares, dado que en el frijol se presenta un complejo de especies del género Empoasca de las que no se han encontrado características morfológicas confiables para su separación (19).
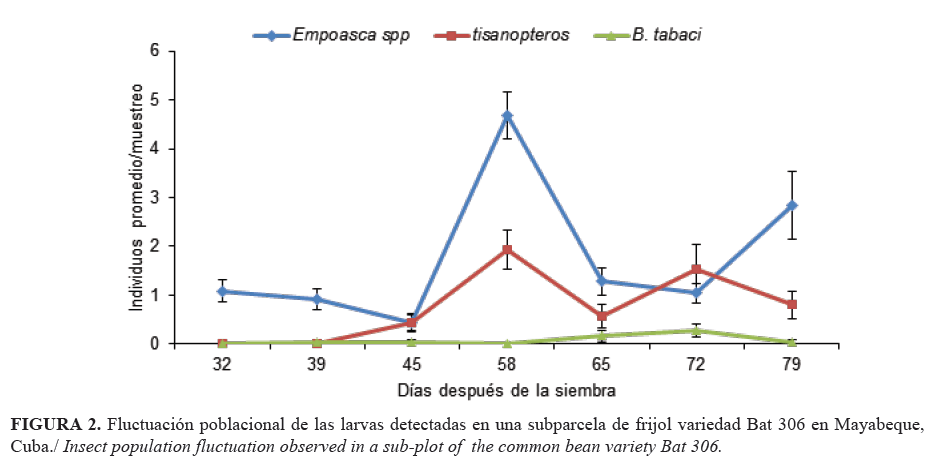
La mosca blanca presentó baja incidencia, inferior a 0,28 individuos promedios por muestreo (Fig. 2). Esto puede deberse a que después del cuarto estado ninfal la acción de los vientos y las lluvias es letal para esta plaga (20). Sin embargo, también se recomienda un manejo de este vector, puesto que no necesita de grandes poblaciones para ocasionar un grado alto de daño; un adulto de mosca blanca puede alimentarse de varias plantas y trasmitir virus (5).
Se presentaron síntomas de arrugamiento, amarilleamiento, moteado, acucharamiento, clorosis y mosaico; en algunas plantas coinciden dos o más síntomas. Esto demuestra que, a pesar de las intensas lluvias, las poblaciones de vectores trasmisores se mantienen en el frijol con densidades que pueden llegar a constituir daños para el cultivo.
Empoasca spp. estuvo desde el primer muestreo (Fig. 2), ya que este se presenta en todo el ciclo de vida del cultivo y se incrementa después de la aparición de las hojas trifoliadas (21). Los tisanópteros se encontraron a partir de los 45 días después de la siembra, con incremento marcado hacia la fase de floración (V4, 58 dds) (Fig. 2), lo que coincide con lo planteado por Murguido (6), quien refiere que ambos insectos se incrementan a los 40 días después de la siembra (dds) y alcanzan poblaciones superiores a cinco individuos por planta.
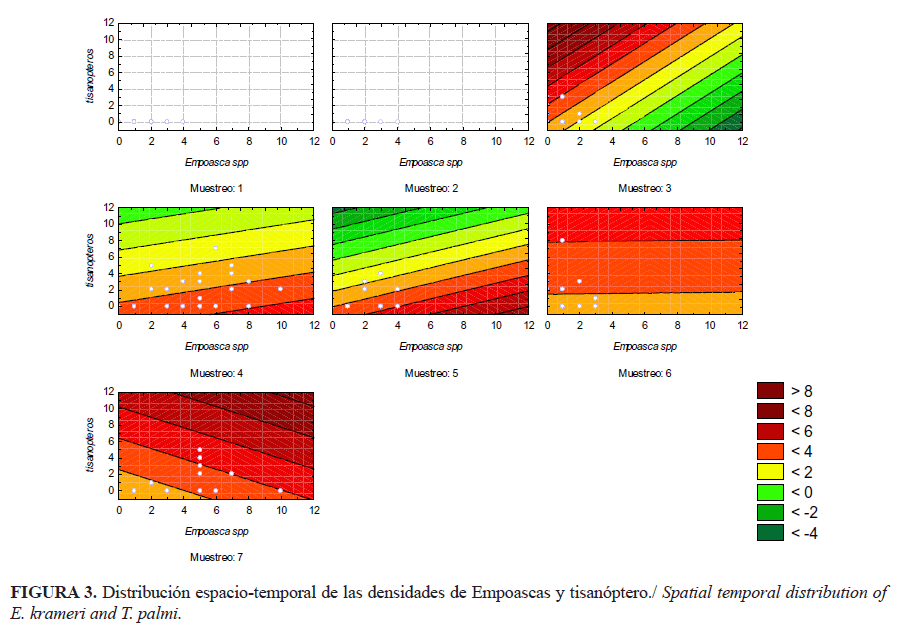
En los diferentes muestreos, las densidades poblacionales por puntos estuvieron muy similares; existió mayor varianza de la densidad por planta en los muestreos cuatro y siete, que coinciden con los momentos de mayor densidad poblacional de ambos insectos (Fig. 3). Los grupos poblacionales presentaron un patrón agregado con índice b de Taylor superior a 1 (Tabla 1). En el caso de los thrips en mayores extensiones de tierra, también hay una tendencia a la agregación, debido a que, en la mayoría de los cultivos, estos insectos prefieren las partes verdes de la planta y las flores (21). En concordancia con otros autores, el índice b es un factor inherente a la especie (22) y los insectos presentan un patrón aleatorio solo cuando están en densidades muy bajas (23).

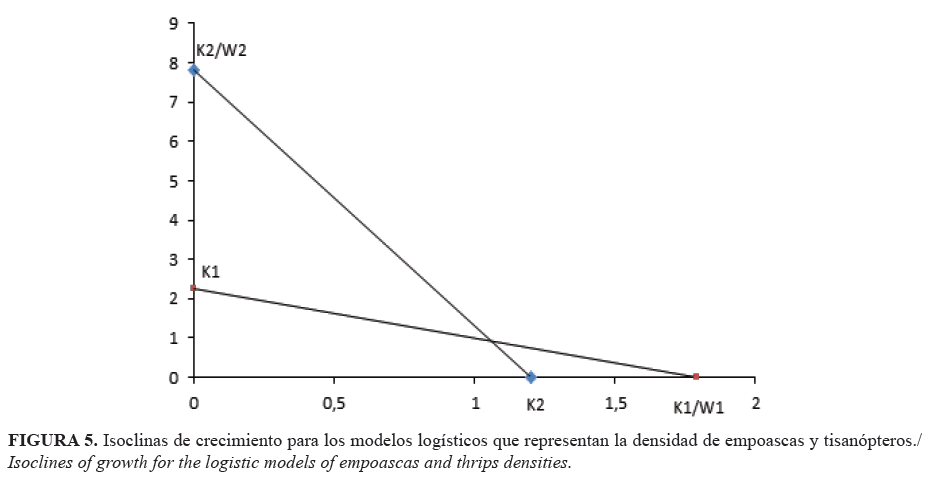
El complejo de saltahojas se multiplicó con mayor rapidez que los thrips, con tasas intrínsecas de incremento de 0,11 y 0,09, respectivamente (Fig. 4). La especie estudiada y las características del cultivo hospedante influyen en los paramétros de crecimiento de las poblaciones (23). La tasa intrínseca de los tisanópteros puede ir de valores inferiores a 0,01 en pobres hospedantes, a valores mayores de 0,20 en buenos hospedantes (24). Por lo que es recomendable estudiar este paramétro en la búsqueda de variedades resistentes. La gráfica puede simular la existencia de un desplazamiento competitivo (Fig. 4.); pero, al graficar las isoclinas (Fig. 5), se demostró la coexistencia de ambos grupos de especies (8).
La coexistencia de Empoasca spp. y los tisanópteros se constató al obtener el valor -0,09 para el coeficiente de competencia interespecífica β, con intervalo negativo para el 95% de confianza (-0,06;-0,12) en el modelo de exclusión competitiva (16):
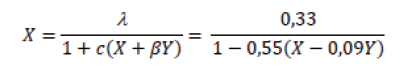
donde X representó densidad de trips, Y densidad de Empoasca spp., λ, c y β parámetros estimados por el modelo, donde λ representa la supervivencia de trips en presencia de Empoasca spp., c un paramétro que afecta la respuesta competitiva, pero no tiene interpretación biológica, y β el coeficiente de competencia.
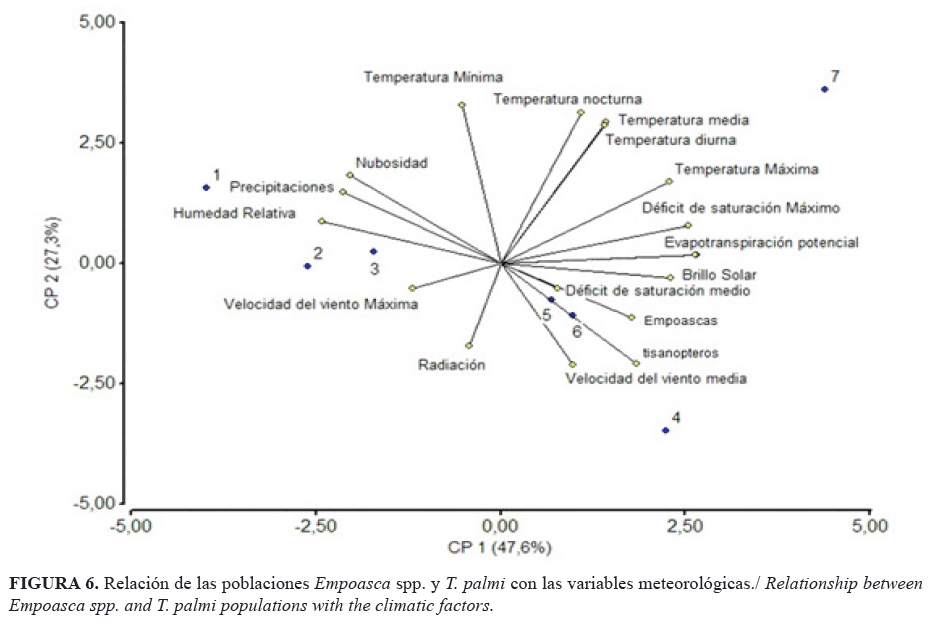
Con dos componentes principales se representó el 74,9% de la variabilidad total de los factores. Las precipitaciones acumuladas fueron superiores a los 61 mm y, de conjunto con la humedad relativa y la nubosidad, presentaron una relación inversa con las poblaciones (Fig. 5); lo que coincide con otros autores que plantean que la densidad de los saltahojas disminuye con las lluvias (25). No obstante, el aumento de las precipitaciones y la alta humedad no redujeron a cero la incidencia de los insectos, dado que estos se incrementan rápidamente y alcanzan valores elevados en dependencia de la fenología del cultivo (26).
La velocidad del viento presentó una relación directa con la densidad poblacional de Empoasca spp. y trips, ya que, al igual que con otros insectos, el viento favorece el movimiento de las poblaciones. En estos casos se recomienda crear barreras físicas que impidan el movimiento poblacional hacia el cultivo (26).
Es de destacar que el déficit de saturación influyó de manera directa en la densidad de los insectos (Fig. 6). No es común que la variabilidad de este factor se tenga en cuenta en los análisis de regulación de poblaciones, pero las plantas tienen que transpirar agua para trasportar nutrientes, para refrigerarse y para regular su crecimiento. La transpiración depende del déficit de saturación entre las estomas y el aire. Cuando los déficits de saturación son muy altos, o muy bajos, influyen en la fisiología del cultivo y en su desarrollo (19).
Podemos concluir que, aun en pequeñas parcelas, el frijol común es atacado por insectos vectores que coexisten en el cultivo, presentan un patrón de disposición agregado y una fluctuación que depende de la fenología del cultivo y la incidencia de las condiciones meteorológicas. La variabilidad de la humedad relativa, las precipitaciones, la nubosidad, la velocidad del viento y el déficit de saturación se deben tener en cuenta en futuros estudios encaminados a establecer modelos de simulación de la dinámica de los fitófagos vectores.
AGRADECIMIENTOS
Los autores desean expresar su agradecimiento al agricultor Ruperto Díaz de la CSS “Manuel Fajardo”, por las facilidades ofrecidas en su finca para realizar el monitoreo de las poblaciones.
REFERENCIAS
1. Ramos Y, Gómez J, Espinosa R, Días F, CrespoII A, Machado R. Etología de los crisomélidos (Coleoptera: Chrysomelidae) asociados a tres variedades de frijol común (Phaseolus vulgaris L.) en época intermedia. Rev Protección Veg. 2015;30(3):165-170.
2. Oficina Nacional de Estadísticas (ONE). Anuario Estadístico de Cuba 2014. Capítulo 9. Agricultura, ganadería, silvicultura y pesca. Edición 2015; 33p.
3. Congreso PCC. Lineamientos de la política económica y social del partido y la revolución. 2011; 38p.
4. Subramanya K, Zitter TA. Plant virus and viroid diseases in the Tropics. Vol 2. Epidemiology and Management. Editorial Springer Science+Business Media. 2014; 489p. ISBN 978-94-007-7819-1.
5. Zamora L. Detección e identificación de Ca. Phytoplasma ssp. y begomovirus en el cultivo Phaseolus vulgaris L. en Cuba. Rev Protección Veg. 2014;29(3):236.
6. Murguido CA, Vázquez L, Eliozondo AI, Neyra M, Velázquez Y, Pupo E, et al. Manejo integrado de plagas de insectos en el cultivo del frijol. Fitosanidad. 2002;6(3):29-41.
7. Suris M. Especies de trips (Thysanoptera) presentes sobre el cultivo del frijol en localidades de la provincia la Habana. Rev Protección Veg. 2001;16(2-3):132-134.
8. Treviño C, Rosas R. El frijol común: factores que merman su producción. Revista de Divulgación Científica y tecnológica de la Universidad Veracruzana. 2013;26(1):1-6.
9. Suris M, Pérez JC, Miranda I. Competencia inter-específica entre Heliothis virescens (F.) y Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) en el cultivo del garbanzo (Cicer arietinum L.). Rev Protección Veg. 2013;28(3):171-177.
10. Barclay HJ, Matlock R, Gilchrist S, Suckling D, Reyes J, et al. A conceptual model for assessing the minimum size area for an area-wide Integrated Pest Management Program. International Journal of Agronomy. 2011; ID 409328:1-12.
11. Vázquez L. Cambio climático, incidencia de plagas y prácticas agroecológicas resilientes. En: Innovación agroecológica, adaptación y mitigación del cambio climático. Instituto Nacional de Ciencias Agrícolas (INCA). La Habana. 2011; 242p. ISBN 978-959-7023-53-1.
12. Zayas F. Entomofauna cubana. Tomo VII. Editorial Científico-Técnica. La Habana. 1988; 259p.
13. Mound LA, Marullo R. The Thrips of Central and South America: An introduction (Insecta: Thysanoptera). Mem Entomol Internat. 1996;6:1-487.
14. Mound LA, Kibby G. Thysanoptera and Identification Guide. Second Edition. CAB International; 1998. 67p.
15. Miranda I. Estadística Aplicada a la Sanidad Vegetal. Editorial Félix Varela 2011; 176p. ISBN 978-959-7125-44-0.
16. Paini D, Funderburk J, Reitz S. Competitive exclusion of a worldwide invasive pest by a native. Quantifying competition between two phytophagous insects on two host plant species. Journal of Animal Ecology. Disponible en: http://www.onlinelibrary.wiley.com/doi/10.1111/j.1365-2656.2007.01324.x/abstract.
17. Di Rienzo JA, Casanoves F, Balzarini MG, Gonzalez L, Tablada M, Robledo CW. InfoStat versión 2016. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. Disponible en: http://www.infostat.com.ar.
18. Castellanos L, Astengo JA, Yero Y, Herrera N, Fernández R. Plagas y enfermedades en 13 variedades de frijol en una localidad de la provincia Sancti Spíritus. Centro Agrícola. 2011;38(1):91-92.
19. Segnini S, Montagne A. Biología y ecología poblacional de Empoasca kraemeri Ross y Moore (Homoptera: Cicadellidae) en caraota (Phaseolus vulgaris L). IV. Disposición espacial de E. kraemeri dentro de campos cultivados con caraota. Agronomía Tropical. 1986;36(4-6):47-64.
20. Naranjo S, Ellsworth P. Mortality dynamics and population regulation in Bemisia tabaci. Entomologia Experimentalis et Applicata. 2005;116(2):93-118.
21. Maya-Hernández V, Vera-Graziano J, Garza-García R. Parámetros poblacionales de Empoasca kraemeri Ross & Moore (Homoptera: Cicadellidae) en genotipos de frijol. Agrociencia. 2000;34:603-610.
22. Niassy S, Ekesi S, Maniania N, Orindi B, Moritz G, et al. Active aggregation among sexes in vean flower thrips (Megalurothrips sjostedti) on cowpea (Vigna unguiculata L). Entomologia Experimentalis et Applicata. 2016;158(1):17-24.
23. Rabinovich J. Introducción a la ecología de poblaciones animales. Méx., Continental. 1980; 313 p.
24. Liang-De T, Kai-Li Y, Bu-Li F, Jian-Hui W, Kui L, Yong-Yue L. The life table parameters of Megalurothrips usitatus (Thysanoptera: Thripidae) on four leguminous crops. Florida Entomologist. 2015;98(2):620-625.
25. Carvalho DP, Oliveira HN. Population fluctuation of Empoasca sp. (Hemiptera: Cicadellidae) in a physic nut crop in Mato Grosso do Sul. Acta Biol Colomb. 2016;21(1):81-85.
26. Vázquez L. Manejo plagas en la agricultura ecológica. Boletín Fitosanitario. 2010;15(1):1-120.
Recibido: 19/05/2016
Aceptado: 7/11/2016
Ileana Miranda Cabrera, Grupo Plagas Agrícolas, Dirección de Sanidad Vegetal, Centro Nacional de Sanidad Agropecuaria (CENSA). Email: ileanam@censa.edu.cu


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}