










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión On-line ISSN 2224-4697
Rev. Protección Veg. vol.31 no.3 La Habana sep.-dic. 2016
ARTÍCULO ORIGINAL
Identificación y caracterización de aislamientos de Fusarium spp. presentes en garbanzo (Cicer arietinum L.) en Cuba
Identification and characterization of Fusarium spp. isolates present in chickpea (Cicer arietinum L.) in Cuba
Yanisia Duarte Leal,I Anayza Echevarría Hernández,II Benedicto Martínez Coca,I
ICentro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, CP 32 700, Mayabeque, Cuba.
IIUnidad Científico Tecnológica de Base Los Palacios (UCTB-LP), Pinar del Río; Instituto Nacional de Ciencias Agrícola (INCA), Cuba.
RESUMEN
El objetivo de este trabajo fue identificar y caracterizar, cultural y morfológicamente, dieciséis aislamientos de Fusarium spp., presentes en semillas y plantas de garbanzo (Cicer arietinum L.) procedentes de zonas de Cuba. Las variables culturales evaluadas fueron: color de la colonia en el anverso y reverso, textura superficial, forma de los bordes, presencia de anillos en el anverso, crecimiento lineal; entre las morfológicas aparecen: tamaño, forma y color de las estructuras vegetativas y reproductivas. Para la identificación de los hongos se utilizó una clave taxonómica. Los aislamientos mostraron gran variabilidad en todos los caracteres analizados. Se identificó la presencia de Fusarium proliferatum (Matsushima)Nirenberg, Fusarium nygamai Burgess & Trimboli, Fusarium dlamini Marasas, Nelson & Toussoun, Fusarium phyllophilum Nirenberg & O’Donnell, representados con un aislamiento, mientras que Fusarium solani (Martius) Appel & Wollenweber emend. Snyder & Hansen y Fusarium oxysporum Schlechtendahl emend. Snyder & Hansen, con dos y tres aislamientos, respectivamente, y uno que, de acuerdo a sus características, se mantuvo como Fusarium sp. Hubo variabilidad cultural entre aislados de una misma especie, lo que denota la posible presencia de biotipos.
Palabras clave: fusariosis, marchitez vascular, podredumbre de raíz, necrosis.
ABSTRACT
The aim of this study was the cultural and morphological identification and characterization of sixteen isolates of Fusarium spp. from chickpea (Cicer arietinum L.) seeds and plants. Colony color (surface and reverse) and texture, shape of the colony edges, presence of rings on the colony surface, and the linear growth, were the cultural variables evaluated. Size, shape, and color of the vegetative and reproductive structures were the morphological variables examined. A taxonomic key was used for identifying the fungi. All the isolate characteristics analyzed showed a great variability. It was identified the presence of Fusarium proliferatum (Matsushima) Nirenberg, Fusarium nygamai Burgess & Trimboli, Fusarium dlamini Marasas, Nelson & Toussoun, and Fusarium phyllophilum Nirenberg & O’Donnell, all them represented by one isolate. Two isolates corresponded to Fusarium solani (Martius) Appel & Wollenweber emend. Snyder & Hansen and three to Fusarium oxysporum Schlechtendahl emend. Snyder& Hansen. According to its characteristics, one of the isolates remained as Fusarium sp. Cultural variability was observed among the isolates of a same species, which showed the possible presence of biotypes.
Key words: fusariosis, vascular wilt, root rot, necrosis.
INTRODUCCIÓN
El garbanzo (Cicer arietinum L.) es la segunda leguminosa de grano de importancia en el mundo (1); sin embargo, las producciones en Cuba, hasta los años 90, no fueron de consideración y, en consecuencia, no aparecen registros estadísticos del cultivo. Debido a ello, para satisfacer el consumo interno, fue necesario realizar importaciones provenientes, mayoritariamente, de México, Canadá y España (2).
El incremento de las áreas en el cultivo del garbanzo en Cuba se logró gracias a la introducción de cultivares que responden a las condiciones del país, así como al conocimiento cada vez mayor del cultivo, el aumento de los precios en la comercialización y los bajos costos de producción con relación a otras especies de granos, por lo que constituye un rubro atractivo para su producción (3). Por otro lado, este cultivo es poco exigente al agua y puede cosecharse en condiciones de sequía severa. Esto constituye un valioso aporte en el empeño de Cuba por sustituir importaciones de alimentos muy costosos en el mercado internacional (4).
Las enfermedades representan una de las causas en la disminución de los rendimientos en el cultivo; entre las más importantes en Cuba y el mundo, se encuentran las ocasionadas por hongos del suelo, como son: Fusarium spp., Sclerotium rolfsii Sacc., Rhizoctonia solani Kühn, Macrophomina phaseolina (Tassi) Goid; sin embargo, por los daños que producen, y la frecuencia con que se presentan, Fusarium spp. son los de mayor significación (5).
Las fusariosis aparecen, principalmente, en los suelos arcillosos o con problemas de drenaje, condición que favorece el desarrollo de estas patologías (6). En la India y Pakistán, el 15% de las cosechas se afectan anualmente por Fusarium. Estos hongos también provocan pérdidas en México y España; en la Cuenca del Mediterráneo y California la enfermedad devastó el cultivo por completo en condiciones favorables para su desarrollo (7).
En las fusariosis en el cultivo se integran diferentes síndromes, denominados Marchitez vascular y Amarillez vascular, que son ocasionados por Fusarium oxysporum Schlechtend.: Fr. f. sp. ciceri (Padwik) Matuo & K. Sato (Foc) (8), y el complejo de síntomas designado como “Marchitez y Podredumbre de Raíz” (MPR), que se asocia con infecciones causadas por diversos hongos de suelo, incluyendo F. oxysporum f. sp. ciceri, Fusarium solani (Mart.) Appl. & Wr. f. sp. pisi (Jones) Snyd & Hans.,Fusarium solani f. sp. eumartii (C. Carpenter)W.C. Snyder & H.N. Hans. (=Fusarium eumartii Carpenter), M. phaseolina (1) y Fusarium redolens Wollenw (9). Estos síndromes son consecuencia de infecciones vasculares en la planta infectada (10, 11).
El sídrome MPR provoca un amarillamiento prematuro de las hojas basales, se relaciona con necrosis en la raíz principal y estrangulamiento a nivel del cuello (10). Sin embargo, F. redolens ocasiona amarillamiento foliar acrópeto y necrosis radical (9).
En Cuba, Dueñas et al. (12) caracterizaron especies de Fusarium que afectaron al garbanzo en las antiguas provincias Ciudad Habana y La Habana, identificadas como F. oxysporum f. sp. ciceri y F. solani. No obstante, la información acerca de las especies de Fusarium en este cultivo en el país es aún escasa.
Por la importancia que tiene este cultivo, este trabajo tuvo como objetivos identificar y caracterizar, cultural y morfológicamente, aislamientos de Fusarium spp. procedentes de garbanzo.
MATERIALES Y MÉTODOS
La investigación se efectuó en el Laboratorio de Micología Vegetal perteneciente al grupo de Fitopatología del Centro Nacional de Sanidad Agropecuaria (CENSA), en el municipio San José de las Lajas, provincia Mayabeque, Cuba.
Aislamiento del agente causal
Se muestrearon plantas de diferentes líneas de garbanzo, que presentaron síntomas típicos de marchitez, con necrosis y amarillamiento y que crecían en suelo Hidromórfico Gley Nodular Ferruginoso, en la Unidad Científico Tecnológica de Base Los Palacios (UCTB-LP), provincia Pinar del Río, perteneciente al Instituto Nacional de Ciencias Agrícolas (INCA), Cuba. Las muestras de plantas con diferentes síntomas y otras, aparentemente sanas, se trasladaron en sobres de papel Kraft, previamente esterilizados, al Laboratorio de Micología Vegetal.
Además, se analizaron semillas con afectaciones (deformaciones, reducción del tamaño y/o decoloración) provenientes de la primera obtención de semillas de Siria e INIFAT (Cuba). Estas se lavaron con abundante agua corriente durante 20 min, se desinfectaron con alcohol (70%) por 45 s y se lavaron tres veces en agua destilada estéril; después se trataron con una solución de hipoclorito de sodio (1%), se enjuagaron tres veces en agua destilada estéril y se secaron con papel absorbente estéril. Posteriormente, se colocaron grupos de cuatro semillas en cámaras húmedas (placas Petri de 150 mm de diámetro con papel de filtro esterilizado humedecido). Las semillas se observaron diariamente, hasta la aparición de estructuras fúngicas. Se observaron fragmentos de estas al microscopio óptico Zeiss (200x) para corroborar la presencia de Fusarium.
Por otro lado, a las muestras de raíces necrosadas y/o tallos con estrangulamiento en la base, se les realizaron cortes longitudinales y se seccionaron en fragmentos de 10×20mm asépticamente; se procesaron de igual forma que las semillas.
Los fragmentos de micelio, que se obtuvieron de semillas y de tallos, se sembraron en placas Petri (Ø=90 mm) con medio Papa Dextrosa Agar (PDA) (Biocen) más cloranfenicol (0,01g.L-1) y se incubaron a 28ºC± 2ºC y oscuridad constante. Se observaron los fragmentos de micelios de los aislados obtenidos en un microscopio óptico Zeiss (200x) para confirmar su identidad con el género Fusarium.
Las colonias de los aislados analizados se purificaron mediante siembras en el mismo medio, hasta la obtención de cultivos puros que, posteriormente, se transfirieron a tubos de ensayos con medio PDA (Biocen) en forma de cuña y se incubaron como se describió anteriormente, por diez días. Los aislados se conservaron a 4ºC en el cepario del Laboratorio de Micología Vegetal del CENSA.
Obtención de cultivos monospóricos de los aislamientos
A los tubos contentivos de los aislados, desarrollados durante siete días en cuñas de PDA (Biocen), se les añadieron 10 ml de agua destilada estéril y se homogeneizaron en agitador Vortex por 30 s. Con un asa bacteriológica desinfectada, se tomó una asada de la suspensión y se depositó en el centro de placas (Ø=90mm) que contenían cinco ml de medio de cultivo Agar agua (2%) y se dispersó con una espátula de Drigalsky. Todas las placas se sellaron con papel parafilm y se incubaron a 28ºC±2ºC y oscuridad constante, durante 24 h. Se hicieron tres réplicas por aislamiento.
Después de este periodo, bajo un microscopio óptico Zeiss (200x), se detectaron y señalaron conidios solitarios germinados por el reverso de la placa, con un marcador de punta fina. Con una aguja de siembra flameada se transfirió el fragmento de agar, donde se encontraba un conidio germinado marcado a una placa Petri con medio de cultivo PDA (Biocen) más cloranfenicol (0,01%). Las placas se sellaron con parafilm e incubaron a 28ºC±2ºC durante siete días. Una vez que se lograron los cultivos puros monospóricos, se conservaron a 4ºC en tubos de ensayo con cuñas del propio medio de cultivo en el Laboratorio de Micología Vegetal del CENSA.
Comprobación de los postulados de Koch
El inóculo se preparó bajo condiciones asépticas, a partir de cultivos de los aislamientos monospóricos sembrados en tubos con cuñas de PDA (Biocen) e incubados a 28ºC±2ºC y oscuridad durante siete días. A los tubos con los aislados se les adicionaron 10 ml de agua destilada estéril y se homogeneizaron en agitador Vortex durante un minuto. Las suspensiones obtenidas se ajustaron a la concentración de 106 conidios.ml-1 para cada aislamiento, con la adición de agua destilada estéril; la concentración se determinó en una cámara de Thoma.
Se tomaron fragmentos cuadrados de papel Kraft y se depositaron 100 g de suelo, se doblaron a la mitad y se enrollaron. Se colocaron en frascos de cristal y se esterilizaron a 121ºC durante 30 minutos. Posteriormente, bajo condiciones asépticas, sobre el suelo esterilizado se depositaron, en línea, diez semillas de garbanzo cultivar 5HA, previamente desinfectadas [con alcohol al 70% (45 s) e hipoclorito de sodio al 10% (2 min) y tres enjuagues con agua destilada estéril]. A continuación, con ayuda de una pipeta estéril, se le aplicaron 20 ml de la suspensión de cada aislamiento independientemente, al suelo y a las semillas. El papel, con el suelo y las semillas inoculadas, se enrolló, se envolvió en papel aluminio esterilizado y se depositó verticalmente en un recipiente de cristal que contenía 10 ml de agua destilada estéril en el fondo. El testigo (semillas desinfectadas en suelo estéril con la adición de 20 ml de agua estéril sin la inoculación del patógeno) se procesó de igual forma que los tratamientos con los aislamientos de Fusarium. Los recipientes con los rollos de papel se incubaron a temperatura ambiente (30ºC ±2ºC) en un local en condiciones asépticas.
Los tratamientos se examinaron diariamente, hasta la brotación de las plántulas (aproximadamente siete-diez días). De las plantas afectadas se hicieron aislamientos del agente causal, según se describió anteriormente, y se compararon las características de la colonia (color, forma, textura), así como de las estructuras fúngicas del reaislamiento con las del aislado respectivo que se utilizó como inóculo.
Se evaluaron los síntomas ocasionados por los aislados en las plantas y se seleccionaron los aislamientos más agresivos sobre la base de la cantidad de plantas afectadas.
Caracterización cultural y morfométrica de los aislamientos de Fusarium
La caracterización cultural de los aislamientos de Fusarium spp. se realizó sobre la base de caracteres cualitativos y cuantitativos, expresados por los aislamientos sobre los medios de cultivo PDA (Biocen), Czapek Dox (CzD) (Fluka) y Leonian (13).
Se emplearon placas Petri de 90 mm que contenían 15 ml de cada medio de cultivo independientemente [esterilizados (121ºC, 15 min y pH ajustado a 6)]. Las mismas se inocularon, central e individualmente, con un disco de micelio de 6 mm de diámetro de cada aislamiento de Fusarium, proveniente de la periferia de colonias, crecidas sobre PDA durante 8-10 días. Las placas, tres por aislado (réplicas), se sellaron con papel parafilm e incubaron (28ºC±2ºC y oscuridad constante). Las evaluaciones se realizaron cada 24 h hasta 168 h.
La coloración de la colonia en el anverso y reverso se determinó por observación visual y se documentó mediante fotografías tomadas con una cámara Olympus. También se describieron la textura superficial, forma de los bordes y presencia de anillos en el anverso. Se midió el crecimiento lineal (diámetro promedio de la colonia sobre la base del crecimiento horizontal en dos diagonales perpendiculares, expresado en milímetros por día), con una regla graduada.
A partir del crecimiento lineal, se calculó la velocidad de crecimiento al séptimo día de incubación, empleando la fórmula siguiente:

Donde:
V crec f= Velocidad de crecimiento final al completar el área de la placa (en mm.h-1).
Dc= Diámetro de las colonias (en mm)
t = Tiempo hasta siete días o momento en que la colonia completará el área total de la placa (en h)
El conteo de esporas se realizó a los siete días de crecimiento de las colonias de cada aislamiento en los medios de cultivo. A las placas con los cultivos desarrollados se les adicionaron 15 ml de agua destilada estéril y se homogeneizaron con una espátula de Drigalsky. Las esporas se colectaron en un tubo de ensayo por réplica. Las suspensiones se homogeneizaron en agitador Vortex por 30 s y se depositaron gotas de estas (de cada aislamiento por separado), en una cámara de Neubauer para determinar la concentración de conidios. Se compararon los resultados según la cantidad de conidios por área de la colonia.
Los datos de las velocidades de crecimiento y las concentraciones de esporas de los aislados en cada medio de cultivo se sometieron a un Análisis de Varianza (ANOVA) bifactorial y las medias se compararon según la Dócima de Rangos Múltiples de Duncan para p≤0,05, empleando el paquete estadístico INFOSTAT Profesional ver. 2.1.
Identificación de los aislamientos
Los aislamientos se sembraron en tres medios de cultivo, según la clave referida por Leslie y Summerell (14), para la identificación de aislamientos de Fusarium: hojas de clavel agar [Carnation Leaf-piece Agar (CLA)], PDA y Spezieller Nährstoffarmer Agar (SNA). Se utilizaron placas Petri de 90 mm de diámetro, contentivas de 15 ml de cada medio de cultivo por separado (esterilizados a 121oC por 15 min y pH 6). El medio de cultivo CLA se elaboró al verter en cada placa 15 ml de Agar agua estéril; seguidamente se depositaron fragmentos (seis-siete) de hojas de clavel de 3-5 mm², previamente esterilizadas.
Las placas se inocularon central e individualmente con cada aislamiento de Fusarium; se sellaron eincubaron a 28ºC±2ºC y oscuridad constante durante cinco días. Se utilizaron tres réplicas por cada aislamiento.
Aspectos taxonómicos evaluados en cada medio
En el medio de cultivo CLA, se visualizaron al microscopio óptico Zeiss (400x) los macroconidios, células conidiógenas y microconidios (presencia y disposición). Además, se realizaron preparaciones microscópicas de cada aislamiento para observar la forma y el tamaño de los macroconidios, septos en los microconidios y determinar la disposición de las clamidosporas.
En el medio PDA se hicieron las observaciones para determinar la pigmentación, la forma y la velocidad del crecimiento de las colonias, la formación de esporodoquios y el número de septos en los macroconidios; mientras que en el SNA se observó la presencia de clamidosporas y forma de microconidios y macroconidios.
RESULTADOS Y DISCUSIÓN
Las muestras de plantas con diferentes síntomas presentaron pudrición negro-rojiza en la raíz principal, que abarcó hasta las raíces secundarias, con estrangulamiento a nivel del cuello. La necrosis se extendió hasta la parte superior del tallo y las ramas. Se observó un cambio de color, de verde oscuro a verde pálido en las hojas de la parte superior de las plantas enfermas, con flacidez de arriba hacia abajo y concluyó con la marchitez de la planta. En las plantas con síntomas avanzados, apareció una clorosis amarillo opaco en la parte inferior de la planta. Este síntoma se detectó en una o más ramas, avanzó de abajo hacia arriba hasta quedar la planta completamente amarilla y ocasionarle la muerte. Estos síntomas se describen por diferentes autores de forma independiente (8, 10) y pudieran ser asociados con diferentes agentes causales.
De estas muestras se obtuvieron 16 aislamientos monospóricos pertenecientes al género Fusarium (Tabla 1).
De semillas aparentemente sanas y de otras afectadas con deformaciones, reducción del tamaño y/o decoloración, se aislaron hongos pertenecientes a los géneros Fusarium spp. y Aspergillus spp.; sin embargo, más del 70% de los aislamientos correspondieron al primer género. Esto corrobora lo informado por Aguaysol et al. (15) y García et al. (16): el agente causal de la marchitez en este cultivo puede ser diseminado por semilla. De ahí, la importancia de obtener semillas de calidad y de realizar tratamientos de desinfección de las mismas.
Todos los aislamientos de Fusarium examinados mostraron patogenicidad, reprodujeron parte de los síntomas típicos de Fusariosis, necrosis en la raíz, marchitez en la parte aérea de la planta joven, semejantes a los notificados para F. solani (17). Los síntomas en la raíz comenzaron con una decoloración externa del tejido que se tornó café-rojizo, al igual que en el tejido vascular, similares a los descritos para F. oxysporum f. sp. ciceri (10). Las lesiones no presentaron un margen definido y ocasionaron grietas longitudinales en la raíz, que se agrandaron y tornaron color café oscuro (11).
De los reaislamientos realizados, a partir de los síntomas procedentes de las inoculaciones artificiales, se obtuvieron aislamientos con características culturales y morfométricas similares a las de los inoculados, con lo que se comprobaron los postulados de Koch. La incidencia de la enfermedad dependió del aislado; se destacaron los aislamientos 8, 12 y 13 (Tabla 1) con el 100% de plantas infectadas.
Caracterización cultural y morfométrica de los aislamientos de Fusarium spp.
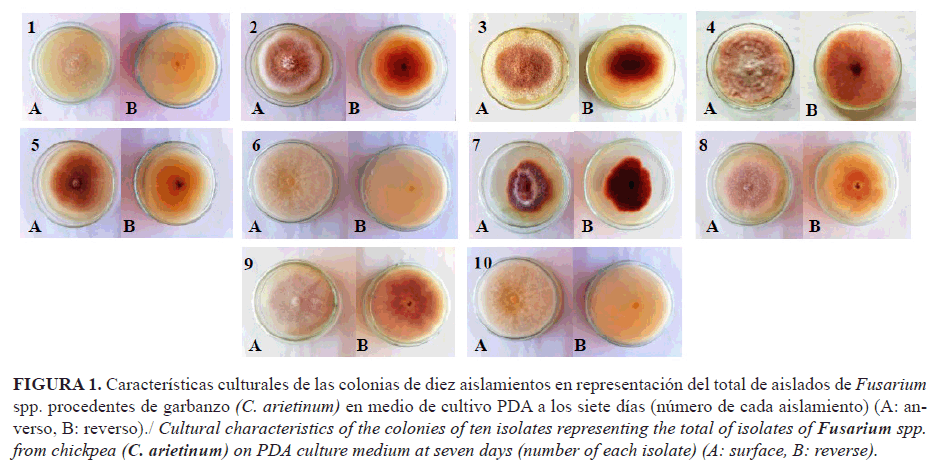
En las colonias de los aislamientos sembrados en medio PDA se observó gran variabilidad, cuyas coloraciones variaron desde violáceo a ligeramente amarillenta, de blanquecino a rojo ladrillo, a lila, a violeta, a rosa salmón; además, tomaron coloraciones rojo violáceo y rojo vino (Fig. 1). Estos resultados coinciden con los informados por Villanueva et al. (18) sobre la existencia de algunas especies de Fusarium que pueden presentar, en medios de cultivo sintético, colonias blancas, cremas, naranjas, pardo, pardo-rojizo, rojo carmín, rosa, púrpura e incluso azuladas. Esta pigmentación puede estar dada por la coloración del micelio o por la difusión de algún pigmento en el medio de cultivo en el cual se desarrolla el hongo. Resultados similares notificaron Arvayo et al. (19), al informar variaciones en la coloración de las colonias de F. oxysporum en medio de cultivo PDA, desde blanquecino, parduzco-blanco, rosáceo-blanco y púrpura-parduzco.
La mayoría de los aislados presentó colonias con bordes ondulados y regulares, con textura afelpada lobulada (Fig. 1).
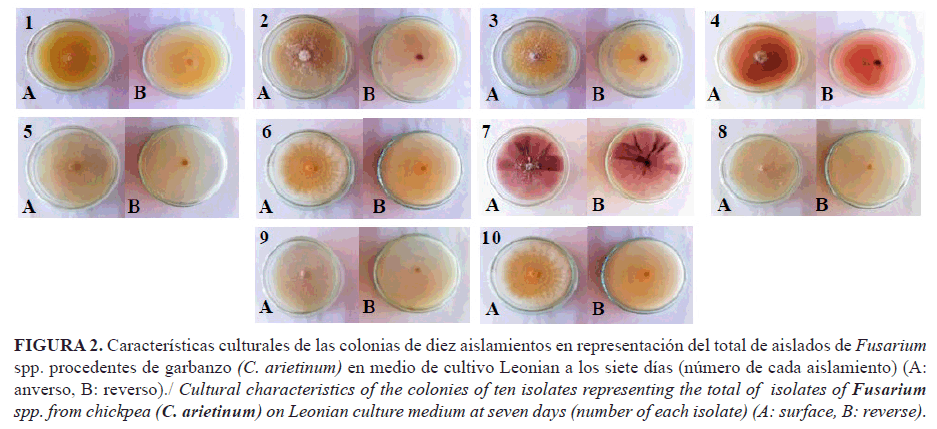
En el medio de cultivo Leonian, la totalidad de los aislamientos desarrollaron colonias de diferentes tonalidades con predominio del amarillo, excepto los aislados 2, 4 y 7 que presentaron tonalidades rojo-violáceas. Los bordes, en su mayoría, fueron regulares y redondeados; la textura rala y radial (Fig. 2).
En el medio CzD, los aislados mostraron colonias con diferentes tonalidades de amarillo, combinadas con blanco y, en algunos casos, con salmón (aislados 6 y 10). Los bordes de las colonias en su mayoría fueron redondeados y regulares, y la textura algodonosa (Fig. 3).
Las colonias de varios aislamientos formaron anillos en los diferentes medios de cultivo; en PDA, los aislados 1, 4, 7, 8 y 12; en Leonian, 6, 7, 10 y 14; y el 2 en CzD, lo que evidencia la influencia del medio de cultivo en la formación de anillos, aspecto que reafirma la variabilidad existente entre los aislamientos.
La presencia de sectores en las colonias se observó en algunos aislamientos y medios de cultivo. Los aislados 4, 8, 9, 11, 15 y 16 presentaron sectores en el medio PDA ; en Leonian el 7, 8, 11 y 12 y en el medio CzD ningún aislado formó sectores.
Aun cuando los aislamientos 8 y 9 proceden de semillas de diferentes líneas genéticas (Tabla 1) y presentan diferencias en la formación de anillos en PDA y de sectores en medio Leonian, mostraron similitud en la textura y la coloración de la colonia en los tres medios, por lo que podrían ser entes de una misma especie.
Sobre la base de los resultados, el medio PDA es idóneo para estudios de caracterización cultural de los aislados de Fusarium spp. de garbanzo, debido a la amplia variabilidad cultural que se puede observar en los aislamientos.
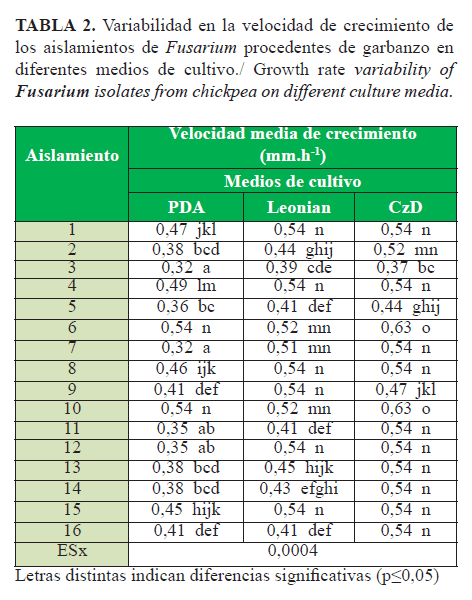
La velocidad de crecimiento fue significativamente mayor en el medio CzD, independientemente de los aislados analizados (Tabla 2). Los aislamientos 6 y 10 mostraron el crecimiento más rápido en los medios de cultivo PDA y CzD; se destacan en medio CzD, cuya velocidad de crecimiento fue de 0,63 mm.h-1.Los aislamientos 1, 4, 8, 9, 12 y 15 mostraron la velocidad más alta en medio de cultivo Leonian. Los aislamientos que alcanzaron la máxima velocidad en los medios, evidencian una asimilación diferencial de los compuestos nutritivos, lo que pudiera ser motivo de una mayor colonización de estos en diferentes tipos de suelo. Lo anterior muestra la existencia de variabilidad entre estos aislamientos (Tabla 2).
Estos resultados coincidieron con lo informado por Dueñas et al. (12) acerca de que los aislamientos de Fusarium procedentes de garbanzo son de rápido crecimiento, entre 0,30-0,54 mm.h-1 en medio PDA, y con lo notificado por Lezcano et al. (20) para F. oxysporum procedente de Leucaena leucocephala (Lam.) de Wit cv. Perú, que mostró una velocidad de crecimiento de 0,38 mm.h-1 en este mismo medio. La alta velocidad de crecimiento de los aislados de Fusarium procedentes de garbanzo pudiera estar directamente relacionada con la colonización del suelo, aspecto que se debe esclarecer en investigaciones futuras.
Por otro lado, aunque los aislamientos esporularon en todos los medios de cultivo, este proceso se favoreció en medio PDA, con diferencias significativas con la obtenida en los medios CzD y Leonian. También, en este medio (PDA) se observó mayor variación en la esporulación entre los aislamientos (Tabla 3).
Los aislados 3 y 9 mostraron la mayor producción de conidios en medio PDA. Sin embargo, los aislados 6 y 10 (en Leonian) y 4, 5 y 16 (en CzD) mostraron las menores producciones de conidios (Tabla 3).
En el medio PDA se obtuvo el menor crecimiento general; sin embargo, aseguró la mayor esporulación por área de los aislamientos de Fusarium. Esto tiene interés para la obtención de inóculos con diferentes fines como las pruebas de patogenicidad, las pruebas de resistencia y la conservación de cultivos, entre otros.
Los 16 aislamientos de Fusarium mostraron gran variabilidad en todos los caracteres analizados. Por otro lado, los aislados 5, 8 y 9 presentaron el 100 % de plantas infectadas, pero no se ubican entre los más destacados respecto al crecimiento y la esporulación; lo que sugiere que estos caracteres cualitativos no presentaron estrecha relación con la patogenicidad de los mismos.
De forma general, esta variabilidad tiene importancia para la identificación de especies, así como para la determinación de presencia de biotipos en el patógeno.
Las características de las colonias y de las estructuras fúngicas, comparadas con las notificadas en la clave de identificación (14), permitieron ubicar los aislamientos en seis especies (Tablas 4 a y b).
Resultados similares para F. oxysporum y F. solani se notificaron por Dueñas et al. (12) y Jiménez et al. (10), los cuales coinciden con las características presentadas para estas especies.
De los aislamientos obtenidos se determinaron, por sus características morfoculturales, seis especies de Fusarium (F. oxysporum f. sp. ciceri, F. solani, F. proliferatum, F. nygamai, F. phyllophilum, F. dlamini), causantes de la fusariosis del garbanzo en áreas de Pinar del Río; las cuatro últimas notificadas por primera vez en el cultivo para Cuba. En este aspecto se profundizará mediante la utilización de técnicas moleculares.
Estos resultados tienen importancia para el programa de selección de cultivares y para la evaluación de estos, así como para el manejo de la enfermedad. Adicionalmente, se detectó variabilidad entre aislamientos de una misma especie, lo que sugiere la posible existencia de biotipos, aspectos en los que se debe profundizar en estudios posteriores. Todo ello refleja la intensa evolución que está sucediendo en el género Fusarium y la variedad de especies capaces de afectar al cultivo de garbanzo en el occidente de Cuba.
REFERENCIAS
1. Sánchez-Yáñez JM, Villegas Moreno J, Vela-Muzquiz GR, Marquez-Benavides L. Respuesta del garbanzo (Cicer arietinum L.) a la inoculación con Azotobacter vineladii y Burkholderia cepacia a dosis reducida de fertilizante nitrogenado. Scientia Agropecuaria. 2014;5:115-120.
2. Echevarría A, Cruz Triana A, Rivero D, Cárdenas RM, Martínez Coca B. Comportamiento agronómico de cultivares de garbanzo (Cicer arietinum L.), en condiciones del municipio Los Palacios, Pinar del Río. Cultivos Tropicales. 2014;35(3):101-106.
3. Shagarodsky T, Morffi L, Chiang ML, Dueñas M, Vega M, López MR, et al. Producción de semilla de Garbanzo (Cicer arietinum L.). Instituto de Investigaciones Fundamentales en Agricultura Tropical «Alejandro de Humboldt» (INIFAT); Unidad de Semilla y Extensión Banao, Sancti Spíritus (INIFAT) y ETIAH, Velasco, Holguín. Conferencia en Curso de Producción de Semillas, INIFAT, Cuba. 2007. 8 pp.
4. González-Leyva M, González-Cruz M, Nápoles-Gallardo E, Baldaquín-Pagan A. Efectividad de algunos biofertilizantes en el cultivo del garbanzo (Cicer arietinum L.) en un suelo Fersialítico Pardo Rojizo Mullido. Innovación Tecnológica. 2012;18(2):1-10.
5. García JL, González L, Shagarodsky T. Enfermedades del garbanzo, su importancia y posibles medidas de control. En: Resultados de las investigaciones para el desarrollo presente y futuro del garbanzo en Cuba. VIII FORUM de Ciencia y Técnica, INIFAT- MINAGRI, Cuba. 1987: 42-52.
6. Tay UJ. Manual para la producción de garbanzo, recomendaciones para la siembra en suelos arcillosos. Chilán, Chile. Instituto de Investigaciones Agropecuarias, 2006; Boletín INIA No 143: 108.
7. De la Fé C, Hernández J. Descripción de seis nuevas líneas de garbanzos (Cicer arietinum L.) en fincas de productores. Cultivos Tropicales. 2011;32(4):44-48.
8. Jiménez Gasco MM. Caracterización molecular de razas patogénicas de Fusarium oxysporum f. sp. ciceris y análisis de su diversidad patogénica. [Tesis doctoral]. Universidad de Córdoba, España. 2001: 171p. [acceso 19 de diciembre de 2016]. Disponible en: http://zeus.plmsc.psu.edu/~jimenez/thesis.pdf.
9. Jiménez-Fernández D, Navas Cortés JA, Montes Borrego M, Jimenez-Dìaz RM, Landa BB. Molecular and pathogenic characterization of Fusarium redolens, a new causal agent of Fusarium Yellows in Chickpea. Plant Disease. 2011;95(7):860-870.
10. Jiménez FD. Estudio de la interacción de Fusarium spp. con cultivares de garbanzo (Cicer arietinum L.) asociados a la Fusariosis Vascular mediante técnicas biotecnológicas. [Tesis doctoral]. Universidad de Córdoba; España. 2011; 285p. [acceso 19 de diciembre de 2016]. Disponible en: http://helvia.uco.es/xmlui/bitstream/handle/10396/4471/9788469416549.pdf?sequence=1.
11. Al-Taae AK, Ali Hadwan H, Eesa Al-Jobory A. Physiological races of Fusarium oxysporum f. sp. ciceris in Iraq. Journal of Life Sciences. 2013;7(10):1070-1075.
12. Dueñas GJM, Shagarodsky T, Fresneda JA, Hernández FY, González J. Caracterización de especies del género Fusarium en el cultivo del garbanzo (Cicer arietinum) en las provincias Ciudad Habana y la Habana. Temas de Ciencia y Tecnología. 2007;11(32):63-66.
13. Booth C. The genus Fusarium. Commonwealth Mycologycal Institute. Ken, England. 1971:273pp.
14. Leslie JF, Summerell BA. The Fusarium Laboratory Manual: Blackwell Publishing Professional, Ames, IA, EEUU, 2006: 388p.
15. Aguaysol NC, Acosta ME, González V, Fogliata G, De Lisi V, Ploper LD. Patógenos detectados en semillas de garbanzo (Cicer arietinum) en Tucumán y áreas de influencia. Avance Agroindustrial 34 (4): 28-30. [acceso 15 de diciembre de 2016]; Disponible en: http://www.eeaoc.org.ar/upload/publicaciones/archivos/384/20140214114810000000.pdf.
16. García JL, González L, Shagarodsky T. Principales enfermedades del cultivo del garbanzo y posibles medidas de control. 9p. [acceso 15 de diciembre de 2016]; Disponible en: http://www.actaf.co.cu/revistas/agrotecnia_05_2008/agrot2008-1/Principales%20enfermedades%20garbanzo.pdf.
17. Alvarez AM, Briner GC. Fusarium solani identified as the causal agent of root rot of chickpea (Cicer arietinum L.) in Chile. Agricultura Técnica (Chile). 1987;47(1):78-79.
18. Villanueva-Arce R, Aguilar-Pompa CA, Gómez YM, Valencia-Del Toro G, Piña-Guzmán AB, Bautista-Baños S. Control de bacterias patógenas y hongos de postcosecha con extractos del pigmento de Gibberella zeae (Fusarium graminearum). Agrociencia. 2013;47(7):691-705.
19. Arvayo-Ortiz RM, Esqueda M, Acedo-Felix E, Sanchez A, Gutierrez A. Morphological variability and races of Fusarium oxysporum f.sp. ciceris associated with Chickpea (Cicer arietinum) crops. Am J Agri & Biol Sci. 2011;6 (1):114-121.
20. Lezcano JC, Martínez B, Alonso O. Caracterización cultural y morfológica e identificación de diez aislamientos de Fusarium procedentes de semillas de Leucaena leucocephala cv. Perú almacenadas. Pastos y Forrajes (Cuba). 2012;35(2):187-196.
Recibido: 6/11/2015
Aceptado: 8/8/2016
Yanisia Duarte Leal, Centro Nacional de Sanidad Agropecuaria (CENSA). Email: yanisia@censa.edu.cu


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}