










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión On-line ISSN 2224-4697
Rev. Protección Veg. vol.33 no.1 La Habana ene.-abr. 2018
ARTÍCULO ORIGINAL
Development and prey preference of Macrolophus basicornis (Hemiptera: Miridae) feeding on Myzus persicae and Macrosiphum euphorbiae (Hemiptera: Aphididae)
Desarrollo y preferencia de Macrolophus basicornis (Hemiptera: Miridae) frente a las presas Myzus persicae and Macrosiphum euphorbiae (Hemiptera: Aphididae)
Heyker L. Baños-Díaz*, María de los Ángeles Martinez-Rivero
Dirección de Sanidad Vegetal. Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas. Provincia Mayabeque. Cuba.
ABSTRACT
Biological characteristics, consumption, and prey preference of the predatory bug Macrolophus basicornis preying on Myzus persicae or Macrosiphum euphorbiae on tomato plants were studied. Incubation period of the predator´s eggs was of 8 days and its nymphs presented five instars regardless of the prey consumed. All the nymphal stages showed a similar development time with the exception of the 2nd instar. There were no significant differences between the size and weight of M. basicornis nymphs feeding on M. persicae or on M. euphorbiae. The average rate of aphid consumption revealed significant differences between the 5th instar nymph and the females of the predator when they had no option to choose the prey. In addition, the fifth nymphal stage consumed significantly more aphids in 24 h than the other stages. The females had a high rate of predation comparable to the fifth nymphal stage. M. basicornis was shown to be capable of completing its cycle on preys like Myzus persicae or Macrosiphum euphorbiae. The biological characteristics and prey consumption by M. basicornis were strongly influenced by the prey species and size, the latter being able to influence the prey preference of the predator.
Key words: aphids,biological characteristics, consumption, Macrolophus basicornis, prey preference
RESUMEN
Se estudiaron las características biológicas, el consumo y la preferencia de presa del depredador Macrolophus basicornis usando como presas Myzus persicae y Macrosiphum euphorbiae, que se alimentan de plantas de tomate bajo condiciones controladas. El tiempo de incubación de los huevos del depredador fue de ocho días y las ninfas de M. basicornis presentaron cinco instares sin importar la presa consumida. Todos los estadios ninfales mostraron un tiempo de desarrollo similar, con excepción del segundo instar. No existen diferencias significativas entre las tallas y el peso de las ninfas de M. basicornis que se alimentaron con M. persicae o las que fueron alimentadas con M. euphorbiae. La tasa media de consumo de áfidos reveló que hay diferencias significativas para el quinto instar ninfal y las hembras del depredador cuando no tienen opción de escoger la presa. Además, el quinto estadio ninfal consumió significativamente más áfidos en 24 h que el resto. Los machos tuvieron una alta tasa de depredación en comparación con la hembra y baja en comparación con el quinto estadio ninfal. Los resultados muestran que el depredador M. basicornis es capaz de completar su ciclo con presas como Myzus persicae y Macrosiphum euphorbiae. Las características biológicas y el consumo de M. basicornis están fuertemente influenciadas por la especie presa, el tamaño y la presa; esta última pudiera influir en la preferencia de presa del depredador.
Palabras clave: áfidos,características biológicas, consumo, Macrolophus basicornis, preferencia de presa.
INTRODUCTION
Species of the genus Macrolophus Fieber (Hemiptera: Miridae) are polyphagous predators that are considered effective as biological control agents for many greenhouse pests, such as thrips, aphids, mites, and leaf miners, and on eggs of some lepidopteran species (1). They have been successfully used in temperate and Mediterranean Europe for augmentative biological control and many of them marketed for controlling arthropod pests in greenhouses (2).
Some investigations on the occurrence, abundance, and effect of various parasitoids and predators on tomato and other crops under greenhouse and field environments have shown the substantial importance of the species within the genus Macrolophus (3).
Some species, such as Macrolophus pygmaeus (Rambur), can successfully develop on Myzus persicae (Sulzer) and Macrosiphum euphorbiae (Thomas) (Hemiptera: Aphididae), which are important pests of eggplant (Solanum melongena L.) and tomato (Solanum lycopersicum L). (4)
In Cuba, two species of this genus, Macrolophus basicornis (Stal) (5) and Macrolophus preclarus (Distant), have been reported associated with lepidopteran species and aphids in tobacco (Nicotiana tabacum L.). Nesidiocoris tenuis (Dist) and Engytatus varians (Distant) (syn. Cyrtopeltis varians) have been other plant bug species also found (6).
At present, the use of natural enemies as biological control agents, especially the use of predators of this genus, have been booming (7,8,9) due to the increase of the occurrence of resistance to pesticides and the interest in environmentally sustainable agriculture. However, before polyphagous predators can be used effectively as biological control agents, it is important to obtain a robust information on their biology, predation rate, and prey preference. The latter item is relevant to the amount of energy obtained concerning search costs and consumption of prey (10). All these limitations are closely associated with the prey size and behavior.
Considering the above information, we found important to determine the biological characteristics and the consumption and prey preference of Macrolophus basicornis feeding on M. persicae and M. euphorbiae on tomato plants.
MATERIAL AND METHODS
Collection of specimens
Adult insects were collected from an area with tomato in Lavras (21° 14. 45' S, 44° 59.59' W, 920 m altitude), and from another area with tobacco in Ribeirão Vermelho (21° 08,596' S and 045° 03,466' W, 808 m altitude) in June and August 2012. The adults were captured with the aid of an exhauster and placed in glass test tubes (2.5 x 8.0 cm), sealed with PVC plastic film, and carried to the Biological Control Laboratory, Department of Entomology, Federal University of Lavras, Minas Gerais.
Prey rearing
The preys were reared on a leaf of Nicandra physalodes (L.) Gaertn, for M. persicae, and of Lectuca sativa. var Veronica, for M. euphorbiae, in 15 cm Petri dishes with a layer of 1c % water agar (5 dishes/sp.). In both cases, 20 aphid adults of each species were placed on each leaf. The dishes were covered with a paper towel with a rubber band and placed into a controlled climatic chamber (BIOD) at 21 ± 1 ◦C, RH 70 ± 10 % and photoperiod of 12 hours; the plates were changed twice a week.
The leaves for the aphid rearing were disinfected with a solution of 1 % sodium hypochlorite for 5 min, washed with tap water, and finally rinsed with distilled water for 10 min. This guaranteed high quality lettuce and nicandra leaves.
Predator rearing. Once in the laboratory, the adult predators were released in acrylic cages (30x30x60 cm) containing Nicotiana tabacum L. plants with approximately 25 cm of height, which were used as the oviposition substrate. The predators were fed with eggs of Anagasta kuehniella (Zeller) ad libitun. The climatic conditions for the rearing were establish a temperature of 25 ± 2 °C, a relative humidity of 70 ± 10 %, and a photoperiod of 12 hours.
The plants with the adult predators remained in their cages for seven days. Afterward, plants with new eggs were placed in cages. Once the nymphs were born, eggs of A. kuehniella were offered twice a week until the emergence of new adults.
Egg embryonic development of the predator Macrolophus basicornis
To obtain M. basicornis eggs, ten couples of the predator were placed in glass containers (1,7 liters) containing a small tomato plant (Solanum lycopersicum L). cv. Santa Clara with two pairs of leaves, and a mixture of 1st to 3rd instar nymphs of M. persicae or M. euphorbiae was used as food. The glass containers were sealed with a voile-type fabric and kept in a room at 28 ± 2 ºC, RH 70 ± 10 %, and photoperiod of 12 hours. After 24 hours and every day until the death of adults, the plants with eggs were replaced by new ones, and the number of eggs counted under a stereo microscope. Finally, the roots of these plants were wrapped with cotton, placed in Petri dishes (15 cm), and sealed with a PVC film.
The embryonic period length was evaluated by observations made on egg development and survival; the number of eggs and percentage of fertile eggs were also taken into account. The female ovipositional period was estimated from the first to the last oviposition day.
Development and reproduction of M. basicornis preying on M. persicae and M. euphorbiae on tomato plants
Plants of tomato cv. Santa Clara with approximately 25 cm of height were placed into a rearing cage of M. basicornis. After 24 hours, they were moved to a rearing cage and placed into a controlled climatic chamber (BIOD) at 28 ± 1 ◦C, RH 70 ± 10 % and a l2 hour photoperiod. They were examined daily until appearance of nymphs. Then, 100 newly hatched nymphs were individually placed on 5 cm discs of S. lycopersicum leaves located on a 1 % water agar layer in 5 cm dishes. They were changed twice a week. The nymphs were daily fed with five 1st instar nymphs of M. persicae or M. euphorbiae under the same previously described conditions. They were daily examined for instar changes indicated by the presence of exuvias. Duration time of each instar, nymph mortality, and sex ratio were recorded.
Once nymphs turned into adults, 20 couples of 24 hour- old M. basicornis were kept in glass jars (1.7 L) containing a small tomato plant with three leave as ovipositional substrate under the same conditions in which the nymphs were born. Each predator couple were fed daily with 10 nymphs of M. persicae or M. euphorbiae. The oviposition substrate was changed daily, and the plants placed in 15 cm Petri dishes with the roots covered with moistened cotton until nymph emergence.
The number of eggs layed, mortality of adults, and number of nymphs emerged were recorded daily. From the data, pre-oviposition and oviposition periods, fecundity (daily and total eggs / female) and longevity were attained.
Measuring and weighing M. basicornis
The size of 20 M. basicornis nymphs from the 1st to the 5th instar were determined. They were measured with a reticle coupled to the micrometrical stereomicroscope (Olympus SZ40). The width of the head (head capsule) (distance between the outer edges of the compounds eyes) was also measured.
Twenty 4th and 5th instar nymphs were weighed individually using an analytical balance (Shimadzu, AW 220). They were weighed one day after each instar change.
Assessment of the consumption capacity of M. basicornis predation feeding on M. persicae or M. euphorbiaeon tomato plants
The foraging arena consisted of a leaf of S. lycopersicum cv. Santa Clara with mixed nymphs from the 1st to the 4th instar of M. persicae and M. euphorbiae placed on a 1 % water agar layer in a 5 cm Petri dish
Without chance to choose. One predator female up to one day old and one 5th instar predator nymph were collected from the laboratory maintenance predator culture and kept individually in a glass tube with moistened cotton and without access to food for 24 hours prior to the experiments. Then, they were released into 9 cm Petri dishes each containing 10 mixed instar nymphs of M. persicae or M. euphorbiae. After 24 hours, the consumed preys were determined.
With chance to choose. In this case the methodology above described was followed, but 40 mixed instar nymphs were placed on this occasion, that is 20 of each prey, M. persicae and M. euphorbiae. After 24 hours the number of consumed preys were determined.
The trials were conducted in a controlled climatic chamber (BIOD) at 28± 2 ºC, 70 ±10 % RH and 12h photophase.
The number of aphids consumed per 24h were used to evaluate the predator’s preference index determined by Manly´s index, which considers the depletion of prey due to predation (12). Manly´s index is given by:
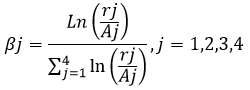
where:
βj - Manly´s Beta for aphid instar j
rj - the number of individuals in aphid instar j not select by the predator
Aj - the number of individuals in aphid instar j available to the predator
Statistical Analysis. All the data were analyzed using Simple ANOVA. The means were compared using the post hoc Tukey test (P≤0.01). The statistical analyses were conducted using the statistical software package InfosStat version 2.0 (11).
RESULTS AND DISCUSION
Nymphal development
Like all predators ofthe genus Macrolophus, Macrolophusbasicornis has a zoophytophagous feeding behavior. This means that he can feed on both, plant and prey. This behavior could be observed during the experimental work due to the appearance of necrotic rings in the feeding areas similar to those reported by Urbaneja et al. (2).
Regardless of the prey supplied, the predator egg incubation period on tomato plants was eight days. Similarly, the predator post-embryonic development included five nymphal stages feeding on each prey, each stage with a maximum duration of eight days. However, when M. persicae was supplied as a prey, an extra day was added to predator´s fifth nymphal stage. Nevertheless, when M. euphorbiae was supplied as a prey, the predator´s first nymphal stage reached eight days, consequently resulting in a longer nymphal development. With both preys, the stadiums of interest (fourth and fifth) were achieved in three days. The development and life cycles of the predator showed significant differences with both preys, being longer when M. euphorbiae was the prey. In both cases, the life cycle duration was longer than one month. Therefore, considering this time and depending on the prey consumed, several annual predator generations per year can be possible, ranging from ten (on M. persicae) to seven (on M. euphorbiae). (Table 1)
Regarding the sexual coefficient, it was always favourable to females regardless of the prey supplied. This is a very convenient fact because of their role for preserving the species. They also had a higher rate of consumption. Females and males reached adulthood at the same time, so there were no difference in the period of development between the sexes. The mean longevity of the adult predator feeding on M. persicae was 8 days, however longevity of those adults kept feeding on M. euphorbiae was longer (25 days). This differences could be caused by the quality and size of the prey offered, maybe along with the predators prey preference.
The sex ratio always proved favorable to females regardless of the prey supplied, although, as Table 1 shows, it was slightly higher when the predator fed on M. euphorbiae. Females and males reached adulthood at the same time, so there was no difference in the period of development between the sexes.
The prey species also influenced on the reproductive parameters of M. basicornis. A higher oviposition period and daily fecundity of M. basicornis were observed when the prey was M. euphorbiae than when the prey was M. persicae. The predator’s total fecundity showed the same behavior with an increase of ten eggs when it fed on M. euphorbiae (Table 2). These results could corroborate the above proposed influence of prey quality, size and predator´s preference on its development and reproduction.

Not much information on development and reproduction of this species preying aphids is available, but there is a lot on other species in the same genus. For example, Hatherly et al. (13) observed a complete development of M. caliginosus on M. persicae on different host plants. The predator sex ratio varied from 0.44 on Chinesse cabage to 0.47 on oil seed rape, and mortality lower than 50 % in both case. However, on each crop, the oviposition period was 5 and 10 days, the progenies per female were 45 and 20 nymphs/female with a survival of 60 and 65 %.
In tobacco, the development period of the predator M. caliginosus feeding on M. persicae was 17 days with 94 % of nymphal survival (14). On the other hand, Alvarado et al. (15) evaluated A. gossypii on cucumber and M. euphorbiae on tomato for their suitability as prey items for the development of M. caliginosus nymphs. Development of nymphs was shorter with M. euphorbiae on tomato than with A. gossypii on cucumber.
The results obtained with both preys studied in the present work differed from those achieved by other authors. The development period was longer than in other species such as M. caliginosus and M. pygmaeus. The differences between the nymphal development, sex ratio, fecundity and the rest of the biological characteristics could be influenced by the host plant and the prey. For instance, the incubation period could be influenced by the type and quality of the prey offered. Prey quality could affect the number of eggs laid (16). According to Urbaneja et al., (17), the host plant is another factor influencing development period and survival.
In species such as M. caliginosus, any testing of host range with the proposal of informing a risk assessment of its establishment potential is further complicated by the zoophytophagous nature of the predator; that is, the capability of the insect to feed on both the plant and arthropod prey (18). In such cases, it is necessary to assess the survival, development, and reproduction of the species in the absence of any prey.
In the case of M. basicornis, the offered preys were low quality foods. There was not marked influence on weight in none of the instars, even when the size varied slightly in the 5th instar with M. euphorbiae as the prey (Table 3). Despite no significant differences were observed in the nymph size of M. basicornis feeding on M. persicae or M. euphorbiae, there were significant differences in the weight of the 5th instar nymphs, which were heavier when M. euphorbiae was the prey.
Aphid consumption
Usually, prey suitability and the innate prey preference of a predator are correlated because natural selection acts to optimize fitness (19). Additionally, if the optimal prey diet maximizes the predator’s net energy gain, the prey will be either accepted or rejected based on whether it fits within this optimal diet (20). Consequently, predators tend to be more selective in environments with high prey availability and less selective in environments where prey is scarce (21).
Observations on the mean aphid consumption rate revealed significant differences in both prey species for 5th instar nymphs and females when they were offered separately; however, no significant differences were observed when both species were offered together. In addition, the 5th instar nymphal stage significantly consumed more aphids in 24 h than the other stages. Females had a higher predation rate compared with the 5th instar nymphal stage. Comparing the two prey species, the 5th instar nymph of M. basicornis consumed more nymphs of M. persicae than of M.euphorbiae (18.96 and 15.08 nymphs in 24 hours); however, the M. basicornis female consumed more nymphs of the aphid M. euphorbiae. (Table 4)


When the two prey species where offered together, no significant differences were found between the 5th instar nymph and the female of the predator, neither between Manly´s preference index for each prey species in any stage evaluated or between stages.
Eubanks and Denno (22) suggest that not always a prey may be preferred. Prey size is another possible criteria involved in predator selection, and it can play a role in the predator preference under the experimental conditions in which is evaluated. According to Lins (23), the encounter rate can be higher in larger preys.
In similar studies, Perdikis et al., (39) showed that the voracity of females and 5th instar nymphs of the predator M. pygmaeus on nymphs of M. persicae is higher than the voracity of other stages of the predator. The predation rate of M. pygmaeus was higher on M. persicae than on Macrosiphum euphorbiae, whereas its preference depended on the aphid species and the ratio of the available instars.
The predation rate per hour of M. pygmaeus in the light and dark phases was examined by Perdikis et al., (24) in different plants. It was significantly higher in the dark than in the light phase. On the other hand, M. pygmaeus showed a strong preference for small preys. According to this author, the preference showed by M. pygmaeus for particular prey instars might also be valuable in the effort to explore its functional response under more realistic, mixed prey experiments.
Therefore, observed modifications of the predatory behavior in mixed prey assemblages may have considerable consequences for population dynamics and regulatory mechanisms acting in predator–prey natural systems.
For predators, prey preference usually refers to active selection or else to the prey selectivity that is exhibited when they are foraging in mixed prey patches. Prey preference is determined not only by prey characteristics such as size, vulnerability, mobility, and caloric value, but also by predator–prey encounter rates (25).
According to the foraging theory, a predator makes the decision whether to attack an available prey type of low quality or shift to a patch of a more preferable prey type. Correspondingly, when a high quality prey is available, a low quality prey is most likely to be ignored (26). This implies that a predator is able to rank prey types according to their suitability, and that prey selection by preference is based on a mechanism of prey discrimination (11).
The ability to locate and utilize a wild prey is another essential requirement for the long term establishment of such non-native species. However, while the ‘host range’ is regarded as a critical component for rearing nonnative biocontrol agents, there is not a commonly agreed methodology as yet, and it is recognized that such methods, as proposed (27), require further refinement.
Our results are very similar to those obtained by other authors. M. basicornis, as the other Macrolophus spp., showed a strong preference for M. persicae. Prey preference can be influenced by the host plant on which the prey was feeding or their volatile emissions. The biological characteristics and the consumption of M. basicornis feeding on different preys are strongly influenced by the prey species and size and the prey host plant, and the latter can influence prey preference of M. basicornis.
ACKNOWLEDGEMENTS
Thanks are due to Elaine Louzada and Nazared Moura for their technical assistance during the experimental work and to Professor Vanda Elena Páez Bueno for the support in the experimental phase. CAPES agency is gratefully acknowledged for the financial support of this research.
This work was supported by the Coordenação de aperfeiçoamento de pessoal de nível superior – CAPES under Grant 118/11
1. Lykouressis DP, Perdikis DCh, Chalkia ChA. The effects of natural enemies on aphids populations on processing tomato. Entomol. Hellenica 13(1999–2000): 35–42.
2. Urbaneja A, Monton H, Molla O. Suitability of the tomato borer Tuta absoluta as prey for Macrolophus pygmaeus and Nesidiocoris tenuis. Journal of Applied Entomology. 2009;133: 292–296.
3. Sampson A, King V. Macrolophus caliginosus, field establishment and pest control effect in protected tomatoes. Bull. IOBC/WPRS 1996; 19(1): 143–146.
4. Lykouressis D, Perdikis D, Tsagarakis A. Polyphagous mirids in Greece: Host plants and abundance in traps placed in some crops. Boll. Lab. Entomol. Agr. Fillippo Silvestri 2000; 56: 57–68.
5. Grillo H. Heterópteros de Cuba. [Tesis presentada en opción al título de Doctor en Ciencias]. Universidad Central de las Villas, Cuba. 2012.
6. Martínez MA, Duarte L, Baños HL, Rivas A, Sánchez A. Predatory mirids (Hemiptera: Heteroptera: Miridae) in tomato and tobacco in Cuba. Rev. Protección Veg. 2014; 29(3): 204-207.
7. Machtelinckx T, Van Leeuwen T, Van De Wiele T, Boon N, De Vos W, Sánchez JA, et al. Microbial community of predatory bugs of the genus Macrolophus (Hemiptera: Miridae). BMC Microbiology. 2012;12(Suppl 1):S9. http://www.biomedcentral.com/1471-2180/12/S1/S9.
8. Bueno VHP, van Lenteren JC, Lins JC, Calixto AM, Montes FC, Silva DB, Santiago LD, Perez LM. New records of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) predation by Brazilian Hemipteran predatory bugs. J. Appl. Entomol. 2012; 137:29-34
9. Krebs JR , Mccleery RH. Optimizations in behavioural ecology. In Krebs J.R. & Davies N.B. (eds): Behavioural Ecology, an Evolutionary Approach. Blackwell Scientific, Oxford, 1984:91–121.
10. Di Rienzo JA, Casanoves F, Balzarini MG, Gonzalez L, Tablada M, Robledo CW. InfoStat versión 2016. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. URL http://www.infostat.com.ar
11. Chesson J. The estimation and analysis of preference and its relationship to foraging models. Ecology. 1983; 64: 1297–1304.
12. Hatherly IS, Pedersen ABP, Bale AJS. Effect of host plant, prey species and intergenerational changes on the prey preferences of the predatory mired Macrolophus caliginous. BioControl. 2009; 54:35–45
13. Vandekerkhove B, De Puysseleyr V, Bonte M, De Clercq P. Fitness and predation potential of Macrolophus pygmaeus reared under artificial conditions. Insect Science. 2011; 18, 682–688.
14. Alvarado P, Baltà O, Alomar O. Efficiency of four heteroptera as predators of Aphis gossypii and Macrosiphum euphorbiae (Hom.: Aphididae). Entomophaga. 1997; 42 (1–2):215–226
15. Hamdi F, Chadoeuf J, Bonato O. Functional relationships between plant feeding and prey feeding for a zoophytophagous bug. Physiol Entomol 2013; 38(3): 241-245.
16. Urbaneja A, Tapia G, Stansly P. Influence of host plant and prey availability on developmental time and surviorship of Nesidiocoris tenius (Het.: Miridae), Biocontrol Sci. Technol. 2005; 15 (5): 513-518.
17. Naranjo SE, Gibson RL. Phytophagy in predaceous Heteroptera: effects on life history and population dynamics. Proceedings Tomas Say Publications in Entomology. 1996:57-96
18. Dicke M, Sabelis MW, Takabayashi J, Bruin J, Posthumus MA. Plant strategies of manipulating predator–prey interactions through allelochemicals: prospects for application in pest control. Journal of Chemical Ecology. 1990; 16 3091– 3118.
19. Pyke GH, Pullian HR, Charnov EL. Optimal foraging: a selective review of theory and tests. Quarterly Review of Biology. 1977; 52:137–154.
20. Williams DD. A laboratory study of predator–prey interactions of stoneflies and mayflies. Freshwater Biology. 1987; 17: 471–490
21. Eubanks MD, Denno RF. Health food versus fast food: the effects of prey quality and mobility on prey selection by a generalist predator and indirect interactions among prey species. Ecol Entomol. 2000; 25(2):140-146
22. Lins Junior JC. Search capacity, prey preference, predation rates and reaction to prey and predator induced volatiles of predatory mirids of two tomato pests, Tuta absoluta (Lep.: Gelechiidae) and Bemisia tabaci (Hem.: Aleyrodidae). [Tese (doutorado)], Universidade Federal de Lavras, Lavras. 2014:116pp.
23. Perdikis DCh, Lykouressis DP, Economou LP. The influence of temperature, photoperiod and plant type on the predation rate of Macrolophus pygmaeus on Myzus persicae. BioControl. 1999; 44: 281–289.
24. Perdikis DCh, Lykouressis DP. Life table and biological characteristics of Macrolophus pygmaeus when feeding on Myzus persicae and Trialeurodes vaporariorum. Entomologia Experimentalis et Applicata.2002;102: 261–272.
25. Cohen AC, Tang R. Relative prey weight influences handling time and extracted biomass in predatory hemipterans. Environmental Entomology. 1997; 26:559–565.
26. Tschanz B, Bersier LF, Bacher S. Functional responses: a question of alternative prey and predator density. Ecology. 2007; 88:1300–1308.
27. van Lenteren JC, Babendreier D, Bigler F, Burgio G, Hokkanen HMT, Kuske S, Loomans AJM, Menzler-Hokkanen I, Rijn van PCJ, Thomas MB, Tomassini MC, Zeng QQ. Environmental risk assessment of exotic natural enemies used in inundative biological control. BioControl. 2003; 48: 3–38.
Recibido:6/9/2017
Aceptado: 2/2/2018
*Autor para correspondencia: Heyker L. Baños-Díaz. E-mail: hlellani@censa.edu.cu
"This work was supported by the Coordenação de aperfeiçoamento de pessoal de nível superior – CAPES under Grant 118/11."


{kind=link}
{kind=link}