










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión On-line ISSN 2224-4697
Rev. Protección Veg. vol.31 no.3 La Habana sep.-dic. 2016
ORIGINAL ARTICLE
Morphological and molecular identification of Fusarium species associated with vascular wilt of babaco (Vasconcellea heilbornii var. pentagona Badillo)
Identificación morfológica y molecular de especies de Fusarium asociadas a la marchitez vascular del babaco (Vasconcellea heilbornii var. pentagona Badillo)
Angel Robles-Carrión,I Michel Leiva-Mora,II Lidcay Herrera-Isla,III Aminael Sánchez-Rodríguez,IV Roldán Torres-Gutiérrez,V
ICentro de Biotecnología , Universidad Nacional de Loja. Ciudad Universitaria Guillermo Falconí Espinosa “La Argelia” - PBX: 072547252 - Casilla Letra “S”, Loja-Ecuador.
IIAgritechbio, Riobamba - Ecuador.
IIIFacultad de Ciencias Agropecuarias Universidad Marta Abreu de Las Villas, Santa Clara-Cuba.
IVDepartamento de Ciencias Naturales. Universidad Técnica Particular de Loja-Ecuador.
VCarrera de Ingeniería Ambiental, Universidad Estatal Amazónica, km 2 1/2 vía Puyo a Tena (Paso Lateral), CP 160150, Puyo-Ecuador.
ABSTRACT
Pathogenic isolates of Fusarium spp. associated with babaco (Vasconcellea heilbornii var. pentagona Badillo) were characterized morphologically and molecularly. Babaco roots with symptoms of root rot were collected in major areas in southern Ecuador, and monosporic isolates were obtained. Twenty-one isolates associated with the vascular wilt of babaco (VWB) were characterized culturally, morphologicaly, and molecularly. Amplification of ITS (Internal Transcribed Spacer) subregions placed between the ribosomal genes (rDNA) 18S was performed using specific primers for Fusarium identification. Based on morphological and cultural characteristics, 12 isolates were identified as belonging to Fusarium. Analysis of nucleotide sequences confirmed that all of them belonged to the Fusarium genus: Fusarium oxysporum (Schlechtendahl emend. Snyder & Hansen) (nine isolates), Fusarium camptoceras (Wollenweber & Reinking emend. Marasas & Logrieco) (one isolate), Fusarium temperatum (Scaufl. & Munaut) (one isolate), and Fusarium sp. (one isolate). This is the first report of these four Fusarium species causing VWB in the south of Ecuador.
Key words: Fusarium, species, babaco, indentification.
RESUMEN
Se caracterizaron morfológica y molecularmente los aislados patogénicos de Fusarium asociados al babaco (Vasconcellea heilbornii, Badillo). Se colectaron raíces con síntomas de pudrición radicular en varias áreas de mayor producción en el sur del Ecuador y se obtuvieron aislados monospóricos. Se caracterizaron morfológica y molecularmente 21 aislados asociados con la marchitez vascular del babaco (MVB). Se amplificaron la subregión ITS (Espaciador Transcripto Interno) ubicados entre los genes 18S (ADNr) ribosomales y se utilizaron cebadores específicos para la identificación de Fusarium. Basados en las características morfológicas y moleculares, 12 aislados pertenecen a Fusarium. El análisis de las secuencias de nucleótidos confirman que todos los aislados pertenecieron al género Fusarium: nueve aislados de Fusarium oxysporum (Schlechtendahl emend. Snyder & Hansen), un aislado de Fusarium camptoceras (Wollenweber & Reinking emend. Marasas & Logrieco), un aislado de Fusarium temperatum (Scaufl. & Munaut) y un aislado de Fusarium sp. Este es el primer informe de estas cuatro especies de Fusarium que causaron la MVB en el sur del Ecuador.
Palabras claves: Fusarium, species, babaco, identificación.
INTRODUCTION
The babaco (Vasconcellea heilbornii var. pentagona, Badillo) is an important crop in Ecuador because of the increasing exports of this exotic fruit and its derivatives to the European Union, mainly to Germany and Netherlands with an export volume of 2 351.43 t. This represents more than 4.2 billion dollars, supporting social development in regions where this fruit is grown by small farmers (1, 2). However, diseases are serious limitations to babaco production by causing yield reduction or complete crop loss (3, 4).
One of the most important diseases of babaco is Babaco Vascular Wilt (BVW). The main symptoms of the disease are chlorosis of the lower leaves and rot at the stem base, which leads to epinasty of leaves and plant wilt. Once flaccidity has set in the affected plants, they die within a short period, reducing the production of babaco (3, 5). During the early stages of the pathogen establishment, the growth of babaco seedlings is normal. However, BVW symptoms appear one month after the pathogen infects seedlings. Leaf discoloration usually starts at the bottom of the seedlings and spreads gradually to the top. In the advanced stages of infection, the plant suffers defoliation. BVW disease may occur in both young and old plants. In serious cases the main roots loose their feeder roots, resulting in dieback and eventual death of the seedling (6, 5).
The causal agent of BVW disease is Fusarium oxysporum (3), which was identified and classified as Fusarium oxysporum f sp. vasconcellae (7). This serious disease causes severe crop losses in most highland provinces in which babaco is grown (1). Fusarium infection reduces the economic lifespan of the plantation from 5 years to 1-3 weeks (3, 8). Affected plants may survive for several weeks, but death occurs frequently in the first week after infection (3).
Molecular methods, which are faster and more sensitive than morphological identification, are also used to identify Fusarium species (12). The use of PCR and DNA sequence analysis of internal transcribed spacer (ITS) regions has become a diagnostic routine for the detection, identification, classification and phylogenetic analysis of many fungi at the species level (13, 14). ITS regions sequences are highly variable in Fusarium, and taxon-selective ITS amplification has been used to detect fungal pathogens such as Fusarium (15, 12).
Nevertheless, according to previous studies, only Fusarium oxysporum is the agent causing the vascular wilt of babaco (4, 3), but an integral characterization using cultural, morphological, and molecular criteria of this causal agent have not been done yet. Therefore, the aim of this research was to characterize pathogenic isolates of Fusarium spp. associated with babaco morphologically and molecularly.
MATERIALS AND METHODS
Sample collection and fungal isolation
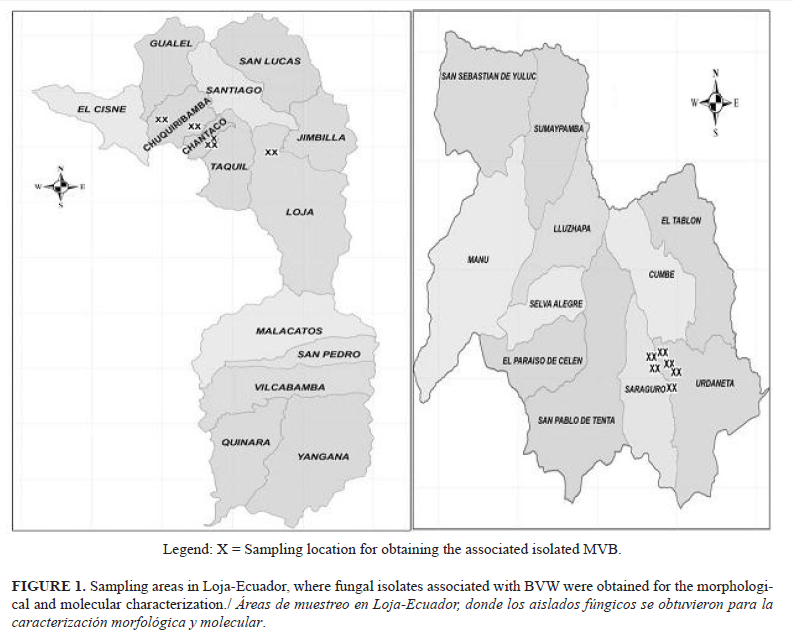
Samples were collected in areas with the highest production of babaco in the province of Loja from January to March 2013. Each area was georeferenced using a Global Positioning System (GPS). Samples were taken in the cantons Loja (locations: Amable Maria, El Carmelo, Chantaco, Reymincha, Gonzabal, and La Retama) and Saraguro (locations: Cochapamba, Tuncarta, Quisquinchir, and Las Lagunas) (Figure 1).
Nine samples of root rots of babaco were collected from the canton of Loja, which is located between the geographical coordinates 03°39’55” to 04°30’38” south latitude and 79°05’58” to 79°32ʹ48” west longitude, with a height of 2 100 mosl; the temperature varies between 16 and 21°C. Twelve samples of root rots of babaco were taken from the canton of Saraguro. It is located between the geographical coordinates 03°31’38’’ to 04°30’20’’ south latitude 79°43’41’’ to 80°60’30’’, with an altitude between 2 000 to 3 500 mosl; the temperature varies between 15 and 20°C (16).
Plants with chlorosis in the lower leaves and rotting at the stem base were selected. They were cut transversely to check vascular stem discoloration (wine-coloured). Root samples and symptomatic stems were packed in Ziploc bags for further analysis in the plant pathology laboratory at the National University of Loja.
Isolation and identification of Fusarium spp. was performed using the following protocol: roots of plants were washed with tap water until they were free of soil. Subsequently, plant tissue of the affected parts of stems and roots were selected and cut into sections of 0.3 to 0.5 cm length. The samples were disinfected in sodium hypochlorite (NaClO) at 2.5% for two minutes, in 70% alcohol for one minute, and rinsed three times in sterile distilled water for one minute. This procedure aims to eliminate those saprophytes and non-pathogenic microorganisms which are found on the plant tissue surface.
Five sections of the affected plant tissue were selected and placed inside Petri dishes (90 X 15 mm) containing PDA medium (Potato 200 g, Dextrose 20 g, Agar 20 g, distilled sterile water 1000 ml; pH 5.5). The plates were then sealed with Parafilm® and incubated at 28°C for 48 hours at 28°C according to Leslie and Summerell (10). Monosporic isolation was done and the fungi were preserved in mineral oil.
Morphological and cultural characterization
The morphological characterization of isolates of Fusarium spp. was performed with the keys proposed by Barnett and Hunter (17), Watanabe (18), and Leslie and Summerell (10), which consisted in determining various characteristic parameters such as: type of mycelium, colony color (surface and reverse) according to the Chart of Rayner (19), radial growth rate, and the observation, quantification, and measurement of the reproductive structures.
For measuring the radial growth speed, 2 mm discs of each fungal isolate were placed in the center of four Petri dishes with PDA medium. . The dishes were sealed and incubated at 28°C in digital incubator (Poleko Series: SN5SE111205, Poland). The radial growth rate calculation (RGR mm.day-1) was performed by comparing the growth at the eighth day of the trial. The RGR was calculated using the formula proposed by Subhani et al. (20):

Where: RGR: Radial growth rate; Fg: Final growth (diameter in mm); Ig: Initial growth (diameter in mm); Et: End time (days); St: Start time (days).
The values of the initial (Ig) and final (Fg) growth and the start (St) and end time (Et) corresponded to the difference between one day of growth and the time elapsed. Isolates were grouped considering the categories of growth: slow (10 mm/day), medium (> 10-12 mm/day), and fast (> 12 mm/day) (21).
In addition, the measurement and characterization of the reproductive structures (micro and microconidia) were done. Microcultures of each isolate were incubated at 28°C for 72 hours. After the incubation time, the samples were observed using optical microscopy (Olympus Series: 1D81617, Germany). One hundred (100) microconidia and 100 macroconidia were randomly taken to determine their average size (30). The conidia were counted in each of the isolates by using the quadrant counting method in the Neubauer chamber with a magnification of 40 X for calculating of final concentration of colony forming units ml-1 (CFU) (22).
Identification of the fungal genus was according to Barnett and Hunter (17), Watanabe (17) Leslie and Summerell (10).
The statistical analysis was performed with the use of the Statgraphic Centurion v.16.1 package. For analysis of RGR, quantification and reproductive structures, measurements were carried out in a completely randomized experimental design with four replications. Differences between isolates were analysed by statistical One Way Anova and Tukey HSD test with a confidence probability of p <0.05.
Molecular characterization
The DNA extraction was performed from axenic-monosporic cultures, according with the methodology proposed by Rojas-Triviño (22) with slight modifications. Single spore isolates were inoculated in PDB (potato dextrose broth) (two Erlenmeyer flasks 250 ml per isolate), two mycelial discs (5 mm in diameter) with active growth were used for inoculation.
The Erlenmeyer flasks were placed in an orbital shaker (Thermo Scientific, IGS60 Germany) at 28°C, constant agitation at 250 rpm for seven days. The harvest of mycelia was done under laminar flow chamber. Filtering was performed with sterile filter paper Whatman # 1 on a glass funnel. The filter paper with the mycelium was placed in a Petri dish for two hours to ensure drying and maceration. The resulting powder maceration was placed in 2 ml tubes (eppendorf).
The fungal DNA was extracted using Purelink Plant Extraction Kit Quick Gel Extraction, Invitrogen TM, (Catalogue K1830-01) according to the manufacturer’s specifications. The extraction product was then analysed by gel electrophoresis on 1% agarose (one g of agarose in 100 ml of 1X TBE buffer).
An internal fragment of ITS (Internal Transcribed Spacer) regions was amplified using the specific primers for Fusarium reported by Abd-Elsalam et al. (12): ITS-Fu-Fwd (5`-CAACTCCCAAACCCCTGTGA-3`) e ITS-Fu-Rev (5`-CGACGATTACCAGTAACGA-3`). PCR amplification contained a final volume of 50μL: 34μL MilliQ water, 5 μL of dNTPs (2 mM each), 5 μl of 10X PCR-buffer, 1.5 μl of each primer (ITS-fuf/ITS-fur) 1.8 μl of MgCl2, 0.2 μl Taq DNA polymerase, 1 μl of extracted DNA. Amplifications were performed with a first cycle of 2 min at 95°C for denaturation, 30 cycles of amplification (1 min at 94°C, 30 s at 54°C and 1 min at 72°C), and a final extension at 72°C for 5 min.
All Polymerase Chain Reactions (PCR) were done in a thermocycler (BioRad serial 583BR01218), amplifications were checked by gel electrophoresis on a 0.7% agarose (1 g of agarose in 150 ml of TAE buffer 1X) with 115 V for 45 min. The gel was stained with 1 ml of ethidium bromide (10 mg ml-1) and bands were visualized on a UV transilluminator according to the procedure of Abd-Elsalam et al. (12).
Sequencing of the amplified ITS fragments
The PCR product was verified by 0.7% agarose gel electrophoresis (band of 400 bp) and the sample was directly purified without removing the gel bands, using a purification kit (Purelink PCR Quick Gel Extraction and Purification, Invitrogen (Catalogue K220001)). Sequencing was performed by capillary electrophoresis and PCR product samples were sent to the Catholic University of Leuven, Belgium, where the sequencing reactions were performed with the same primers used for amplification.
An automatic sequencer Applied Biosystem 3100 was used. Nucleotide sequence assembly and alignment were performed with the Codon Code Aligner program and compared with a search for homologous sequences using the BLAST-N program and algorithm MEGABLAST (23), on the basis data NCBI gene bank (National Center for Biotechnology Information http://www.ncbi.nlm.nih.gov).
RESULTS AND DISCUSSION
Morphological and cultural characterization of fungal isolates from babaco
A total of 21 fungal isolates were obtained from the of roots and stems with symptoms of root rot disease associated with BVW. All isolates showed highly variable macroscopic cultural characteristics, mainly in the pigmentation of the PDA culture medium. The color of the colony surface was white or predominantly white magenta (41.67%) and of the colony reverse was predominantly purple, purple or whitish yellow (25%), pink or magenta pink (25%) and white or yellowish white (25%) (Table 1).
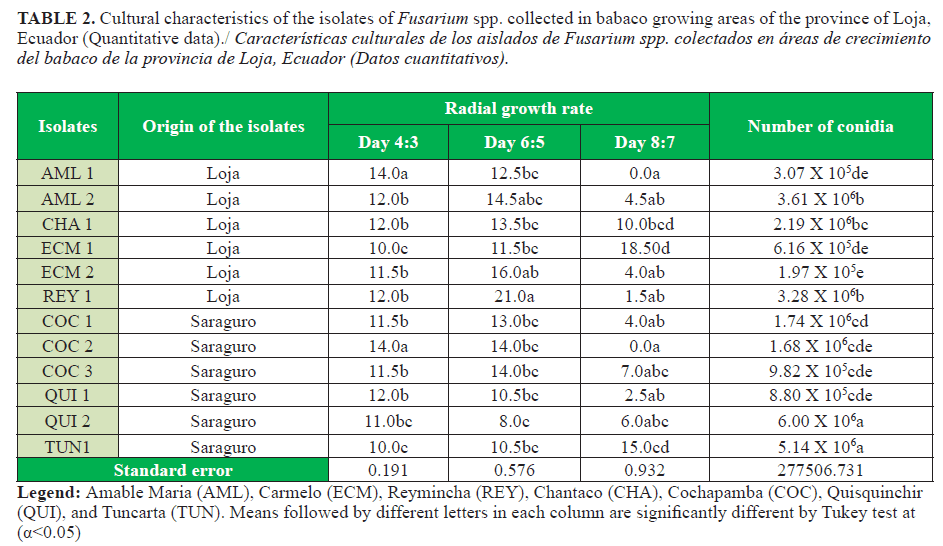
Almost 75% of all isolates showed a type copious or floccose mycelium. According to the radial growth rate AML1 and COC2 were fast growing isolates, which covered the Petri dish completely before the last evaluation period (Table 2).
Isolates QUI2 (Quisquinchir) and TUN1 (Tuncarta) produced the longest conidia, both isolates belonged to the canton of Saraguro. Therefore, there was variability in this parameter among the isolates obtained (Table 3). A great variability was observed among the twelve isolates, which suggested the possible coexistence of several Fusarium species in the same lesions.
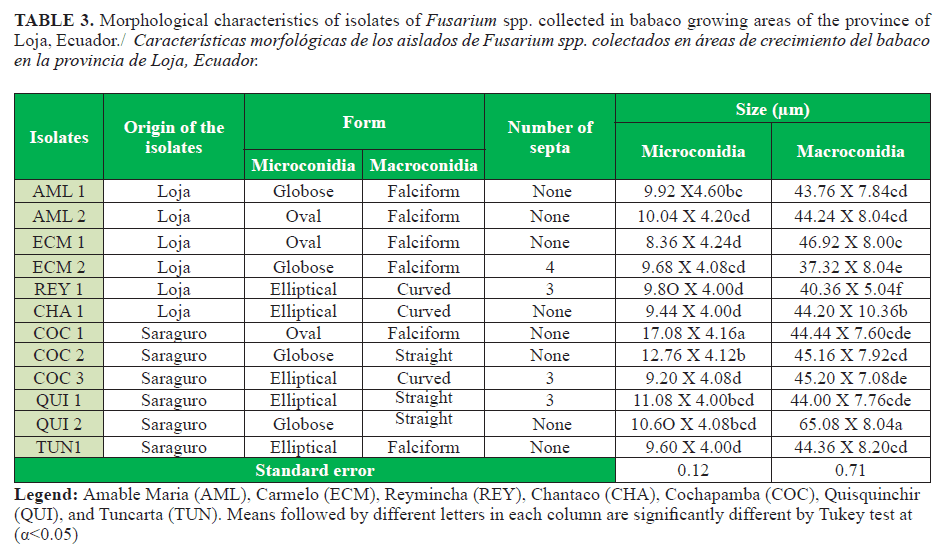
The twelve isolates associated with MVB showed hyaline, globose, oval or elliptical microconidia. None of the conidia showed septa, and they were 17.08 to 8 36 X 4.60 to 4.0 µm (length x width). In addition, all isolates showed hyaline macroconidia boat shaped, curved or straight pointed at the ends with 65.08 to 37.32 X 8.20 to 5.04 µm (length x width). It should be noted that the great majority of the isolates showed macroconidia without septa, except the isolates ECM 2, REY1, COC 3, and QUI 1, in which 3-4 septa were observed (Table 3).
Based on the morphological and cultural characteristics, the twelve fungal isolates associated with VWB sampled in different agroclimatic zones of Loja province belonged to several species of Fusarium spp. Ochoa and Fonseca (24) reported F. oxysporum isolates from babaco tissues showing a cottony mycelium and a typical pale-pink to purplish discoloration of the species when it sporulated. On the other hand, according to Leslie and Summerell (10), F. oxysporum mycelium produces dark purple or dark magenta pigments when is grown on PDA medium, never clear or whitish yellow. However, the pigmentation observed by Dubey et al. (21) was violet, yellow and grey. Furthermore, Shahnazi et al. (25) reported F. solani isolates from black pepper producing pigmentation that was cream, white or whitish cream, wine, coffee, black, and grey. In our study, the different isolates of Fusarium produced pigmentation with different colors such as purple, purple or whitish yellow, pink or magenta pink, and white or yellowish white, mainly in the centre of the colony. This result showed a great variability among different isolates of Fusarium associated with babaco vascular wilt.
Ochoa and Fonseca (3) and Irzykowska et al. (26), argued that F. oxysporum was the causal agent of BVW, which had a rich mycelium (cottony or floccose). On the contrary, the descriptions by Dubey et al. (21) assert that F. oxysporum is a predominantly floccose mycelium. Conversely, Lezcano et al. (26) and Shahnazi et al. (24) reported that Fusarium species had an aerial mycelium, and sometimes plush. In our study, the results based on the type of mycelium of Fusarium agreed with those reported by the first three authors above cited; moreover, the results concerning the morphological diversity of the isolates obtained in the present study suggested the presence of various Fusarium species including in the species F. oxysporum.
Variability was also observed among isolates in the daily mycelial growth, which might be caused by nutrient defficiency (5). The macroconidia were less abundant compared with the microconidia. Morphology of these structures varied and the sickle, curved, and straight shapes were the most frequent. Most isolates showed microconidia without septa, except the isolates El Carmelo 2 (ECM 2) and Reymincha (REY 1). The microconidia were globular, oval, and elliptical to oval. The macro and microconidia size were outside the range informed by Watanabe (17) and Leslie and Summerell (10), with an average of (29,1-45 µm x 2,9-4,7 µm) for F. oxysporum macro and microconidia, which suggested thar other species of the genus might be present (Table 3).
Similarly, Córdova-Albores et al. (11) reported that the morphological characteristics of the species of Fusarium spp. isolated from gladiolus corms were greatly variable. The macroconidia showed varied forms: papillate, blunt, hooked, and foot shaped. Likewise, the microconidia were oval, reniform, pyriform, and globose. The great variability in macroconidia size (87,51 to 82,54 µm x 50,50 to 48,72 µm), with three to four septa, was evident, and the microconidia were non-septate or with one septum. Similar results were observed in the present study.
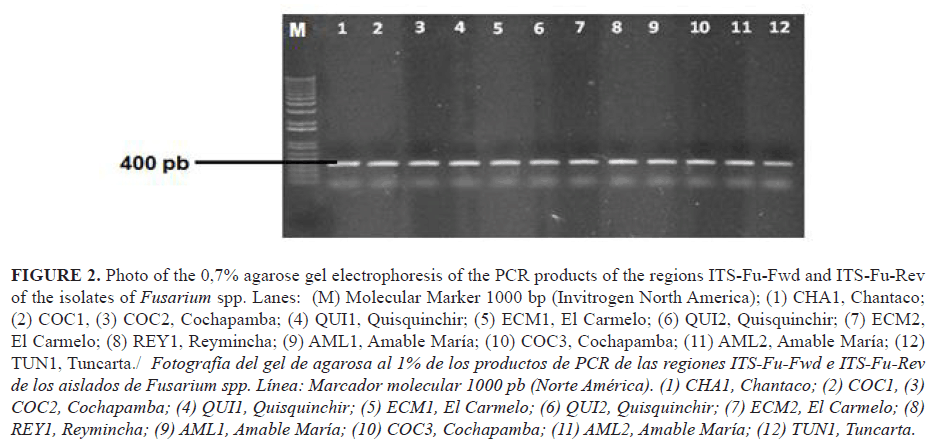
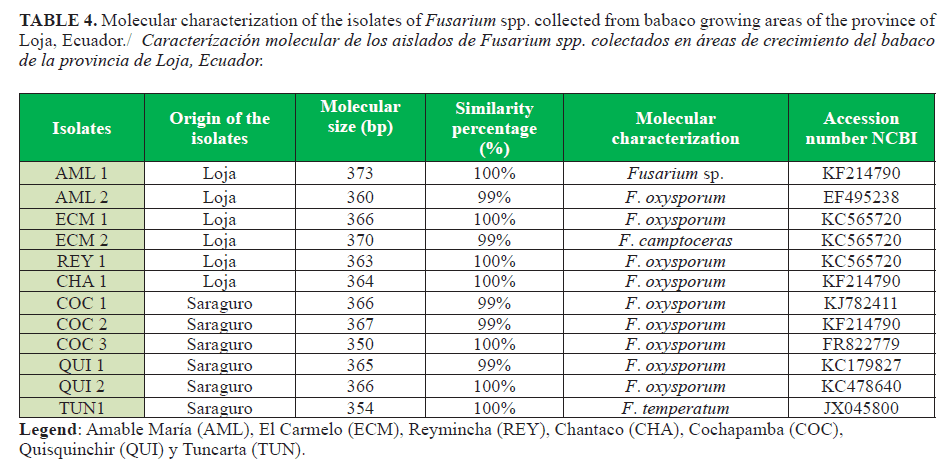
The twelve isolates associated with BVW identified by their cultural and morphological characteristics were analized for identification by using molecular techniques PCR products of approximety 400 bp were obtained (Figure 2). Comparative analysis of sequenced ITS region showed an identity of 99 to 100% with isolates of Fusarium spp. from the Genbank. According to the sequence homology, four species of Fusarium (F. camptoceras, F. temperatum, F. oxysporum, and Fusarium sp.) were identified (Table 4).
Based on this result, Abd-Elsalam et al. (12) used the primers ITS1 and ITS4 to identify F. oxysporum f. sp. vasinfectum, F. oxysporum, Fusarium moniliforme, and F. solani, and they obtained bands with molecular size from 550 to 570 bp, results which did not agree with the those obtained in the present work. Nevertheless, these authors did not perform sequencing of the PCR products. Similarly, Abd-Elsalam et al. (12) designed the specific primers ITS-Fu-Fwd and Rev ITS-Fu-Rev for a rapid identification of Fusarium. These primers showed high specificity for amplifying a Fusarium region with a molecular size of 389 bp. In this research, the PCR products obtained with these primers showed an estimated molecular size of 400 bp (Table 4).
In addition, the studies conducted by Córdova-Albores et al. (2014) using the universal primers ITS 1, ITS2, and ITS5 to identify species of Fusarium (F. oxysporum, F. solani, and F. proliferatum) from gladiolus, showed bands with a molecular size of 500 bp . Furthermore, they also used specific primers (ITS-Fu-Fwd and ITS-Fu-Rev) for identification of species of Fusarium, which had a molecular size of approximately 410-429 bp. These results are within the range observed in our research.
Therefore, of the twelve molecularly characterized isolates, only 11 corresponding to F. camptoceras, F. temperatum, or F. oxysporum could be identified. The remaining isolate showed similarity to align the BLAST 100% and it was identified as Fusarium. sp. It was evident that using the molecular tools used in this research allowed the identification of microorganisms, integrating morphological, cultural, and molecular data. Future studies will help the understanding of the pathogenic mechanisms allowing the development of integrated strategies for a more effective and environmentally friendly management (27).
In the present study, the cultural, morphological and molecular characterization determined showed great variability among species of Fusarium spp. reporting new species of Fusarium (F. camptoceras, F. temperatum, F. oxysporum, and Fusarium sp.) associated with vascular wilt of babaco (BVW) in southern Ecuador.
CONCLUSIONS
From 21 samples from babaco roots associated with BVW in Loja province and based on the cultural and morphological characteristics of fungal isolates, we identified 12 isolates belonging to the genus Fusarium spp. on wilting babaco plants. Likewise, based on the molecular features, four groups of species of the genus Fusarium (nine isolates of F. oxysporum, one isolate of F. camptoceras, one isolate of F. temperatum, and one of Fusarium sp.) were identified. This is the first report about the interaction of these four Fusarium species, which cause the BVW disease.
ACKNOWLEDGMENTS
The authors acknowledge the National University of Loja, the Flemish University Council (VLIR) of Belgium and Secretaría Nacional de Ciencia, Tecnología e Innovación (SENESCYT) for the financial support to conduct this study. To Dr. Roldan Torres Gutiérrez and Dr. Aminael Sánchez Rodríguez for their contribution to the conduction of the research.
REFERENCES
1. Bravo C, Larriva W, Minchala L. Manejo integrado de la marchitez o fusariosis (Fusarium oxysporum) en el cultivo del babaco. Quito, Ecuador: Instituto Nacional Autónomo de Investigaciones Agropecuarias. 2012: 17.
2. García P. Evaluación de la tolerancia de cinco accesiones de Vasconcelleas a Fusarium sp. como posible portainjertos para babaco (Vasconcelleae x heilbornii) bajo cubierta plástica en la estación experimental del austro de INIAP. [Tesis presentada en opción al título de Magister en Gestión de la Producción de Flores y Frutas Andinas para Exportación] Universidad Técnica de Ambato. Ecuador. 2011; 74 pp.
3. Ochoa J, Fonseca G, Ellis M. First report of Fusarium wilt of Babaco (Carica× heilbornii var. pentagona) in Ecuador. Plant Disease. 2000;84(2):199.
4. Argotti E, Cazar M, Motte E, Cedeño V. Análisis molecular de la región ITS de Fusarium spp., agente causal de la marchitez vascular de Vasconcellea heilbornii y Solanum quiotense en Ecuador. ESPE Ciencia y Tecnología. 2011;3(1):25-36.
5. Agrios GN. Plant Pathology. 5th ed. United States of America: Elsiever; 2005; 534-550.
6. Nelson P, Toussoun T, Morasas W. Fusarium species: An Illustred Manual for Identification. Pennsylvania, United States of America: Pennsylvania State University Press; 1983; 193.
7. Ochoa J, Yangari B, Ellis M, Williams R. Dos nuevas formae specialis de Fusarium oxysporum causantes de la marchitez vascular del babaco (Carica heilbornii var. pentagona) y de la marchitez vascular de la naranjilla (Solanum quitoense) en Ecuador. Fitopatología. 2004;39(1):10-17.
8. Quillay N. Determinación de la capacidad embriogénica de babaco (Vasconcelleae x heilbornii) a partir de óvulos y hojas multiplicadas in vitro vía embriogénesis somática. [Tesis presentada en opción al título de Magister en Gestión de la Producción de Flores y Frutas Andinas para Exportación] Universidad Técnica de Ambato. Ecuador. 2011; 114 pp.
9. Biswas S, Pandey N, Rajik M. Inductions of defense response in tomato against Fusarium wilt through inorganic chemicals as inducers. J Plant Pathol Microb. 2012;3(4):3-4.
10. Leslie JF, Summerell BA. The Fusarium Laboratory Manual. United States of America: Blackwell. 2006; 387.
11. Córdova L, Bautista S, Martínez J, Barrera L, Hernández M, Cruz A. Morphological and molecular characterization of pathogenic isolates of Fusarium spp. obtained from gladiolus corms and their sensitivity to Jatropha curcas L. oil. Afr J Microbiol Res. 2014;8(8):724-733.
12. Abd-Elsalam KA, Aly IN, Abdel-Satar MA, Khalil MS, Verreet JA. PCR identification of Fusarium genus based on nuclear ribosomal-DNA sequence data. Afr J Biotechnol. 2004;2(4):82-85.
13. Hafizi R, Salleh B, Latiffah Z. Morphological and molecular characterization of Fusarium solani and Fusarium oxysporum associated with crown disease of oil palm. Brazilian Journal of Microbiology. 2013;44(3):959-968.
14. Zhao B, Yan J, Zhang S, Liu X, Gao Z. Phylogeny and pathogenicity of Fusarium spp. isolated from greenhouse melon soil in Liaoning Province. Saudi Journal of Biological Sciences. 2014;21(4):374-379.
15. White TJ, Bruns T, Lee S, Taylor J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR protocols: a guide to methods and applications. 1990;18:315-22.
16. Scheldeman X. Distribution and potential of Cherimoya (Annona cherimola Mill.) and highland papayas (Vasconcelleae spp.) in Ecuador. [Tesis presentada en opción al grado científico de Doctor in Applied Biological Sciences]. Faculty of Agricultural and Applied Biological Sciences. Ghent University. Belgium. 190 p.
17. Barnett H, Hunter BB. Illustrated genera of imperfect fungi. 4th ed. MacMillan Publishing Company: United States of America; 1987; 218.
18. Watanabe T. Pictorial Atlas of Soil and Seed Fungi: Morphologies of Cultured Fungi and Key to Species. 2nd ed. United States of America: CRC Press; 2002; 506.
19. Rayner RW. A mycological colour chart. 1970; 34 p.
20. Subhani MN, Talib S, Sahi AL, Hussain S, Iqbal J, Hussain N. Management of chickpea wilt caused by Fusarium oxysporum f. sp. ciceris through antagonistic microorganisms. Canadian Journal of Plant Protection. 2013;1(1):1-6.
21. Dubey S, Singh SR, Singh B. Morphological and pathogenic variability of Indian isolates of Fusarium oxysporum f. sp. ciceris causing chickpea wilt. Archives of Phytopathology and Plant Protection. 2010;43(2):174-190.
22. Rojas-Triviño A. Conceptos y Práctica de Microbiología General. Palmira, Colombia: Universidad Nacional de Colombia Sede Palmira. 2011; 161.
23. Altschul S, Gish W, Miller E, Meyers E, Lipman D. Basic local alignment search tool. J Mol Biol. 1990;215(3):403-410.
24. Shahnazi S, Meon S, Vadamalai G, Ahmad K, Nejat N. Morphological and molecular characterization of Fusarium spp. associated with yellowing disease of black pepper (Piper nigrum L.) in Malaysia. J Gen Plant Pathol. 2012;78(3):160-169.
25. Irzykowska L, Bocianowski J, Waskiewicz A, Weber Z, Karolewski Z, Golinski P, et al. Genetic variation of Fusarium oxysporum isolates forming fumonisin B1 and moniliformin. J Appl Genetics. 2012;2(53):237-247.
26. Lezcano J, Martínez B, Alonso O. Caracterización cultural y morfológica e identificación de diez aislamientos de Fusarium procedentes de semillas de Leucaena leucocephala cv. Perú almacenadas. Pastos y Forrajes. 2012;35(2):187-196.
27. Hill A, Reeves P, Larson R, Fenwick A, Hanson L, Panella L. Genetic variability among isolates of Fusarium oxysporum from sugar beet. Plant Pathology. 2011;60(3):496-505.
28. Mwaniki P, Abang M, Wagara I, Wolukau J, Schroers H. Morphology, pathogenicity and molecular identification of Fusarium spp. from wilting eggplants in Tanzania. 10th African Crop Science Conference Proceedings, Maputo, Mozambique, 10-13 October 2011. African Crop Science Society.
29. Vengadaramana A, de Costa D. Morphological and pathogenic variations of the causal organisms of leaf twister disease of red onion (Allium cepa L.) in Jaffna district of Sri Lanka. Tropical Agricultural Research. 2014;25(3):412-31.
Recibido: 22/04/2016
Aceptado: 27/10/2016
Angel Robles-Carrión, Centro de Biotecnología , Universidad Nacional de Loja. Email: anroca1980@gmail.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}