










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión On-line ISSN 2224-4697
Rev. Protección Veg. vol.33 no.1 La Habana ene.-abr. 2018
ARTÍCULO ORIGINAL
Characterization of Cuban native bacteria isolated from nematodes as potential biological control agents for Meloidogyne spp.
Caracterización de bacterias aisladas a partir de nematodos como controladores biológicos potenciales de Meloidogyne spp.
Ileana Sánchez Ortiz1*, Irene Alvarez Lugo1, Idania Wong Padilla1, Danalay Somontes1, Rosa Basulto Morales1, Rolando Morán Valdivia1, Eulogio Pimentel Vázquez2, Jesús Mena Campos2
1Center of Genetic Engineering and Biotechnology, Camagüey. Circunvalación Norte. Zip: 70100. Camagüey, Cuba.
2Center of Genetic Engineering and Biotechnology. Ave. 31 e/158 y 190, Cubanacán, Playa, P.O. Box 6072, Havana, Cuba.
ABSTRACT
The objectives of this study were the molecular and conventional characterization of three native strains isolated from parasitic nematodes, evaluation of their potential to control Meloidogyne spp., and determination of their possible pathogenicity traits against nematodes. The identity of the strains Bacillus thuringiensis CIGBR23, Sphingobacterium sp. CIGBTb, and Stenotrophomonasmaltophilia CIGBG1 was confirmed by 16S rRNA sequencing. Their effectiveness to reduce infestation of Meloidogyne spp. was evaluated in Cucurbita maxima RG-T150 as host plant growing in pot. The plant growth-promoting effects of the strains were also determined. The root-galling index, branch length, and branch and root fresh weights were determined 35 days after nematode inoculation. The three strains controlled Meloidogyne spp. The treatment with Sphingobacterium sp.CIGBTbwasthe most effective, reducing the infestation index from 3 to 1; whereas it was 1.6 for B. thuringiensis CIGBR23 and 1.7 for S.maltophilia CIGBG1 (Hussey and Janssen’s scale 0 -5). Sphingobacterium sp.CIGBTb and S.maltophilia CIGBG1 also significantly reduced the number of root galls by C. maxima (higher than 50 %)in relation to the control (p˂0.05). On the other hand, the treatment with B. thuringiensis CIGBR23 increased plant weight in 17 %. The three strains produced chitinase enzymes. Two of the strains (CIGBR23 and CIGBG1) also excreted lipases and proteases, and CIGBG1, in addition to these enzymes, also produced phospholipase and hydrogen sulfide.
Key words:Bacillus, biological control, root knot nematodes, Sphingobacterium, Stenotrophomonas.
RESUMEN
Los objetivos de este estudio fueron caracterizar, por métodos moleculares y convencionales, tres cepas nativas aisladas de nematodos parasíticos para evaluar sus potencialidades biocontroladoras sobre Meloidogyne spp. y determinar sus posibles atributos de patogenicidad sobre estos organismos. La identidad de las cepas se confirmó mediante secuenciación del ARNr 16S. La efectividad de Bacillus thuringiensis CIGBR23, Sphingobacterium sp. CIGBTb y Stenotrophomonasmaltophilia CIGBG1 para reducir la infestación de Meloidogyne spp. se evaluó en macetas con Cucurbita maxima RG-T150. También se determinó el efecto promotor del crecimiento de las cepas. El índice de agallamiento, la longitud y la masa fresca de las ramas y las raíces se determinaron a los 35 días de inoculación del experimento. Las tres cepas controlaron Meloidogyne spp.. Sphingobacterium sp. CIGBTb fue el más efectivo de los tratamientos y redujo el índice de infestación de 3 a 1; mientras que B. thuringiensis CIGBR23 a 1,6 y S. maltophiliaCIGBG1 a 1,7 (escala 0-5 de Hussey y Janssen). Sphingobacterium sp. CIGBTb y S. maltophilia CIGBG1 también disminuyeron significativamente (mayor que 50 %) el número de nódulos en las raíces de C. maxima (p˂0,05) respecto al control. Además, el tratamiento con B. thuringiensis CIGBR23 aumentó la masa de las plantas en un 17 %. Las tres cepas presentaron enzimas quitinasas, dos (CIGBR23 y CIGBG1) excretaron además lipasas y proteasas; mientras que, CIGBG1 produce también fosfolipasas y sulfuro de hidrógeno.
Palabras clave: Bacillus, control biológico, nematodos agalleros, Sphingobacterium, Stenotrophomonasmaltophilia
INTRODUCTION
Meloidogyne spp.is one of the pests most commonly found on fruits and vegetables grown under protected cultivation. These parasitic nematodes attack the plant roots affecting the utilization of water and nutrients by plants. The disease causes reductions in yield, mainly in tropical and sub-tropical agricultural areas. Yield losses have been estimated to exceed 30 % in vegetables such as eggplant, watermelon and tomato (1). In Cuba, the root knot nematodes (RKN) are also an important phytosanitary issue for the protected system, particularly in tomato and watermelon (2).
Control of nematodes by carbamates and organophosphates is fast and effective (3). However, public awareness on the damage to the environment and the residual effects of chemicals has increased the interest to find safe substitutes for the control of plant-parasitic nematodes. The application of native microbial antagonists as biological agents to control parasitic nematode is a friendly alternative to the ecosystem (4).
The native bacteria used in this work were the strains Bacillus thuringiensis CIGBR23, isolated from juveniles of phytonematodes, and Sphingobacterium sp.CIGBTb and Stenotrophomonasmaltophilia CIGBG1, isolated from eggs of zoonematodes. Previously, they were selected for their in vitro activity against Haemonchus spp.(5). Due to the similar chemical composition of phytonematodes and parasitic zoonematodes (6), the bacterial strains used may be expected to control Meloidogyne spp. infestation in plants.These microbial genera can adapt well to varied environmental conditions (7, 8, 9) and can offer new alternatives to control phytonematodes. Therefore, the aims of this study were the molecular and conventional characterization of three native strains isolated from parasitic nematodes, evaluation of their potential to control Meloidogyne spp., and determination of their possible pathogenicity traits against nematodes.
MATERIALS AND METHODS
Bacteria
B. thuringiensis CIGR23, Sphingobacterium sp. CIGBTb and S.maltophilia CIGBG1, were from the Collection of the Center of Genetic Engineering and Biotechnology (CIGB), Camagüey, Cuba. The strains were grown in Tryptone Soy Broth (TSB) (Oxoid; 30 g l−1) in an orbital shaker at 250 rpm and 30ºC for 24 h. Bacillus thuringiensis CIGR23 was shaken for 48 hours to induce sporulation.
Nematodes
Meloidogyne spp.was collected from cucumber plants growing in protected houses in Ciego de Ávila, Cuba. The population of Meloidogyne spp.was propagated inCucurbita maxima varRG-T150 plants at CIGB Camagüey, Cuba. The egg masses were removed from the roots of C. maxima varRG-T150 with dissecting needles. The eggs were disaggregated with 0.5 % sodium hypochlorite. The egg suspension was firstly sieved through a 60 µm mesh and then through a 30 µm mesh. The eggs retained on the last mesh were dipped in sterile distilled water. They were preserved at 8ºC until the assay was performed. Eggs were counted under an inverted binocular microscope (x40) Olympus CK 2.
Identification of bacteria using 16S rRNA gene sequencing
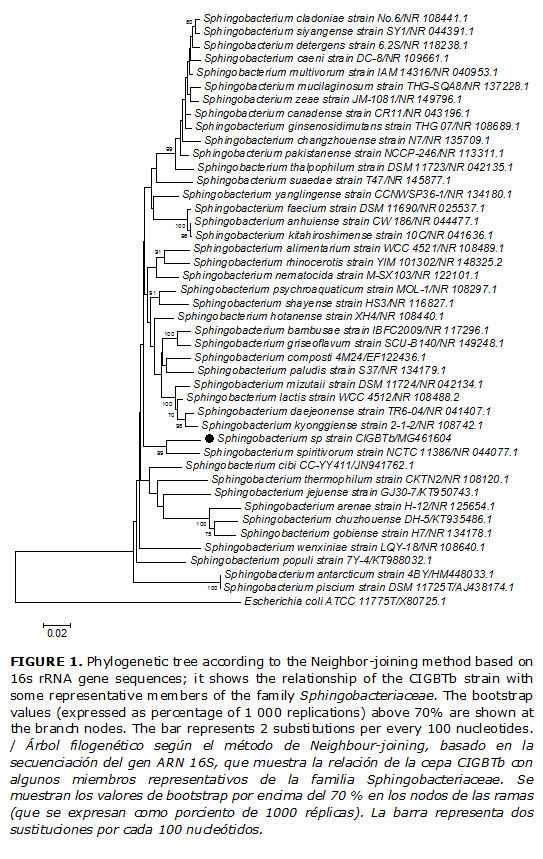
A PCR amplification of the 16S rRNA gene of the strains was performed using universal forward and reverse primers: 27F (5’-AGAGTTTGATC(AC)TGGCTCAG-3’) and 1492R (5’-TACGG(ACT)TACCTTGTTACGACTT-3’). DNA template was prepared by picking an individual colony and dissolving it in 1 X Tris-EDTA buffer solution. The reaction was performed with 5 µl of DNA, 2.5 mM of MgCl2, 100 µM of each dNTP in reaction buffer (50 mM de KCl and 10 mM Tris-HCl pH 8. 24), one unit of DNA polymerase of Thermus aquaticus, and 1 pmol/µl of each primer. Reaction mixture (100 µl) was denatured at 95ºC for 1 min, followed by primer annealing at 55ºC for 1 min and the extension at 72ºC for 1. 5 min. Twenty-five cycles was carried out in a thermocycler. The purified PCR products were sequenced by MACROGEN (Seoul, Korea) with the same primers. The sequences obtained were compared with available sequences retrieved from GenBank using the BLAST program (http://www.ncbi.nlm.nih.gov/blast/) to determine an approximate phylogenetic affiliation. Phylogenetic analysis was performed to the strain CIGBTb using the software packages MEGA version 6.06 after multiple alignment of data using CLUSTAL _ X (10). Evolutionary distances of strain CIGBTb was calculated according to the Kimura two-parameter model (11), and clustering were based on the neighbor-joining (12) method. Bootstrap analysis (1000 replications) was used to evaluate the topology of the neighbor-joining tree (13).
Determination of extracellular enzymes
Chitinase production by the bacterial strains was tested by growing them in M9 medium with colloidal chitin (14). Chitosanase production was corroborated by using chitosanase detection agar plates (15). Other enzymes detected were lipases, by formation of a halo with micelles on nutrient agar medium with 1 % tween 80 (16); phospholipases, by the appearance of opalescence in Nutrient Agar medium with egg yolk at 0.2 % (17); proteases, on plates containing nutrient agar and gelatin at 0.5 % and the subsequent development with Frazier´s reagent (18); phosphatase, esterase, glucoronidases, fucosidase, mannosidase, arylase, N-acetyl-β-glucosaminidase, and Naftol- AS-BI-fosfohydrolase, by using the API ZYM kit. Casein hydrolysis was tested on Nutrient Agar medium with skim milk at 10 % (17).
Production of hydrogen sulfide
Production of hydrogen sulfide was tested by holding a strip of lead acetate paper in the mouth of cotton-wool plugged tubes containing 5 mL of Nutrient Broth with cysteine (0.1 %) and inculated with the bacterial strains. Strip darkening indicated a positive reaction (19).
Detection of cry genes through Polymerase Chain Reaction (PCR)
The following primers were used for cry gene amplification:
cry1:(d)5'-CATGATTCATGCGGCAGATAAAC-3' and
(r)5'-TTGTGACACTTCTGCTTCCCATT-3'
cry3: (d)5'-TAACCGTTATCGCAGAGAAATGA-3'
(r)5'-CATCTGTTGTTTCTGGAGGCAAT-3'
cry7: (d)5'-TTAGGTGTCCCAGGTGCAAG-3'
(r)5'-GGTCTGGTTGAATCTCATGTG-3'
cry8: (d)5'-TTAACAGATAGACTCCAACAAGC-3'
(r)5'-TTTGTGATATAGCTGGTTTGATCA-3'
The reaction mixture contained Taq DNA polymerase 1X buffer (Enzibiot, Heber-Biotec, Cuba); 0.2µM of each oligonucleotide; 0.2 mM dNTPs (Promega); 2.5 mM MgCl2 (Enzibiot, Heber-Biotec, Cuba); 3.0 U Taq DNA polymerase (Enzibiot, Heber-Biotec, Cuba). Amplification was carried out as follows: a first cycle of denaturation (5min; 95°C), hybridation (1 min; 52°C) and extension (1 min; 72°C). It was followed by 35 cycles: denaturation (1min; 93°C), hybridation (1 min; 52°C), and extension (1 min; 72°C). Finally, one last extension (3 min) was made, which allowed completion of the amplified fragments. The results from each reaction were checked on 1 % Agarose gel.
Pot trials
Seed preparation
C. maxima varRG-T150 seeds (10 per treatment) were dipped in 70 % ethanol and washed twice with sterile distilled water. They were placed in sterile Petri dishes, on Whatman filter paper No. 1 (4.25 cm diam.) damped with sterile distilled water, and incubated at 30ºC for 5 days. After germination, and before planting, the seeds in each treatment were dipped in bacterial suspensions (at 105-106 ufc/ml concentrations) for 5 minutes.
Substratumpreparation
Nylon bags (8 cm diameter ×15 cm deep) were filled with 500 cm3 of substratum (3:1 of sterile sand: enriched sterile pit (Terraplant)). The substratum was infested by distributing 500 eggs of Meloidogyne spp.in three different points at 3 cm depthinside the bags.
Experimental design
A completely randomized design with five repetitions of each treatment was used. The treatments were the seed bacterization with: B. thuringiensis CIGBR23, B. thuringiensis CIGBR23 (stationary phase), Sphingobacterium sp. CIGBTb S.maltophilia CIGBG1 (50 ml of culture, at 107-108 ufc/ml concentrations) and the control (50 ml of TSB medium). The previously bacterized seed were planted at 2 cm depth 48 hours after soil infestation with Meloidogyne spp.
Galling index, branch length, branch, and root weights were determined on the 35th day. The galls per grams of root were determined by counting the knots in 5 g of root segments from each treatment. Galling index was ranked as follows: 1 = trace infection with few small galls; 2 = ≤ 25 % of galled roots; 3 = 26 to 50 %; 4 = 51 to 75 %; and 5 = >75 % of galled roots (20).
Data analysis
All calculations were made using Statgraphics plus 5.0. Variance analysis (ANOVA) was made to all the data, and the means were compared according to the Duncan´s test (p<0.05).
RESULTS
According to BLAST search results in NCBI WEB site, the highest similarity (96) of the strain CIGBG1 was with S. maltophilia IAM 12423 (NR_041577.1). CIGBR23 was closely related (97 %) to B. thuringiensis IAM 1207 (NR_043403.1), and CIGBTb was more related to Sphingobacteriumspiritivorum (NR_044077.1) but with only 89 % of similarity. The phylogenetic analysis using the rRNA16S sequences of the type species of the genera Sphingobacterium confirmed that, although the similarity value was less than 97% (95,58 %). It Indicates that CIGBTb belongs to the genus Sphingobacterium, having the highest similarity with S.spiritivorum NCTC 11386 (Fig. 1), but CIGBTb strain does not belong to this species.
All the strains showed nematicidal effects on Meloidogyne spp.. Sphingobacterium sp.CIGBTb had the best behavior, with a reduction of 2 grades of the scale; however, B. thuringiensis CIGBR23, and S.maltophilia CIGBG1 reduced the infestation index in 1 grade (Table 1).
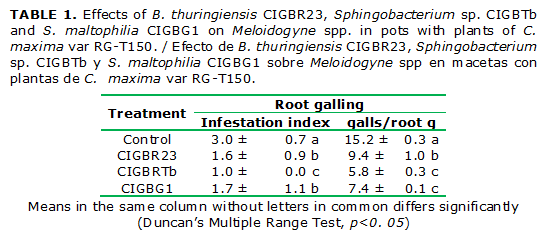
The application of Sphingobacterium sp.CIGBTb and S.maltophilia CIGBG1 significantly (p˂0.05) reduced the number of root galls by more than 50 %, regarding the control. B. thuringiensis CIGBR23 reduce galling in 38, 2 % (Table 1).
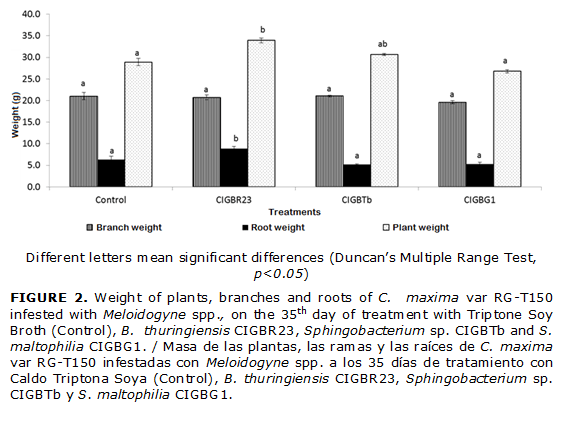
Some microbial antagonists also stimulate plant growth. Plant height and weight are some of the parameters stimulated by the action of these bacteria. In this particular case, the evaluation of C. maxima height showed no significant differences among the treatments.
However, the plants treated with B. thuringiensis CIGB R23 weighed more than the rest of the treatments, with a 17 % increase over the control. Such difference was mostly made by the roots. The roots of B.thuringiensis CIGB R23 were heavier than the control roots (Fig. 2) in spite of having fewer number of galls. (Table 1)
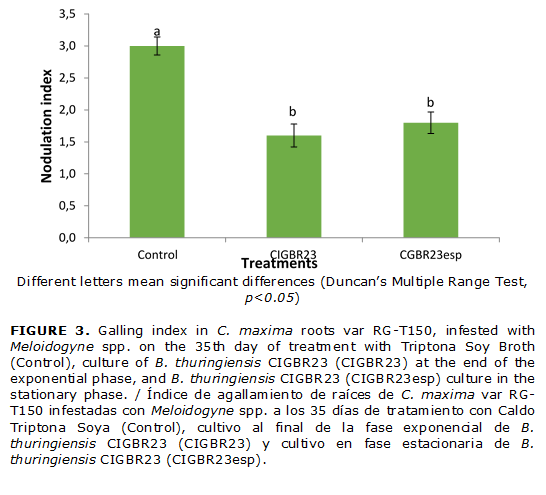
The nematicidal activity of B. thuringiensis CIGB R23 was similar, both for the culture at the end of the exponential phase and in the stationary phase (48 hours), which means that its antagonistic effect was not associated with the Cry proteins released by these bacteria. (Fig. 3)
The extracellular enzymes and the production of H2S are some possible pathogenic traits that may contribute to the nematicidal activity of the bacterial strains. In this case, none of the three strains showed some any of these traits. (Table 2)

Sphingobacterium sp. CIGBRTb did not release extracellular proteases or hydrogen sulfide into the culture medium. Nevertheless, growth without hydrolysis halo formation was observed at 72 hours of incubation in M9 minimal media with chitin or chitosane as the only source of carbon. The previous observation indicated that the strain excreted chitinase and chitosanase enzymes in small quantities. (Fig. 2)
S.maltophilia CIGBG1 produced gelatinases, caseinases, chitinases, phospholipases and hydrogen sulfide. Besides the Cry I toxin present in the parasporal crystals and detected by PCR, B. thuringiensis CIGBR23 also released several hydrolytic enzymes (lipase, caseinase, and chitinase) into the medium.
DISCUSSION
This study confirmed an effective antagonism against Meloidogyne of three bacterial strains isolated from nematode juveniles and eggs, previously chosen for their in vitro activity against Haemonchus sp. (5). The similar chemical composition of eggs suggested a similar mechanism to reduce infestation of that particular nematode in plants (6), so the addition of the bacteria to the soil should reduce the number of juveniles that infect the roots, and consequently, a smaller number of galls are produced. These findings suggest the hypothesis that antagonist bacteria of zoonematode eggs are also effective on plant nematodes.
The three bacterial genera were previously reported as components of vine roots, with nematode suppressing activity. Aballay et al. proved the nematicidal effect of Bacillus brevis 200, Bacillus cereus 146, Bacillus megaterium 185, and Stenotrophomonas maltophilia 168 on the ectoparasitic nematode Xiphinema index Thorne and Alleninpot trials and in vitro assays,whereas S. spiritivorum 64 was effective only in vitro assays (21).
S. maltophilia has shown potential to be used as a biocontrol agent, particularly, on phytopathogenic fungi in soils with high salinity (7) and on the nematode X. index (21). However, there are few reports on its application to control sedentary plant nematodes. In this study, S. maltophilia CIGBG1 was able to control the sedentary nematode Meloidogyne spp.
Sphingobacterium merely had three species until 2000. Though it was found in nematode suppressive soils (21), only S. spiritivorum (strain C926), isolated from rhizosphere, was known to have effective activity against Meloidogyne spp. and Radopholus spp. in field trials (22). Sphingobacterium sp.CIGBTbis another strain from the same genus (candidate to a novel specie), now isolated from Trichostrongylus sp (5), which can be used for nematode biocontrol. Furthermore, it showed the best behavior of the three strains studied. In the last fifteen years, twelve species of this genus have been described, including Sphingobacterium nematocida (8), a bacterium found as a result of the research on nematicidal endophytic microorganisms in China.
Rather than to parasitism, the antagonistic effects of bacteria on plant nematodes are due to the activity of metabolic products such as antibiotics, butyric acid, hydrogen sulfide, proteases, chitinases, peroxidases, compounds related to induced systemic resistance, and the release of volatile compounds with a strong nematicidal activity (9). Therefore, the study of these traits in the above mentioned isolates may indicate the possible mechanism they use to control Meloidogyne spp.infestation. Chitinase enzymes were found in three nematicidal strains, whereas lipases and proteases were observed in two strains. Besides, phospholipases, chitosanases and hydrogen sulfide were detected in at least one of the strains studied.
Sphingobacterium sp. CIGBTb was the best strain, though it has fewer pathogenic traits than the rest. This could indicate the possible presence of a new virulence factor. Only chitosanase, trypsin, esterases, and N-Acetyl-β-D-Glucosaminidase enzymatic activity was observed. These enzymes might hydrolyze some nematode egg surface components (chitin, protein, and short-chain esters) and facilitate the access of some metabolites released by the bacterium with nematicidal activity. Several Sphingobacterium and Stenotrophomonas strains that degrade chitosane could produce chitosanases similar to ChoA of Mitsuaria chitosanitabida (23).
B. thuringiensis CIGBR23 had several possible pathogenicity traits. However, the production of Cry 1 type endotoxin, which is known to act against Lepidoptera (24), was detected by PCR. It meant that the biological activity against Meloidogyne spp. was not caused by delta endotoxins. The previously detected chitinase might play a critical role in the strain´s pathogenicity. Phospholipases and proteases of CIGBR23 might also affect the vitellin and lipid layers on the egg cover of invertebrates (25).
S.maltophilia CIGBG1 had the same enzymes, but it also produced hydrogen sulfide (H2S), which has nematicidal properties (9). It might penetrate the already-weakened outer coverings, due to the action of hydrolytic enzymes, accumulate inside the organism and cause its death. Those pathogenic traits might contribute to the great versatility and adaptability of this bacterium. to different environmental conditions. Consequently, there is an increase in the number of studies for its application as biocontrol and as a plant growth stimulator (7). This genus is able to suppress diseases thanks to its antibiotic secretions, production of extracellular enzymes (proteases and chitinases), and potential colonization of roots (7).
CONCLUSIONS
B. thuringiensis CIGBR23, Sphingobacterium sp.CIGBTb, and S. maltophilia CIGBG1 are native bacterial strains from the juveniles and eggs of plant and animal parasitic nematodes that create new potential alternatives for the biological control of
Meloidogyne spp.. The three strains used at concentrations between 107 and 108 reduced nematode damage to plant roots; hence, they could be used to develop more versatile nematicidal formulations for different kinds of soils. However, in the case of S. maltophilia, it has emerged as an opportunist pathogen in humans, and, at present, it is impossible to distinguish harmful and beneficial strains (7). The genomics and transcriptomic techniques open new possibilities for the knowledge of the taxonomy, ecology and therapeutic treatment of S. maltophilia and otheropportunist pathogens and, therefore, for their application in biotechnology.
1. Sikora RA, Fernández E. Nematode parasites of vegetables. In: Luc M, Sikora RA, Bridge J, editors. Plant parasitic nematodes in subtropical and tropical agriculture. CAB International. Wallingford, UK. 2005; p. 319-92.
2. Gómez L, Rodríguez M, Enrique R, Miranda I, González E. Factores limitantes de los rendimientos y calidad de las cosechas en la producción protegida de hortalizas en Cuba. Rev Protección Veg. 2009;24(2):117-22.
3. Whitehead AG. Plant nematode control: CAB International. 1998;384 pp
4. Berg G. Plant–microbe interactions promoting plant growth and health: perspectives for controlled use of microorganisms in agriculture. Applied microbiology and biotechnology. 2009;84(1):11-8.
5. Sánchez I, Mena J, Coca Y, Marín M, Hernández A, Olazábal A , et al.Acción in vitro de cepas bacterianas sobre Haemonchus spp. Informe Preliminar. Rev Salud Anim. 2003;25(3): 145-8.
6. Bird AF, Bird J. The structure of nematodes. San Diego, USA: Academic Press. 1991;316 p.
7. Berg G, Martinez JL. Friends or foes: can we make a distinction between beneficial and harmful strains of the Stenotrophomonas maltophilia complex? Frontiers in Microbiology. 2015;6:241.
8. Liu J, Yang L-L, Xu C-K, Xi J-Q, Yang F-X, Zhou F, et al. Sphingobacterium nematocida sp. nov., a nematicidal endophytic bacterium isolated from tobacco. International journal of systematic and evolutionary microbiology. 2012;62(8):1809-13. doi: doi:10.1099/ijs.0.033670-0.
9. Tian B, Yang J, Zhang K-Q. Bacteria used in the biological control of plant-parasitic nematodes: populations, mechanisms of action, and future prospects. FEMS microbiology ecology. 2007;61(2):197-213. doi: 10.1111/j.1574-6941.2007.00349.x.
10. Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic acids research. 1997;25(24):4876-82.
11. Kimura M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. Journal of molecular evolution. 1980;16(2):111-20.
12. Saitou N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Molecular biology and evolution. 1987;4(4):406-25.
13. Felsenstein J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution. 1985;39(4):783-91.
14. Shimahara K, Takiguchi Y. Preparation of crustacean chitin. Methods in enzymology. 1988;161:417-23.
15. Cheng CY, Li YK. An Aspergillus chitosanase with potential for large‐scale preparation of chitosan oligosaccharides. Biotechnology and Applied Biochemistry. 2000;32(3):197-203.
16. Plou FJ, Ferrer M, Nuero OM, Calvo MV, Alcalde M, Reyes F, et al. Analysis of Tween 80 as an esterase/lipase substrate for lipolytic activity assay. Biotechnology Techniques. 1998;12(3):183-6.
17. Seeley HW, VanDemark PJ, Lee JJ. Microbes in Action. A Laboratory manual of Microbiology. Fourth Edition ed. WH Freeman and Company. New York; 1991. p. 450.
18. Frazier WC. A method for the detection of changes in gelatin due to bacteria. The Journal of Infectious Diseases. 1926:302-9.
19. Clarke PH. Hydrogen sulphide production by bacteria. Microbiology. 1953;8(3):397-407.
20. Hussey R, Janssen G. Root-knot nematodes: Meloidogyne species. In: Starr J, Cook R, Bridge J, editors. Plant resistance to parasitic nematodes. CAB International. Wallingford, UK. 2002,p. 43-70.
21. Aballay E, Prodan S, Zamorano A, Castaneda-Alvarez C. Nematicidal effect of rhizobacteria on plant-parasitic nematodes associated with vineyards. World J Microbiol Biotechnol. 2017;33(7):131. doi: 10.1007/s11274-017-2303-9.
22. Mena J, Pimentel E, Veloz L, Hernández A, León L, Ramírez Y, et al. Aislamiento y determinación de cepas bacterianas con actividad nematicida. Mecanismo de acción de C. paurometabolum C-924 sobre nematodos. Biotecnología Aplicada. 2003;20(4):248-52.
23. Yun C, Amakata D, Matsuo Y, Matsuda H, Kawamukai M. New Chitosan-degrading strains that produce chitosanases similar to ChoA of Mitsuaria chitosanitabida. Appl Environ Microbiol. 2005;71(9):5138-44. doi: 10.1128/AEM.71.9.5138-5144.2005.
24. Palma L, Muñoz D, Berry C, Murillo J, Caballero P. Bacillus thuringiensis Toxins: An overview of their biocidal activity. Toxins. 2014;6(12):3296-325. doi: 10.3390/toxins6123296.
25. Yang J, Liang L, Li J, Zhang KQ. Nematicidal enzymes from microorganisms and their applications. Appl Microbiol Biotechnol. 2013;97(16):7081-95. doi: 10.1007/s00253-013-5045-0.
Recibido: 25/10/2017
Aceptado: 27/3/2018
*Autor para correspondencia: Ileana Sánchez Ortiz. E-mail: ileana.sanchez@cigb.edu.cu

