










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión impresa ISSN 1010-2752versión On-line ISSN 2224-4697
Rev. Protección Veg. vol.32 no.3 La Habana set.-dic. 2017
ARTÍCULO ORIGINAL
Antagonismo in vitro de aislamientos de Trichoderma asperellum Samuels, Lieckfeldt & Nirenberg frente a Sclerotium rolfsii Sacc.
In vitro antagonism of Trichoderma asperellum Samuels, Lieckfeldt & Nirenberg isolates against Sclerotium rolfsii Sacc.
Yanisia Duarte-Leal1*, Alexis Lamz-Piedra2, Benedicto Martínez-Coca1
1Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, CP 32 700, Mayabeque, Cuba.
2Instituto Nacional de Ciencias Agrícolas (INCA), Mayabeque, Cuba.
RESUMEN
El presente trabajo tuvo como objetivo determinar los modos de acción (antibiosis, competencia por espacio y nutrientes, micoparasitismo) de diez cepas de Trichoderma asperellum Samuels, Lieckfeldt & Nirenberg (Ta.), frente a dos aislados de Sclerotium rolfsii Sacc. (Scl) procedentes de frijol (Phaseolus vulgaris L.). La evaluación del antagonismo in vitro se realizó por el método de cultivo dual, a través del cual se determinaron los diferentes modos de acción. La mayoría de las cepas de Ta. se ubicaron en el grado 2 de la escala de Bell, con excepción de Ta.17, que mostró grado 5. Los metabolitos producidos por las cepas Ta. 28, Ta. 13 y Ta. 90 presentaron el mayor efecto inhibitorio del crecimiento de Scl 5, mientras que para Scl 6 fueron las cepas Ta. 13, Ta. 79, Ta. 28, Ta. 75, Ta. 85 y Ta. 90. La mayor competencia por nutrientes se observó con la cepa Ta. 28. Los resultados del micoparasitismo mostraron que la mejor cepa fue Ta. 90 con tres tipos de interacciones hifales entre ambos hongos, seguido de Ta.13 y Ta. 56 con tres y dos tipos de interacciones, respectivamente. Acorde a los modos de acción que poseen las cepas Ta. 13, Ta. 28 y Ta. 90 pueden ser valoradas como posibles candidatas de ingredientes activos de bioplaguicidas para el control de S. rolfsii en el cultivo del frijol.
Palabras clave: control biológico, interacciones hifales, patógeno de suelo.
ABSTRACT
The objective of this work was to determine the modes of action (antibiosis, competition for space and nutrients, and mycoparasitism) of ten strains of T. asperellum Samuels, Lieckfeldt & Nirenberg (Ta.) against two isolates of S. rolfsii Sacc. (Scl) isolated from common bean (Phaseolus vulgaris L.). In vitro antagonism was assessed by the dual culture method, through which the different modes of action were determined. Most strains of Ta. were located in grade 2 of the Bell scale, except Ta. 17, which showed grade 5. The metabolites produced by strains Ta. 28, Ta. 13, and Ta. 90 showed the highest inhibitory effect on Scl 5 growth, whereas those produced by the strains Ta. 13, Ta. 79, Ta. 28, Ta. 75, Ta. 85, and Ta .90 did on Scl 6. Greater competition for nutrients was observed with the strain Ta.28. Mycoparasitism results showed the best strain was Ta. 90 with three types of hyphal interactions between the two fungi, followed by Ta. 13 and Ta. 56 with three and two types of interactions, respectively. According to the modes of action of the strains Ta. 13, Ta. 28, and Ta. 90, they can be evaluated as possible candidates for active ingredients of biopesticides for the control of S. rolfsii in the bean crop.
Key words: biological control, hyphal interactions, soil pathogen.
INTRODUCCIÓN
Sclerotium rolfsii Sacc. es un hongo fitopatógeno que causa pudriciones de raíces, tallos, tubérculos y frutos en numerosos cultivos, como son: frijol (Phaseolus vulgaris L.), manzano (Malus domestica Borkh.), remolacha (Beta vulgaris L.), girasol (Helianthus annus L.), papa (Solanum tuberosum L.), soya (Glycine max L.), arroz (Oryza sativa L.), pimiento (Capsicum annuum L.) y cebolla (Allium cepa L.), entre otros (1,2).
En frijol común, este agente fitopatógeno ocasiona la enfermedad conocida como Tizón sureño, que provoca amarillamiento, marchitez, caída de las hojas y finalmente la muerte de la planta. En el cuello de la raíz se observa una lesión necrótica y, sobre los tejidos afectados, un crecimiento micelial blanco donde se forman los esclerocios del hongo, inicialmente blancos y que, posteriormente, se tornan color marrón oscuro (1).
Esta enfermedad causa pérdidas considerables en el frijol. Su control se realiza con el uso de prácticas preventivas, como rotación de cultivos, uso de semillas sanas, preparación del suelo y otras tácticas que incluyen el control biológico y químico. Los productos químicos demuestran ser eficientes inicialmente, pero la enfermedad persiste en el campo debido al poco efecto de estos sobre los esclerocios. Además, son cuestionados por su impacto ambiental, sobre la salud humana y la resistencia que desarrollan los microorganismos, lo cual genera una creciente y justificada preocupación por la contaminación, el deterioro ambiental y el desequilibrio de los ecosistemas (3,4,5).
Uno de los métodos utilizados para disminuir el uso de productos químicos es la aplicación de microorganismos antagonistas. Dentro de estos, uno de los más promisorios para combatir enfermedades fúngicas en las plantas es el género Trichoderma con especies y cepas que exhiben versatilidad, adaptabilidad y fácil manipulación, lo que permite que se utilicen en el manejo de enfermedades causadas por agentes fitopatógenos cuyo hábitat es el suelo, como Phytophthora, Rhizoctonia, Pythium, Fusarium y Sclerotium, entre otros (6,7,8).
El presente trabajo tuvo como objetivo determinar los mecanismos de acción de diez cepas de Trichoderma asperellum Samuels, Lieckfeldt & Nirenberg (Ta.) frente a dos aislados de S. rolfsii (Scl) procedentes de frijol.
MATERIALES Y MÉTODOS
El ensayo se realizó entre enero y abril de 2017, en el Laboratorio de Micología Vegetal del Centro Nacional de Sanidad Agropecuaria (CENSA), municipio San José de las Lajas, Mayabeque, Cuba.
Evaluación de los diferentes modos de acción de T. asperellum frente a S. rolfsii
El trabajo se desarrolló con diez cepas de T. asperellum denominados Ta. 1, Ta. 13, Ta. 17, Ta. 28, Ta. 56, Ta. 75, Ta. 78, Ta. 79, Ta. 85 y Ta. 90 (9) y dos aislamientos de S. rolfsii (Scl 5 y Scl 6), obtenidos de raíces de plantas de frijol enfermas, provenientes del Instituto Nacional de Ciencias Agrícolas (INCA), Mayabeque, Cuba. Todos los hongos utilizados pertenecen a la Colección de cultivos microbianos del Laboratorio de Micología Vegetal del CENSA.
Se evaluaron los modos de acción (competencia por espacio, micoparasitismo y antibiosis) de las cepas de T. asperellum frente a S. rolfsii, utilizando el método de cultivo dual (CD) descrito por Martínez y Solano (10).
Se incluyó un testigo del agente fitopatógeno sin el antagonista. Todos los hongos, en los diferentes tratamientos, se incubaron a 28±2°C y en oscuridad constante.
Antibiosis
La antibiosis se evaluó por el crecimiento de S. rolfsii en el CD antes del momento de contacto entre ambos hongos (T. asperellum-S. rolfsii) a las 48 h, respecto al testigo (S. rolfsii).
Competencia por espacio
El experimento se desarrolló y se evaluó según lo referido por Reyes et al. (7). Las evaluaciones se llevaron a cabo hasta los siete días y la clasificación como antagonista de los aislamientos se realizó según la escala descrita por Bell et al. (11). Se determinó el Porcentaje de Inhibición del Crecimiento Radial (PICR) según la fórmula:
![]()
donde:
r1 - Crecimiento radial de la colonia en el control.
r2 - Crecimiento radial del aislamiento patógeno enfrentado al antagonista.
Micoparasitismo
Para evaluar los tipos de interacciones hifales (enrollamiento, penetración, vacuolización, granulación y lisis), se tomaron tres muestras de la zona de interacción entre ambos hongos por cada réplica (placa Petri, tres en total) y se depositaron en portaobjetos sobre una gota de lactofenol. Las observaciones se realizaron con microscopio óptico modelo N-800M (aumento de 400x). Se evaluaron nueve réplicas por tratamiento (cepas de T. asperellum).
Los datos obtenidos se procesaron por un Análisis de Varianza (ANOVA), y las medias se compararon según la Dócima de Rangos Múltiples de Duncan para p≤0,05, utilizando el paquete estadístico INFOSTAT Profesional ver. 2.1.
RESULTADOS Y DISCUSIÓN
Antibiosis
Las cepas de T. asperellum evaluadas inhibieron, significativamente, el crecimiento del agente fitopatógeno desde las 48 h, en comparación con el testigo, aunque este efecto se observó, más marcado, a las 72 h (Fig. 1). En el enfrentamiento con el aislado Scl. 5, la cepa Ta. 28 presentó mayor efecto inhibitorio, aunque sin diferir significativamente de las cepas Ta.13 y Ta. 90 (Fig. 1A); mientras que, para el aislamiento Scl. 6, se destacaron Ta. 13 y Ta. 79 sin diferir de Ta. 28, Ta. 75, Ta. 85 y Ta. 90 (Fig. 1B). La acción fungistática que se originó antes del contacto hifal infiere la secreción de metabolitos difusibles al medio por parte de las cepas de Trichoderma.
El efecto inhibitorio de Trichoderma sobre otros hongos antes del contacto hifal se explicó con la secreción de metabolitos volátiles y no volátiles al medio (gliotoxina, viridina, trichodermina, suzukacilina, alameticina, dermadina, trichotecenos y trichorzianina), que poseen efectos fungistáticos, y a la producción de enzimas extracelulares (peptinasas, cutinasas, glucanasas y quitinasas) difusibles hacia el medio, que afectan al patógeno (6,8,5). Estos compuestos pueden limitar la colonización del sustrato por los fitopatógenos, en su radio de acción (12).
Al analizar el crecimiento del agente fitopatógeno frente a las cepas más destacadas (Ta. 13, Ta. 28 y Ta. 90) cada 24 h, se observó que desde las 48 h las cepas inhibieron el crecimiento del aislado Scl. 5 (Fig. 2A); mientras que la cepa Ta. 90 inhibió el crecimiento del aislado Scl. 6 (Fig. 2B). Lo anterior es una evidencia de que estas cepas segregan metabolitos al medio de cultivo desde periodos tempranos de la interacción. La respuesta del aislado Scl. 6 frente a las cepas Ta. 13 y Ta. 28 en este periodo de tiempo pudiera estar relacionada con una baja concentración de los metabolitos inhibidores del crecimiento del mismo, o por ser estos poco difusibles en el medio, aspecto que se debe esclarecer en investigaciones futuras.
Los resultados de este estudio corroboran los notificados por Reyes et al. (7) y Martínez et al. (12), quienes demostraron la reducción del crecimiento de hongos fitopatógenos por el efecto de metabolitos no volátiles segregados por Trichoderma al medio de cultivo.
Competencia por espacio
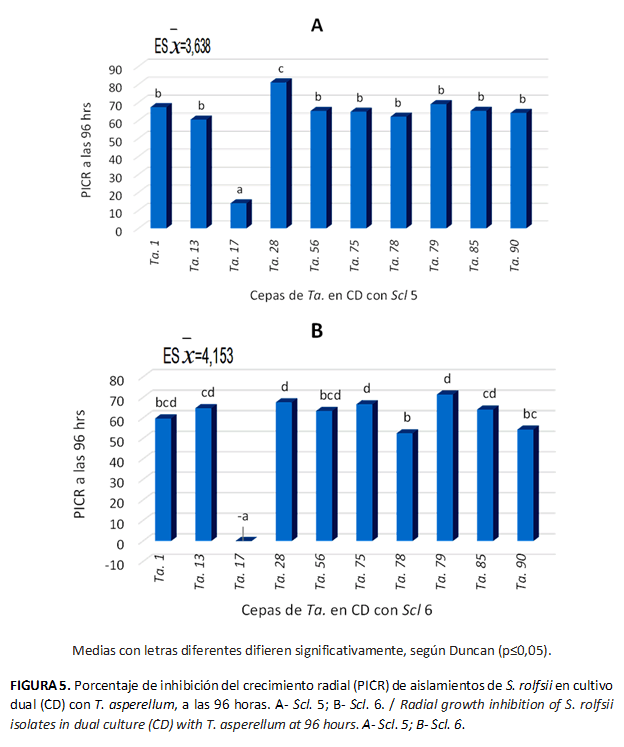
La mayoría de las cepas de T. asperellum evaluadas (Ta. 1, Ta. 13, Ta. 17, Ta. 28, Ta. 56, Ta. 75, Ta. 78, Ta. 79, Ta. 85, Ta. 90) a las 96 h, frente a los dos aislamientos del patógeno, se ubicaron en el grado 2 de la escala de Bell et al. (11). La excepción fue la cepa Ta. 17, que mostró características representativas del grado 5 de la escala (Fig. 3).
El PICR de los aislados Scl. 5 y Scl. 6 varió entre 62,3-81,1 y 52,48-71,42 %, respectivamente. Resultados similares obtuvieron Aceves et al. (5) con cepas de Trichoderma spp. frente a S. rolfsii aislado de cacahuete (Arachis hypogaea L.), con 81,1 % de inhibición del crecimiento micelial del agente fitopatógeno.
Aunque la inhibición del crecimiento de los agentes fitopatógenos está relacionada directamente con la potencialidad de los aislados del antagonista, en el presente estudio se observó un menor efecto de estos sobre Scl 6, lo que evidenció una menor sensibilidad de este aislado a la acción de los antagonistas probados. La inhibición del crecimiento de las colonias de los aislados de los agentes fitopatógenos en estudio coincidió con los resultados informados por Guédez et al. (6), los que justificaron con la mayor velocidad de crecimiento de T. harzianum frente a S. rolfsii.
Reyes et al. (7) y Pino (13) notificaron que una de las características significativas de Trichoderma es su elevada velocidad de crecimiento, lo que le brinda la posibilidad de ser un buen competidor por espacio y nutrientes, fundamentalmente, en condiciones in vitro frente a patógenos causantes de enfermedades en las plantas. La mayor competencia por espacio de las cepas de T. asperellum mencionadas anteriormente, frente a los aislamientos de S. rolfsii, se debió a la elevada velocidad de crecimiento del antagonista, ya que la cepa Ta. 17 (Fig. 3) a las 72 h tuvo un crecimiento mayor que el agente fitopatógeno; sin embargo, a las 120 h el hongo fitopatógeno se desarrolló sobre la colonia de Trichoderma.
Este resultado corroboró lo obtenido por Rollan (14), quien informó que a una temperatura de 30°C, S. rolfsii fue capaz de inhibir el crecimiento de Trichoderma spp. Aceves et al. (5), en un estudio similar, observaron sobrecrecimiento de S. rolfsii sobre uno de los antagonistas (Tcn-7). También Corrêa et al. (15) observaron efecto antagónico nulo de cepas de T. harzianum y Trichoderma aureoviride Rifai frente a S. rolfsii que permitió el crecimiento del patógeno sobre ellas.
Es evidente que, además de la velocidad de crecimiento del antagonista, en este mecanismo intervengan otros elementos, probablemente de origen bioquímico, decisivos para frenar el crecimiento del patógeno.
La mayor parte de los antagonistas, aunque se ubicaron en el grado 2 de la escala, solo detuvieron el desarrollo del patógeno al crecer a su alrededor (Fig. 4). A partir de las 120 h, los aislados del fitopatógeno comenzaron a crecer de nuevo en la zona de interacción, más fácilmente observable a las 168 h. Esto podría ser un indicativo de que los aislamientos de S. rolfsii son capaces de liberar sustancias al medio de cultivo al entrar en contacto con el antagonista, que impiden el avance de este y/o desintoxican los metabolitos excretados por Trichoderma, aspectos que se deben continuar investigando.
De forma general, la cepa Ta. 28 tuvo el mayor PICR para ambos aislados del patógeno, con valores diferentes significativamente al resto de las cepas de Trichoderma, respecto Scl. 5, y sin diferencias estadísticas con Ta. 13, Ta. 56, Ta. 75, Ta. 79 y Ta. 85 frente al aislado Scl. 6. (Fig. 5)
Micoparasitismo
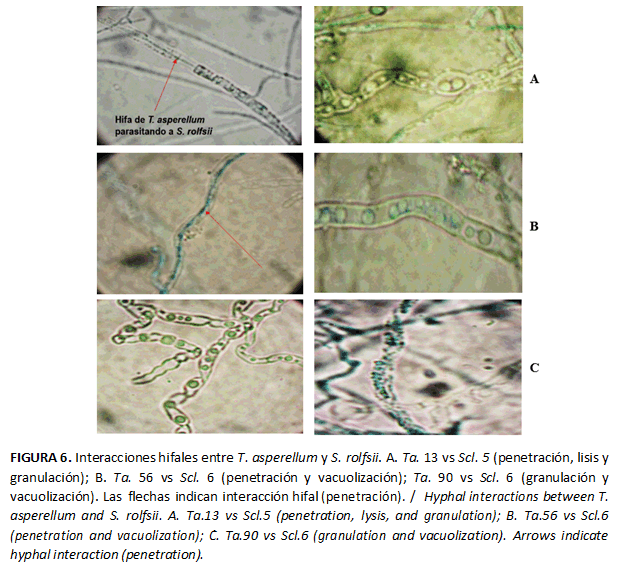
Las observaciones microscópicas de los fragmentos de micelios de la zona de contacto hifal, entre las cepas del antagonista y S. rolfsii en el CD mostraron, al menos, un tipo de interacción hifal definido (Tabla 1).
Todas las cepas del antagonista ocasionaron granulación del contenido citoplasmático en el micelio de los patógenos (Tabla 1). La mejor cepa fue Ta. 90, con tres tipos de interacciones hifales frente al agente fitopatógeno, seguido de Ta. 13 y Ta. 56 con tres tipos de interacción frente a los aislados del fitopatógeno (Scl. 5 y Scl. 6), indistintamente (Fig. 6). Las cepas Ta. 28 y Ta. 78 poseen interés práctico, pues penetraron las hifas de ambos aislados del patógeno.
Diferentes autores notificaron estas interacciones de Trichoderma como un mecanismo antagónico eficaz para el control de hongos fitopatógenos, las cuales tuvieron similitud con las observadas en este estudio. León et al. (16) detectaron micoparasitismo de dos aislados de Trichoderma spp. (Tb111 y Tc241) frente a S. rolfsii con la presencia de estrangulamiento, degradación y lisis de las paredes de las hifas del fitopatógeno. Asimismo, Sandoval et al. (17) observaron hifas de T. harzianum y Trichoderma pseudokoningii Rifai parasitando el micelio de Phytophthora capsici Leonian y Phytophthora nicotianae Breda de Haan.
Al integrar los resultados de los diferentes modos de acción evaluados, se concluye que las cepas de T. asperellum Ta. 13, Ta. 28 y Ta. 90 resultan promisorias para el control biológico de S. rolfsii en frijol. Estos resultados son la base para la determinación de la eficacia técnica de las mismas y para el establecimiento de indicadores como son la dosis, la formulación, el modo, el momento y la frecuencia de aplicación en condiciones de campo.
1. Hernández CA, Quintero E, Herrera L. Respuesta varietal y pérdidas causadas por el hongo Sclerotium rolfsii en frijol común en diferentes épocas de siembra. Centro Agrícola. 2012; 39(1): 75-78.
2. Granados MM. Pudrición blanca de la cebolla: una enfermedad difícil de combatir. Agronomía Costarricense. 2005; 29(2): 143-156.
3. Díaz M, Bernal A, Delgado M. Afectaciones por hongos del suelo en el cultivo del frijol común, en época temprana. Centro Agrícola. 2007; 34(3): 35-38.
4. Rodrigues KP. Avaliação de Trichoderma e de fosfito no controle de Sclerotium rolfsii agente da mucrcha de esclerócio em feijoeiro. 2012. Tesis de maestría en Agronomía. [Consultado: 26 de abril de 2017]. Disponible en: http://repositorio.unb.br/bitstream/10482/11184/1/2012_KleniaRodriguesPacheco.pdf
5. Michel-Aceves AC, Otero-Sánchez MA, Ariza-Flores R, Barrios-Ayala A, Alarcón-Cruz N. Eficiencia biológica de cepas nativas de Trichoderma spp., en el control de Sclerotium rolfsii Sacc., en cacahuate. Avances en Investigación Agropecuaria. 2013. 17(3): 89-107.
6. Guédez C, Cañizalez L, Castillo C, Olivar R. Evaluación in vitro de aislamientos de Trichoderma harzianum para el control de Rhizoctonia solani, Sclerotium rolfsii y Fusarium oxysporum en plantas de tomate. Revista de la Sociedad Venezolana de Microbiología. 2012; 32:44-49.7.
7. Reyes Y, Martínez B, Infante D. Evaluación de la actividad antagónica de trece aislamientos de Trichoderma spp. sobre Rhizoctonia sp. Rev. Protección Veg. 2008; 23(2):112-117.
8. García S, Moya J, Avilés E, Andújar F, Núñez P. Efectividad de Trichoderma spp. sobre el crecimiento micelial de fitopatógenos de suelo en plato Petri. Revista APF. 2015; 4(2): 43-66.
9. Infante D, Reyes Y, Gonzalez N, Martínez B. Molecular identification of thirteen isolates of Trichoderma spp. and evaluation of their patogenicity towards Rhizoctonia solani Kühn. Biotecnología aplicada, 2013; 30: 23-28.
10. Martínez B, Solano T. Antagonismo de Trichoderma spp. frente a Alternaria solani (Ellisy Martin) Jones y Grout. Rev. Protección Veg.1995; 10 (3):221-225.
11. Bell DK, Wells HD, Markam CR. In vitro antagonism of Trichoderma spp. against six fungal pathogens. Phytopathology. 1982; 72:379-382.
12. Martínez B, Infante D, Reyes Y. Trichoderma spp. y su función en el control de plagas en los cultivos. Rev. Protección Veg. 2013; 28(1):1-11.
13. Fernández RJ, Suárez CL. Antagonismo in vitro de Trichoderma harzianum Rifai sobre Fusarium oxysporum Schlecht f. sp passiflorae en maracuyá (Passiflora edulis Sims var. flavicarpa) del municipio zona bananera colombiana. Rev. Fac. Nal. Agr. Medellín. 2009; 62(1): 4743-4748.
14. Rollan M, Mónaco C, Nico A. Efecto de la temperatura sobre la interacción in vitro de especies de Trichoderma y Sclerotinia sclerotiorum, S. minor y Sclerotium rolfsii. Invest. Agr. Prod. Prot. Veg. 1999; 14(1-2):33-48.
15. Corrêa S, Mello M, Ávila Z, Minaré L, Pádua R, Gomes D. Cepas de Trichoderma spp. para el control biológico de Sclerotium rolfsii Sacc. Fitosanidad. 2007; 11(1):3-9.
16. León R, Pino SC, Núñez DB. Aislamiento y selección de cepas de Trichoderma y su efecto antagonista frente a Sclerotium rolfsii, Rhizoctonia sp. y Fusarium sp. Centro Agrícola. 2012; 39(2):43-48.
17. Sandoval I, López MO. Hiperparasitismo de Trichoderma harzianum, T. viride y T. pseudokoningii sobre diferentes hongos fitopatógenos. Fitosanidad. 2001;5(1):41-43.
Recibido: 4/5/2017
Aceptado: 28/9/2017
*Autor para correspondencia: Yanisia Duarte-Leal. E-mail: yanisia@censa.edu.cu


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}