










Meu SciELO
 Serviços customizados
Serviços customizadosServiços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Protección Vegetal
versão impressa ISSN 1010-2752versão On-line ISSN 2224-4697
Rev. Protección Veg. vol.32 no.3 La Habana set.-dez. 2017
ARTÍCULO ORIGINAL
Actividad antagónica de Pseudomonas spp.fluorescentes ante Alternaria solani Sorauer, patógeno de la papa(Solanum tuberosum L.)
Antagonist activity of fluorescent Pseudomonas against Alternaria solani Sorauer pathogen of potato crop (Solanum tuberosum L.)
Edwin Ronnie Gakegne1*, Odaylín Plasencia Márquez1, Annia Hernández-Rodríguez2, Yeised de La Fe Pérez2; Benedicto Martínez Coca1*
1Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, Mayabeque, CP 32700, Cuba.
2Departamento de Microbiología y Virología, Facultad de Biología, Universidad de La Habana, Calle 25 # 455 e/ J e I. Plaza. La Habana. Cuba.
RESUMEN
Este trabajo tuvo como objetivo determinar el efecto antagonista de tres cepas de Pseudomonas spp.fluorescentes frente a una cepa patógena de Alternaria solani Sorauer, procedente del cultivo de la papa (Solanum tuberosum L.). Para ello se realizaron ensayos en cultivo dual in vitro y se determinaron el efecto antagónico de las cepas de Pseudomonas spp. en dos medios de cultivo, la acción preventiva, el efecto fungistático y de los metabolitos de las cepas del antagonista frente a A. solani. En cada experimento, los tratamientos se compararon mediante análisis de varianza y prueba de Tukey; además, los porcentajes en la acción preventiva y en el efecto fungistático se procesaron según la prueba de comparación de proporciones. Las cepas de Pseudomonas spp. inhibieron, marcadamente, el crecimiento del patógeno; se destaca Pseudomonas protegens Pf-5 por presentar los mejores resultados en todos los ensayos realizados y, también, por exhibir aparente micoparasitismo. Los resultados evidenciaron que esta cepa pudiera ser promisoria para el control biológico de este patógeno en el cultivo de la papa.
Palabras clave: control biológico, Pseudomonas, Tizón temprano.
ABSTRACT
The objective of this work was to determine the antagonistic effect of three strains of fluorescent Pseudomonas spp. against a pathogenic strain of A. solani isolated from potato. Two culture media were tested in in vitro dual culture assays to determine the antagonistic effect of the Pseudomonas strains, the preventive action, and the fungistatic effect of the bacterial strains and of their metabolites. In each experiment, the treatments were compared by analysis of variance and Tukey's test; in addition, the percentages of the preventive action and the fungistatic effect were tested according to the comparison of proportions. Pseudomonas spp. strains markedly inhibited the growth of the pathogen; Pseudomonas protegens Pf-5 excelled for being the best in all the tests performed, besides the apparent mycoparasitism. These results showed that this latter strain could be promising for the biological control of this pathogen in the potato crop.
Key words: biological control, Pseudomonas, early blight.
INTRODUCCIÓN
Solanum tuberosum L. es originaria de los Andes y es uno de los aportes más importantes de la región andina al mundo, por ser uno de los cultivos alimenticios más consumidos y apreciados, además por contribuir, de manera importante, con el fortalecimiento de la seguridad alimentaria de la humanidad (1). Representa uno de los cuatro alimentos más importantes del mundo junto al maíz, el trigo y el arroz (2,3).
Uno de los factores que limita la producción de papa, en la mayoría de los países tropicales, es la incidencia de enfermedades. Entre estas, el Tizón temprano (Alternaria solani Sorauer) adquiere vital importancia por estar ampliamente difundida a nivel global, pues su agente causal infecta el follaje, los tallos y los tubérculos, y afecta el rendimiento en áreas con condiciones climáticas favorables para su desarrollo (4).
Tradicionalmente, el control de este patógeno se realizó con agroquímicos (5); sin embargo, su uso trae como consecuencia efectos nocivos al ambiente debido a su toxicidad y residualidad.
La necesidad de buscar y aplicar alternativas amigables con el medio ambiente, que no afecten la salud humana, y dada la importancia de este cultivo a nivel nacional y mundial, hace que el uso de bacterias como agentes de control biológico (ACB) sea una opción a considerar para el control de este patógeno. Dentro de estas destacan las bacterias del género Pseudomonas y especialmente las del grupo fluorescente (6,7).
Las especies de Pseudomonas fluorescentes son bacterias baciliformes aerobias, que poseen varios flagelos polares. Son conocidas por su capacidad de estimular el crecimiento de las plantas que viven en contacto con ellas (8). Estas bacterias son bacilos Gram-negativo, rectos o ligeramente curvados y saprofíticas. Se pueden encontrar en ecosistemas acuáticos y en el suelo. No forman esporas y las temperaturas más favorables para su desarrollo están entre 25 y 30°C, aunque puede crecer desde 5 a 42°C. Requieren pH neutro y no crecen en condiciones ácidas (pH ≤ 4,5). Sus flagelos polares hacen posible su movimiento activo en líquidos. Su pigmento fluorescente (fluoresceína) las hacen reaccionar frente a la luz ultravioleta, aunque algunas veces no fluorecen cuando el cultivo es reciente o después de varios cultivos en el laboratorio (6,8,9).
Estas bacterias se pueden encontrar en la superficie foliar y al nivel de la raíz, especialmente en la rizosfera, donde la colonización de estas depende de características como la planta y el tipo, el manejo y la irrigación del suelo (6,7). Ellas se destacan como promotoras del crecimiento vegetal (6,9) o como inhibidoras de crecimiento de algunos hongos y/o bacterias fitopatógenas (8). Los mecanismos involucrados en la inhibición incluyen la producción de antibióticos (6,7,10), la producción de quelatos de hierro y enzimas hidrolíticas (10), así como la competencia por sitios nutricionalmente favorables (o ricos en nutrientes) (6,7,10).
Este trabajo tuvo como objetivo determinar el efecto antagonista de tres cepas de Pseudomonas spp.fluorescentes frente a una cepa patógena de A. solani, procedente de papa.
MATERIALES Y MÉTODOS
Origen de los cultivos microbianos y producción de inóculos
El trabajo se realizó en el Laboratorio de Bacteriología Vegetal del Centro Nacional de Sanidad Agropecuaria (CENSA) (Latitud 22.991867; Longitud -82.153892), provincia Mayabeque, Cuba.
Como agente de control biológico se utilizaron las cepas Pseudomonas protegens Pf-5 (cepa de referencia ATCC® BAA-477™) aislada de suelo de Texas, USA (7), Pseudomonas putida AJ13 y Pseudomonas fluorescens AI05, obtenidas de la rizosfera de la variedad de arroz J-104 sobre suelo Gley Vértico Crómico-Nodular Ferruginoso, con actividad antagónica ante Pyricularia grisea (Sacc.) (11), procedentes de la Colección de Cultivos Microbianos de la Facultad de Biología de la Universidad de La Habana, Cuba.
Para la obtención de los inóculos a utilizar en todos los experimentos, se sembraron las tres cepas en placas Petri contentivas de medio King B (BIOCEN) y se incubaron durante 24 h a 28°C. Posteriormente, se pasaron a tubos de ensayo con 5 ml de medio Caldo Nutriente (CN) (BIOCEN) y se incubaron durante 48 h con agitación en zaranda de marca Thys 2 (28°C; 150 rpm). Finalmente se prepararon suspensiones celulares con una concentración final aproximada de 108 UFC, las que se utilizaron en cada uno de los experimentos.
Como agente patógeno, se usó el aislamiento de A. solani No. 160, previamente aislado de hojas de papa con síntomas típicos de Tizón temprano, procedentes de la provincia Las Tunas, perteneciente a la colección del Laboratorio de Micología Vegetal del CENSA. Para la obtención del micelio del hongo, se tomaron discos de 5mm de diámetro y se sembraron en placas Petri con medio Papa Dextrosa Agar (PDA) (BIOCEN) durante siete días a 30±1°C y oscuridad.
Los inóculos del hongo, para todos los experimentos, consistieron en discos de 5 mm de diámetro, que se tomaron a partir de la periferia de las colonias. Todas las placas inoculadas con el hongo se incubaron a 30±1°C y en oscuridad.
Actividad antagónica de cepas de Pseudomonas spp. fluorescentes frente a A. solani en diferentes medios de cultivo
Para la realización de este experimento, se embebieron discos de micelio del patógeno durante cinco minutos, en cada una de las suspensiones celulares de las bacterias. De los discos tratados, unos se sembraron, de forma individual, en el centro de las placas Petri (90 mm de Ø) con medio PDA y los otros, en agar nutriente (AN) de igual manera. Las placas, con los discos del patógeno, se incubaron a 30±1°C en condiciones de oscuridad. Los controles se trataron de igual forma, con la diferencia de que unos discos del hongo se embebieron en agua destilada estéril y otros en medio de cultivo CN. Para cada tratamiento se realizaron tres réplicas.
La actividad antagónica de las bacterias se determinó a través de mediciones del diámetro de crecimiento de las colonias del hongo en presencia de las bacterias a los 3, 5 y 7 días, utilizando una regla graduada.
Acción preventiva de cepas de Pseudomonas spp. fluorescentes frente a A. solani
En placas Petri contentivas de PDA se inocularon, en el centro de la placa, 10 µl de cada una de las suspensiones bacterianas por separado y se extendieron, uniformemente, con una espátula de Drigalsky.
A partir de micelio de A. solani de colonias que tenían una semana de crecimiento en placas Petri con medio PDA, se realizaron suspensiones ajustadas a una concentración de 108 UFC, con agua destilada estéril. La suspensión micelial se asperjó, asépticamente, sobre las placas con crecimiento bacteriano de 24 h. Los testigos fueron las placas Petri con medio PDA que, a las 24 h, se inocularon con la suspensión micelial. Todas las placas se incubaron a 30±1°C. Se realizaron cinco réplicas por variante. Se evaluó el número de colonias a los 3, 5 y 7 días. Se calcularon los porcentajes de inhibición según la fórmula de Abbott (13) y se procesaron según la prueba de comparación de proporciones.
Efecto fungistático de cepas de Pseudomonas spp. fluorescentes frente a A. solani
En este experimento se procedió según describió Santoyo et al. (12) con algunas modificaciones; la suspensión micelial de A. solani se asperjó sobre placas Petri contentivas de PDA, que se incubaron por 24 h. Transcurrido este tiempo, las placas con el hongo se inocularon con 10 µl de cada una de las suspensiones bacterianas de forma individual en el centro de la placa y se extendieron, uniformemente, con una espátula de Drigalsky.
Los controles consistieron en placas Petri inoculadas solamente con el hongo, se realizaron cinco réplicas por variante y se evaluó el número de colonias del patógeno a los 3, 5 y 7 días. Se calcularon los porcentajes de inhibición según la fórmula de Abbolt (13) y se procesaron según la prueba de comparación de proporciones.
Efecto de los metabolitos de cepas de Pseudomonas spp. fluorescentes sobre A. solani
De las suspensiones bacterianas obtenidas previamente, la mitad (2,5 ml) se filtró a través de filtros miliporos de 0,20 µm (Sartorios).
Por cinco minutos se embebieron los discos de micelio del patógeno en la suspensión bacteriana y en el filtrado de cada una de las cepas, de manera separada. Finalmente, en ambas variantes, los discos del hongo se sembraron en placas Petri con medio de cultivo PDA. Como controles se usaron placas sembradas con discos del hongo embebidos en agua destilada estéril y en medio de cultivo CN, por cinco minutos, individualmente. Se hicieron cinco réplicas por variante. Se evaluó el crecimiento del patógeno con una regla graduada a los 3, 5 y 7 días.
Efecto antagonista de cepas de Pseudomonas spp. fluorescentes frente a A. solani mediante difusión en medio de cultivo
Se procedió según describió Yesid (14) para el método de perforación (pocitos) con algunas modificaciones. Se realizaron cuatro perforaciones de 5 mm de diámetro en placas con PDA y en cada una de ellas se depositaron, con una micropipeta, 50 µl de cada una de las suspensiones bacterianas de forma individual.
En el centro de cada placa se colocó un disco de micelio A. solani obtenido de colonias de siete días de crecimiento. Como testigo se dejaron placas Petri, en las cuales se inocularon los pocitos con agua destilada estéril. Se evaluó el crecimiento micelial del patógeno con una regla graduada a los 3, 5 y 7 días.
Determinación del efecto de los metabolitos inhibidores del crecimiento de A. solani en la cepa seleccionada
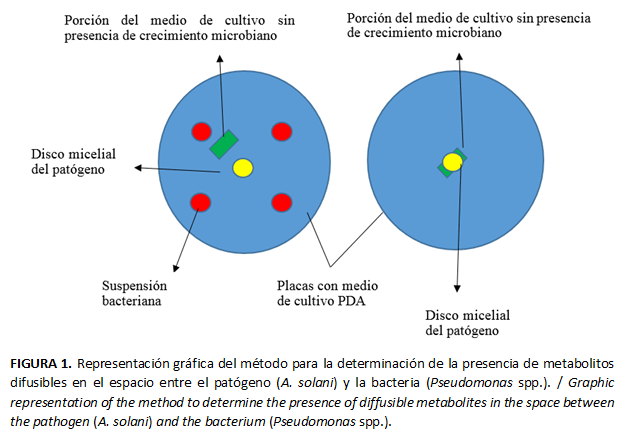
Se extrajeron porciones del medio de cultivo sin presencia de crecimiento microbiano, en la zona de interacción bacteria-patógeno. Las porciones extraídas se depositaron en el centro de placas Petri de 7 cm de Ø, con medio PDA y sobre estas se inoculó un disco de micelio del patógeno (Fig. 1). Como testigo se dejaron placas sembradas de igual manera que las anteriores, pero con porciones de medio de cultivo de las placas testigo, sin micelio del patógeno del experimento anterior. Se realizaron cinco réplicas de cada tratamiento. Se evaluó el crecimiento micelial del patógeno a los 3, 5 y 7 días con una regla graduada.
Los datos recolectados en todos los experimentos se tabularon en Microsoft Excel y se procesaron mediante análisis de varianza simple. Las medias se compararon según prueba de comparación de rangos múltiples de Tukey para un nivel de 95 %. Se empleó el paquete estadístico InfoStat/Profesional versión 1.1 (15).
RESULTADOS Y DISCUSIÓN
Actividad antagónica de cepas de Pseudomonas spp.fluorescentes frente a A. solani en diferentes medios de cultivo
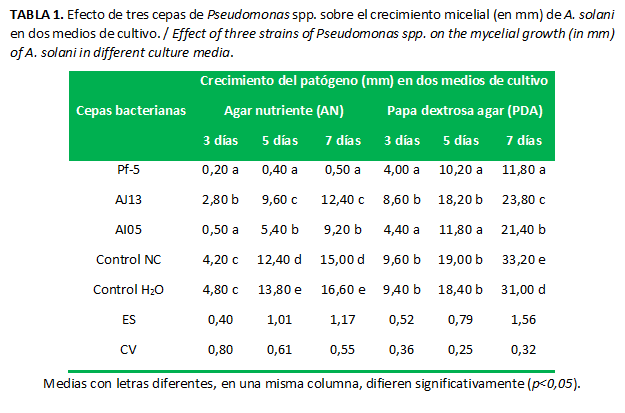
Las cepas de Pseudomonas spp. inhibieron el crecimiento de A. solani en el medio de cultivo AN, con diferencias significativas respecto a los controles en todos los momentos de evaluación (Tabla 1). En el medio PDA también se observó efecto inhibitorio, con la excepción de la cepa AJ13, cuyo efecto tuvo diferencias significativas solo el séptimo día de la evaluación, con respecto a los controles. En ambos medios de cultivo, de forma general, P. protegens cepa Pf-5 mostró el mejor comportamiento al compararla con las otras dos cepas.
El medio PDA favoreció el crecimiento de hongo (Tabla 1). En sus investigaciones, Guerra et al. (16) observaron que el medio de cultivo AN favoreció el crecimiento de Pseudomonas, ya que en el medio de cultivo PDA su desarrollo fue mínimo. Teniendo en cuenta ambos aspectos se infiere que para seleccionar la cepa bacteriana con mayor efecto sobre el hongo fitopatógeno es más adecuado realizar los experimentos utilizando medio PDA, a fin de que el hongo se vea favorecido en su crecimiento y que la bacteria, a pesar de estar en condiciones no óptimas, demuestre sus potencialidades como agente de control biológico.
Acción preventiva de las cepas de Pseudomonas frente a A. solani
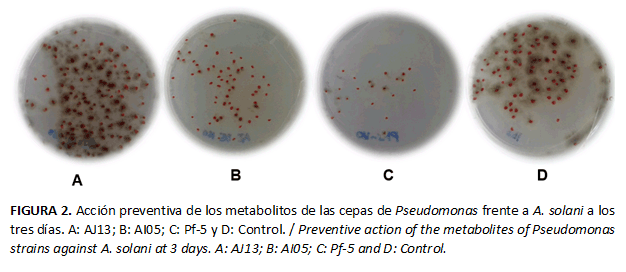
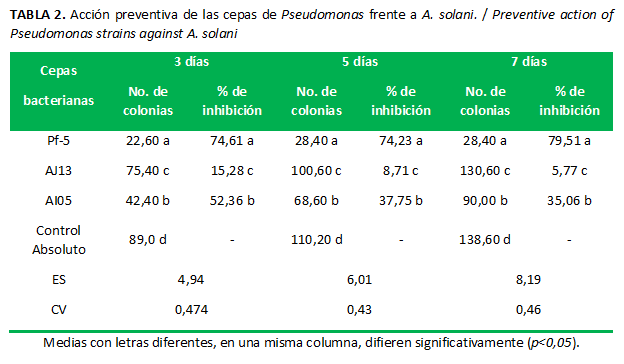
Las cepas de Pseudomonas spp.inhibieron al hongo, expresado en un menor número de colonias del patógeno en comparación con el control absoluto.En orden descendente, en cuanto a efecto observado, lacepa Pf-5 fue la mejor, seguida de AI05 y AJ13, que presentó el menor efecto inhibitorio en todos los momentos evaluados (Fig. 2). A partir del quinto día el comportamiento de la cepa Pf-5 se mantuvo constante, su efecto preventivo fue aún mayor con el tiempo. Sin embargo, el efecto de las cepas AI05 y AJ13 disminuyó en los momentos evaluativos. (Tabla 2)
Efecto fungistático de cepas de Pseudomonas spp. frente a A. solani
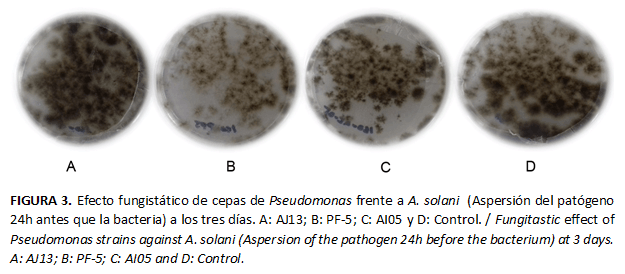
Las cepas de Pseudomonas inhibieron el número de colonias de A. solani de 24 horas de edad. El efecto fungistático más evidente se produjo por acción de la cepa Pf-5, con diferencias significativas, en comparación con el testigo y las restantes cepas, durante todo el periodo de evaluación. La Figura 3 muestra que la cepa Pf-5 (la más eficaz en la reducción del número de colonias) también provoca cambios en la intensidad de color pardo de las colonias de A. solani que disminuye en este caso, en comparación con las restantes cepas y el testigo. De forma similar al resultado anterior, se mantiene la tendencia de que la cepa AJ13 muestra la menor actividad antagónica, a pesar de que no presentó diferencias significativas al quinto día con la cepa AI05. (Tabla 3)
Estos resultados coinciden con los obtenidos por Fernández (17), quien analizó diferentes cepas de Pseudomonas y encontró diferencias en su comportamiento y entre los colores del micelio del hongo patógeno producto de la acción de la bacteria, aun cuando esta especie es considerada eficaz para controlar enfermedades foliares. Esta atenuación del color se hace más visible a los siete días, por lo que este aspecto debe ser estudiado posteriormente, a fin de esclarecer si existe alguna relación con otros daños que provoca la bacteria sobre el hongo y que influyan o no sobre su capacidad como patógeno.
Santoyo et al. (12) encontraron, en estudios similares, que el mayor grado de inhibición se observó en el tratamiento donde se inoculó la cepa de P. fluorescens ZUM80 con 24 h de anterioridad a Colletotrichum lindemuthianum Briosi & Cavaray (76 %), comparadas con las variantes de inoculación simultánea (30 %) o 24 h después (28 %). Estos autores observaron un patrón similar para el patógeno Colletotrichum gloeosporioides (Penz.) Penz. & Sacc, ya que al inocularlo 24 h después de la cepa bacteriana, su crecimiento se restringió hasta en el 72 %. Ellos encontraron que el crecimiento del micelio de Phytophthora cinnamomi Rands se inhibió hasta en un 70 %, cuando la cepa bacteriana se inoculó 24 h después, mientras que cuando se inoculó 24 h antes que P. fluorescens, la inhibición fue totalmente nula. Los resultados de este trabajo coinciden, en parte, con estos informes, porque en este caso la bacteria logró inhibir al patógeno cuando se inoculó 24 h antes o después de este.
La acción antagónica demostrada en este trabajo también coincide con los resultados obtenidos por Fleitas y Grabowski (18), quienes observaron que las cepas de P. fluorescens lograron inhibir 17,1 % del crecimiento de Bipolaris sp., 22,4 % de Curvularia spp. y 39,3 % de Exserohilumo turcicum K.J. Leonard & Suggs.
De acuerdo con Santoyo et al. (12), los altos porcentajes de inhibición detectados, cuando P. fluorescens ZUM80 se inoculó 24 h antes que los patógenos, pudieran deberse a que las bacterias necesitan tiempo para sintetizar los metabolitos secundarios que causan la inhibición. Varios autores (12,18) plantearon que Pseudomonas tiene un alto potencial como agente de control biológico mediante la producción de sideróforos, pero requiere tiempo para sintetizarlos.
Efecto de los metabolitos de cepas de Pseudomonas spp. fluorescentes sobre A. solani
De manera general, se encontró que solo los metabolitos de la cepa Pf-5 produjeron un efecto inhibitorio significativamente diferente a los controles analizados, aun cuando no se encontraron diferencias significativas entre el efecto causado por todas las cepas bacterianas (Tabla 4). El análisis del efecto provocado por las suspensiones bacterianas sobre A. solani mostró diferencias significativas para todas las cepas respecto a los controles y nuevamente se destacó la cepa Pf-5. Los tratamientos donde se utilizaron las cepas tuvieron un mejor efecto que aquellos donde se utilizaron los metabolitos (Fig. 4), con diferencias significativas entre sí.
Hernández et al. (11) demostraron que todos presentaron efecto antagónico ante el fitopatógeno al enfrentar los 17 aislamientos de P. fluorescens a la cepa de Curvularia.
El efecto antagónico encontrado fue similar al observado por Lee et al.(19) ante Fusarium oxysporum Schltdl., Pythium sp. y Gaeumannomyces graminis (Sacc.) Arx & D.L. Los resultados que se presentan están en correspondencia con los de Guerra et al. (16) y Pérez et al. (8), que demostraron la efectividad antagónica de P. fluorescens contra Fusarium oxysporum Schltdl. Por otro lado, Hernández et al. (20) manifestaron que las cepas de Pseudomonas spp. tienen actividad antagonista in vitro y podrían ser potencialmente eficientes en el control de Phytophthora palmivora (Butler) en el cultivo de cacao (Theobroma cacao L.).
Majumder et al. (7) determinaron el éxito de P. fluorescens como control biológico frente a enfermedades fúngicas de las plantas, debido a sus diversos mecanismos de acción como son la producción de diversos metabolitos antifúngicos que contribuyen, entre otros, a la inhibición del crecimiento (21), la producción de sideróforos, la liberación de antibióticos, la producción de enzimas líticas, el parasitismo, la competencia por nutrientes y espacio, la inducción de resistencia, así como su capacidad para producir una gran variedad de metabolitos secundarios tóxicos a hongos y bacterias fitopatógenos, como los antibióticos (22). Los mismos señalaron que estas bacterias no utilizan un solo mecanismo, sino que en algunos casos pueden presentarse varios al unísono o en sucesión.
En estudios realizados por Izzeddin y Medina (23) se inhibió el crecimiento de Fusarium monilliforme Sheldon y Phytophthora spp. cuando estos se enfrentaron a cepas de Pseudomonas y obtuvieron halos de inhibición mayores a 5 mm de diámetro.
Efecto antagonista de cepas de Pseudomonas spp. frente a A. solani mediante difusión en medio de cultivo
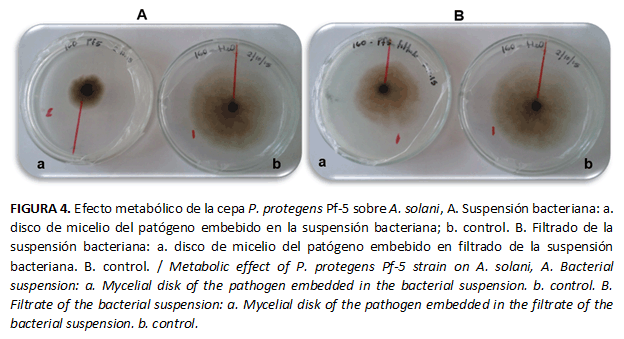
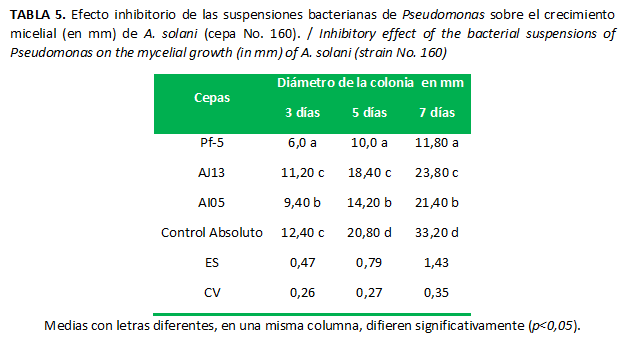
Las suspensiones bacterianas de las tres cepas de Pseudomonas mostraron un efecto inhibitorio sobre A. solani, aun sin estar en contacto con el patógeno. Los metabolitos producidos por la cepa Pf-5 provocaron la mayor inhibición del crecimiento del hongo (Fig.5), con diferencias significativas con respecto al resto de los tratamientos. Aun cuando los resultados obtenidos con la cepa AJ13 fueron diferentes significativamente a los del control a partir del quinto día, en esta cepa se alcanzó la menor inhibición (Tabla 5).
Villamil et al. (24) evaluaron la eficacia de la actividad antagónica de microorganismos nativos aislados del suelo y observaron que una cepa de P. fluorescens redujo el crecimiento de Moniliophthora roreri Cif & Par. Además, documentaron que es posible que el mecanismo ejercido por la bacteria es la antibiosis, debido a la producción de metabolitos secundarios volátiles o difusibles, pero que en cualquiera de los casos, sin entrar en contacto físico con el patógeno, estos pueden inhibir o restringir su crecimiento.
Determinación de la presencia de metabolitos de la cepa de P. protegens Pf-5, inhibidores del crecimiento de A. solani
Se demostró que en el espacio existente en la placa, entre el patógeno y la cepa PF-5, hubo presencia de metabolitos difusos en el medio de cultivo, pues lograron disminuir el crecimiento del patógeno sobre el disco procedente de dicha zona (Fig.6). En la literatura consultada no se encontraron antecedentes del análisis de los modos de acción de esta bacteria para el control de A. solani.
El método más común para controlar las enfermedades en los cultivos es el uso de compuestos químicos; sin embargo, se comprobó que estos tienen consecuencias negativas en el ambiente y en la salud humana (7). Entre las nuevas alternativas para manejar las enfermedades en los cultivos, el uso de microorganismos con capacidades para restringir el crecimiento de fitopatógenos se convierte en una opción viable.
Los principales mecanismos involucrados en el control biológico de fitopatógenos con el uso de bacterias antagonistas son: la competencia por los nutrientes, minerales y espacio; la síntesis de metabolitos, tales como sideróforos, antibióticos, toxinas y biosurfactantes, y la inducción de resistencia sistémica en la planta (20,25).
De acuerdo con Plascencia et al. (22), uno de los grupos de microorganismos evaluados como controladores de fitopatógenos son las bacterias antagonistas, entre las que se destacan las pertenecientes al género Pseudomonas, debido a su capacidad para producir una gran variedad de metabolitos secundarios tóxicos a hongos y bacterias fitopatógenos.
No obstante, aunque es reconocido el efecto como agentes de control biológico de las bacterias de la especie Pseudomonas fluorescens, en este trabajo se demuestra que existen diferencias en la eficacia y en los mecanismos de acción entre aislados. De ahí la necesidad de realizar estudios que profundicen en sus características, modos de acción y metabolitos, que sirvan de base para la selección de aquellas cepas que resulten ser las más promisorias.
CONCLUSIONES
Pseudomonas fluorescentes pueden ser utilizadas como agente de control biológico, ya que tienen la capacidad de inhibir el crecimiento de fitopatógenos en condiciones in vitro. La cepa de P. protegens PF-5 tiene efecto inhibitorio, fungicida y fungistático sobre A. solani, lo que demuestra sus potencialidades y las diferencias entre las cepas estudiadas en cuanto a sus mecanismos de acción. No obstante, deben realizarse otros estudios a escala de campo para comprobar los verdaderos efectos de esta cepa en el control de la plaga y el manejo integrado de la enfermedad en el cultivo de la papa.
1. Gasteiz V. Avances en ciencia y desarrollo de la Patata para una Agricultura Sostenible. III Congreso Iberoamericano de Investigación y Desarrollo en Patata. 2008; 1-161. ISBN: 978-84-457-2804-8.
2. Salomón D, Castillo H, Estévez V, Arzuaga S, Ortiz C, Torres W, Caballero N, Ramiro VE. Estudio de la floración y producción de semilla botánica de polinización libre en genotipos de papa (Solanum tuberosum, L.) en Cuba. Cultivos Tropicales. 2012; 33 (2): 61-67 ISSN: 0258-5936.
3. Ministerio de Agricultura. Instructivo Técnico para la Producción de Papa en Cuba. República de Cuba. Ministerio de la Agricultura. Dirección de Agricultura. 2016; 1-62.
4. Salazar JR, López SR, Castaño AE, Ospina TB, Rave AS, Velásquez JC. Manejo fitosanitario del cultivo de la papa. ICA. Bogotá D.C. Colombia. 2011ª. 1-35. ISSN: 00.09.31.11.
5. Rodríguez M, Stefanova M. Control biológico del tizón temprano (Alternaria solani Sorauer) en el cultivo de la papa (Solanum tuberosum L.) en condiciones de campo. Fitosanidad. 2005; 9 (4): 35-37. ISSN: 1562-3009.
6. Seaton SC, Silby WM. Genetics and functional genomics of the Pseudomonas fluorescens Group. Springer-Verlag Berlin Heidelberg 2014; 99-127. DOI: 10.1007/978-3-642-55378-3_5.
7. Majumder D, Kongbrailatpam J, Suting E, Kangjam B, Lyngdoh D. Pseudomonas fluorescens: A potential biocontrol agent for management of fungal diseases of crop plants. Springer International Publishing AG, Part of Springer Science Business Media. Fungal Biology. 2014; 317-342. ISBN 978-1-4939-1188-2. DOI: 10.1007/978-1-4939-1188-2_11.
8. Pérez Á, Coto A, Echemendía P, Ávila QG. Pseudomonas fluorescens Migula, ¿control biológico o patógeno? Rev. Protección Veg. 2015; 30 (3): 225-234. ISSN: 2224-4697.
9. Xie K, Peng H, Hu H, Wang W, Zhang X. OxyR, an important oxidative stress regulator to phenazines production andhydrogen peroxide resistance in Pseudomonas chlororaphis GP72. Microbiological Research. 2013; 168: 646– 653. DOI: org/10.1016/j.micres.2013.05.001.
10. Kidarsa TA, Shaffer BT, Goebel NC, Roberts DP, Buyer JS, Johnson A, Kobayashi DY, Zabriskie T, Paulsen I, Loper JE. Genes expressed by the biological control bacterium Pseudomonas protegens Pf-5 on seed surfaces under the control of the global regulators GacA and RpoS. Environmental Microbiology. 2013; 15(3): 716–735. DOI:10.1111/1462-2920.12066.
11. Hernández-Rodríguez A, Plasencia L, Rives R, Díaz A, Almaguer C, Acebo Y. Identificación de aislamientos autóctonos de Pseudomonas fluorescentes con actividad antagónica ante Curvularia spp. Rev. Protección Veg. 2010; 25 (3) 181-189. ISSN 2224-4697.
12. Santoyo G, Valencia E, Orozco M, Peña C, Farías R. Papel de los sideróforos en la actividad antagónica de Pseudomonas fluorescens Zum80 hacia hongos fitopatógenos. Chapingo, MX. Terra Latinoamericana. 2010; 28 (1): 53-60. ISSN: 2395-8030.
13. CIBA-GEIGY. Manual para ensayos de campo en protección vegetal. Segunda edición revisada y ampliada. Editado y publicado por Werner Püntener, División Agricultura, CIBA-GEIGY S.A. Basilea, Suiza. 1981; 33-35.
14. Yesid A y Sánchez L. Determinación de metabolitos secundarios a partir de Bacillus subtilis con efecto biocontrolador sobre Fusarium sp. Nova - Publicación Científica en Ciencias Biomédicas, 2012; 10 (18): 135 – 250. ISSN: 1794-2470.
15. Di Rienzo J, Balzarini M, Casanoves F, González L, Tablada M, Walter CR. InfoStat versión 1.1. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. 2002.
16. Guerra G, Betancourth C, Salazar C. Antagonismo de Pseudomonas fluorescens Migula frente a Fusarium oxysporum f sp. pisi Schtdl en arveja Pisum sativum L. Rev. U.D.C.A Act. & Div. Cient. 2011; 14(2): 33 – 42. ISSN: 0123-4226.
17. Fernández LVO. Microorganismos antagonistas para el control fitosanitario. Manejo Integrado de Plagas (Costa Rica) 2001; (62): 96 - 100. ISSN: 1016-0469.
18. Fleitas C, Grabowski OJ. Control biológico del complejo de hongos causantes de la mancha foliar en maíz dulce (Zea mays var. saccharata) con bacterias benéficas. Investig. Agrar. 2014; 16(2): 83-92. ISSN: 2305-0683.
19. Lee K, Byung O, Seralathan KK. Advances in Plant Growth Promoting Rhizobacteria for Biological Control of plant diseases. D.K. Maheshwari (ed.), Bacteria in Agrobiology: Disease Management. 2013; 1-13. DOI 10.1007/978-3-642-33639-3_1.
20. Hernández-Rodríguez A, Ruíz Y, Acebo Y, Miguélez Y, Heydrich M. Antagonistas microbianos para el manejo de la pudrición negra del fruto en Theobroma cacao L. Estado actual y perspectivas de uso en Cuba. Rev. Protección Veg. 2014; 29 (1): 11-19. ISSN: 2224-4697.
21. Abraham A, Laing, M, Bower JP. Isolation and in vivo screening of yeast and Bacillus antagonists for the control of Penicillium digitatum of citrus fruit. Biol Control. 2010; 53: 32-38. DOI:10.1016/j.biocontrol.2009.12.009.
22. Plascencia T, Olalde P, Mena V, Ceja T, Venegas G, Oyoque S, Angoa PM. Antagonismo in vitro de aislados bacterianos de fresa comercial y silvestre vs Botrytis cinerea y Rhizopus stolonifer. Ra Ximhai. Universidad Autónoma Indígena de México. Mochicahui, El Fuerte, Sinaloa. 2012; 8 (3): 103-110. ISSN: 1665-0441.
23. Izzeddin A, Medina TL. Efecto del control biológico por antagonistas sobre fitopatógenos en vegetales de consumo humano. Salus Valencia. 2011; 15 (3):8-18. ISSN 1316-7138.
24. Villamil C, Blanco V, Viteri RE. Evaluación in vitro de microorganismos nativos por su antagonismo contra Moniliophthora roreri Cif & Par en Cacao (Theobroma cacao L.). Revista Facultad Nacional de Agronomía. 2012; 65 (1): 6305-6315. ISSN: 0304-2847.
25. Acebo Y, Hernández-Rodríguez A, Heydrich M, El Jaziri M, Hernández AN. Management of black pod rot in cacao (Theobroma cacao L.). Fruits. 2012; 67:41-48. DOI: 10.1051/fruits/2011065.
Recibido: 8/6/2017
Aceptado: 18/10/2017
* Autor para correspondencia: Edwin Ronnie Gakegne. E-mail: gakegne@yahoo.fr, ronnie@censa.edu.cu
* Autor para correspondencia: Benedicto Martínez Coca. E-mail: bmcoca@censa.edu.cu


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}