










Meu SciELO
 Serviços customizados
Serviços customizadosServiços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista de Protección Vegetal
versão impressa ISSN 1010-2752versão On-line ISSN 2224-4697
Rev. Protección Veg. vol.33 no.1 La Habana jan.-abr. 2018
ARTÍCULO RESEÑA
Moniliophthora roreri H.C. Evans et al. y Moniliophthora perniciosa (Stahel) Aime: impacto, síntomas, diagnóstico, epidemiología y manejo
Moniliophthora roreri H.C. Evans et al. and Moniliophthora perniciosa (Stahel) Aime: impact, symptoms, diagnosis, epidemiology and management
Luis Pérez-Vicente*
Instituto de Investigaciones de Sanidad Vegetal (INISAV), Gaveta 634, 11300, Playa, La Habana Cuba.
RESUMEN
La escoba de brujas y la moniliasis del cacao, causadas por Moniliophthora perniciosa (Stahel) Aime ypor Moniliophthora roreri H.C. Evans et al. respectivamente, son dos enfermedades originariasde América del Sur, de distribución geográfica limitada y con un fuerte impacto en la producción y la calidad del cacao. Los informes de la presencia de ambos patógenos en Cuba son inexactos y no se encontraron en las encuestas realizadas por el autor, entre2013 y 2016. Los informes previos, relativos a la presencia de escoba de brujas en Cuba, se corresponden con la enfermedad denominada agalla de punto verde| causada por Fusarium decemcellulare Brick. En el presente trabajo se documentan los estudios realizados por el autor sobre los síntomas, estructuras morfológicas y colonias de ambos hongos en Ecuador, Venezuela y Costa Rica y se actualizan las informaciones sobre sus relaciones filogenéticas, su epidemiología y manejo. Las esporas de ambas especies se dispersan por salpicaduras de agua, el viento y por partes de plantas infectadas. Los cosecheros son importantes vectores de las esporas de M. roreri a través de la ropa contaminada. El manejo de ambas enfermedades se basa en medidas estrictas de cuarentena, saneamiento y destrucción de partes de plantas infectados, el uso de clones resistentes y aplicaciones de fungicidas y antagonistas a troncos, ramas y frutos. Ambas especies de Moniliophthora se encuentran en la lista de plagas cuarentenarias de Cuba. La presencia de M. roreri en Jamaica constituye una amenaza para la producción cacaotera del oriente de Cuba.
Palabras clave: cacao, control, cuarentena, escoba de brujas, moniliasis
ABSTRACT
Witches’ broom and frosty pod rot of cacao caused by Moniliophthora perniciosa (Stahel) Aime and Moniliophthora roreri H.C. Evans et al. respectively, are two diseases original from South America with a limited geographic distribution and a strong impact on production and quality of cacao. The reports of their presence in Cuba are not exact and the pathogens were not found in the surveys conducted by the author between 2013 and 2016. The previous reports on the witches’ broom presence in Cuba were related to the disease known as green point gall caused by Fusarium decemcellulare Brick. In the present paper, the studies carried out by the author on the symptoms, morphological structures, and colonies of both pathogens in Ecuador, Venezuela, and Costa Rica are documented, and the information on their phylogenetic relationships, epidemiology and management are updated. The spores of both species are disseminated by water, wind, and infected plant parts. Cacao harvesters are important vectors of M. roreri spores being transported on clothes. Prevention and management of both diseases are based on strict quarantine measures; sanitation and destruction of branches, stems, and infected fruits; use of resistant clones and applications of fungicides and antagonists to stems, branches, and fruits. Both species are included in the list of quarantined pests of Cuba. The presence of M. roreri in Jamaica is a threat for the cacao production in the west of Cuba.
Key words: cacao, control; quarantine, witches’ broom, frosty pod rot.
INTRODUCCIÓN
Theobroma y otros dos géneros, Herrania y Guazuma, pertenecen a la familia Sterculiaceae. El cacao Theobroma cacao L. es uno de los más importantes cultivos tropicales y posee dos centros de diversidad reportados: (1) uno en la Amazonia de América desde el Sur de Colombia y Venezuela hasta el este del Perú y Bolivia en el Sur, y (II): otra área de diversificación en México, América Central y las islas del Caribe. Las semillas de cacao son la fuente del chocolate, la manteca de cacao y polvo; actualmente la industria chocolatera mundial es valorada en 80 mil millones USD/año (2). Antes del siglo XX, los principales países exportadores de cacao estaban en América del Sur y Central, región que ahora solo produce un poco más del 10 % debido, en gran medida, a las enfermedades (3). La producción del oeste de África representa el 70 % del volumen mundial.
El mayor factor limitante a nivel mundial del cultivo de cacao es una trilogía de importantes enfermedades (4): la pudrición negra de la mazorca, causada por Phytophthora palmivora (E. J. Butler) E. J. Butler, Phytophthora megakarya Brasier & M.J. Griffin, Phytophthora capsici Leonian y Phytophthora citrophthora (R.H. Sm. & E. Sm.) Leonian (5), enfermedad que más afecta la producción globalmente; la escoba de brujas, provocada por Moniliophthora perniciosa (Stahel) Aime y la moniliasis, cuyo agente etiológico es Moniliophthora roreri H.C. Evans et al., las cuales tienen el potencial de devastar el suministro mundial de cacao si se diseminan fuera de América.
En el año 2014, la superficie cultivada de cacao en Cuba fue, aproximadamente, de 4155 ha, la producción de 2188 ton, con rendimientos entre 200 y 450 kg/ha (6). En Cuba se informó la presencia de la escoba de brujas y de la moniliasis (7,8).
El presente trabajo tiene como objetivos actualizar los conocimientos acerca de la distribución geográfica, el impacto económico, la epidemiología, el diagnóstico y el manejo de M. perniciosa y M. roreri, así como informar los resultados de las observaciones sobre los síntomas que causan y la morfología de ambos patógenos, junto a los obtenidos en una encuesta de enfermedades realizada en Cuba para confirmar la presencia de ambas enfermedades en plantaciones de cacao.
Relaciones filogenéticas y taxonomía de M. perniciosa y M. roreri
Evans (9) fue el primero en aportar evidencias de la naturaleza hemibiotrófica de los entonces llamados Crinipellis perniciosa y Monilia roreri. Sobre la base de observaciones in vivo e in vitro, mostró evidencias de que el agente causal de la moniliasis del cacao era un basidiomiceto (10,11) y sugirió el nuevo nombre genérico Moniliophthora. El nuevo binomio utilizado para nombrar este patógeno es Moniliophthora roreri (Cif.) H.C. Evans, Stalpes, Sampson, & Benny (11, 12, 13). A partir del análisis filogenético de las regiones 28S rDNA,18S rDNA, ITS, RPB1 y EF1-a, Aime y Phillips-Mora (14) confirmaron que Crinipellis perniciosa y Monilia roreri son taxas hermanas que pertenecen a los Marasmiaceae (Euagaricales) con un basidioma agaricoide y que forman un linaje separado dentro de la familia, que incluyen los hongos hemibiotróficos Moniliophthora perniciosa comb. nov. y M. roreri. Los géneros hermanos de Moniliophthora son Marasmius, Crinipellis y Chaetocalathus, que son saprófitos de las hojas en descomposición.
La escoba de brujas por M. perniciosa
Distribución. El patógeno y la enfermedad se encuentran presentes en Bolivia, Brasil, Colombia, Ecuador, Granada, Guyana, Guyana Francesa, Panamá (al este del Canal), Perú, Trinidad y Tobago, Sta. Lucia, San Vicente y las Granadinas, Surinam y Venezuela. (Figura 1)

Los informes de su presencia en Cuba (7, 8) son inexactos; Pérez-Vicente et al. (15) realizaron una encuesta de enfermedades en
2010 en diferentes regiones de la Sierra Maestra, desde Manzanillo, Buey Arriba, Santiago de Cuba, Baracoa hasta Cienfuegos. Muestras de tejidos con síntomas tipo agallas y pequeñas escobas de yemas vegetativas y florales, que se encontraron en la zona de Mosquitero en Baracoa y en Cienfuegos, se sembraron en agar agua y los crecimientos se pasaron a PDA. Los síntomas de agallas y escobas se colocaron por 10 días en cámara húmeda para inducir la formación de estructuras fúngicas y, eventualmente, carpóforos. Sobre la base de este estudio, se concluyó que la enfermedad no está presente en Cuba y que los síntomas de agallas y pequeñas escobas asociados a agallas vegetativas y florales, encontrados en plantas de cacao en ambas zonas, se producen por Fusarium decemcellulare Brick (1908) [(Albonectria rigidiuscula (Berk. & Broome) Rossman & Samuels (1999)], agente causal de la agalla de punto verde o buba del cacao. No se documentaron la presencia de basidiocarpos de M. perniciosa en plantaciones ni a partir de las agallas colocadas en condiciones alternantes de humedad y secado; tampoco se obtuvieron colonias del patógeno a partir de tejidos enfermos. Observaciones similares fueron documentadas en Haití por Purdy (16).
Síntomas e impacto. Se describen los síntomas observados en plantas de cacao afectadas, presentes en plantaciones de cacao cercanas a la ciudad Pucalpa en la Amazonia del Perú, durante 1996 -1998; de la Estación del INIA, en plantaciones cercanas al Vigía al Sur del Lago Maracaibo (Venezuela) en 2010 y de Guayaquil; así como en la Estación Experimental de Pichilingue en Quevedo en 2012.
Los síntomas de la enfermedad son conspicuos (Figura 2) y su desarrollo sigue a la infección de los meristemos activos vegetativos y florales por las meiosporas (basidiosporas) durante el tiempo húmedo y cálido y a la instalación de hifas haploides uninucleadas gruesas en los tejidos (17, 18); se manifiestan por la aparición de brotes vegetativos engrosados por la hiperplasia e hipertrofia de los tejidos en desarrollo, a la proliferación de brotes vegetativos y florales que se desarrollan como escobas que alcanzan hasta 60 cm de largo (Figura 2). Los brotes florales afectados producen cojinetes vegetativos y rosetas de frutos tipo fresas sin semillas o de forma de zanahoria. M. perniciosa puede infectar las mazorcas y necrosar los granos. Los árboles afectados reducen, drásticamente, la producción de mazorcas. En los tejidos afectados se producen los carpóforos típicos de los agaricales.

La producción de cacao recibió impactos de diferente magnitud en los países donde la enfermedad está presente. La enfermedad destruyó las plantaciones de Guyana y Surinam a inicio del siglo XX, donde causó el abandono de la producción de cacao posteriormente a su introducción (11,13). Se diseminó a Ecuador en 1920 y al Caribe en 1928. En áreas costeras del Ecuador la producción se redujo en el 50 %; mientras que, en Bahía, Brasil, determinó que se convirtiera en importador neto de granos de cacao (13, 20, 22). Las pérdidas estimadas a nivel mundial por la escoba de brujas en el año 2001 fueron de 250 mil t y de 380,000 t en 2012 (19,21). Donde la enfermedad aparece, se reduce dramáticamente la producción (20,21).
Ciclo de la enfermedad. El ciclo de la enfermedad (Figura 3) se inicia con la germinación de las basidiosporas que producen hifas monocarióticas gruesas (5-20 μm), las que infectan los tejidos meristemáticos de los brotes vegetativos y florales a través de estomas o heridas, colonizan el apoplasto y causan transformaciones drásticas en los tejidos del hospedante debido a la producción de hormonas que inducen la formación de las escobas (23). Los basidiocarpos de M. perniciosa nunca se forman durante la fase monocariótica biotrófica del hongo, en escobas verdes. Después de 6-9 semanas, cuando las escobas en tallos y mazorcas infectados mueren y se secan y bajo condiciones alternantes de humedad y secado, el patógeno cambia a su fase necrotrófica y se forman los basidiocarpos a partir de un micelio de hifas delgadas dicarióticas con fíbulas ( 24,25), los cuales liberan las basidiosporas al aire durante 2-8 días y se reinicia el ciclo de infección. En esta fase, conocida como escoba seca, M. perniciosa coloniza la planta muerta y puede ser encontrado en los espacios intercelulares del cacao.

La formación de las escobas se debe, mayormente, al agrandamiento de las células que a la división celular. Bajo condiciones de humedad alternantes (8 horas de humedad/16 horas secado), las temperaturas entre 20 y 25°C son favorables para la formación de basidiocarpos, mientras que por encima de 30°C son menos favorables (26). Se tomaron muestras e imágenes de basidiocarpos del hongo encontrados en campo e inducidos en escoberos en la Estación de Pichilingue. (Figura 2 D)
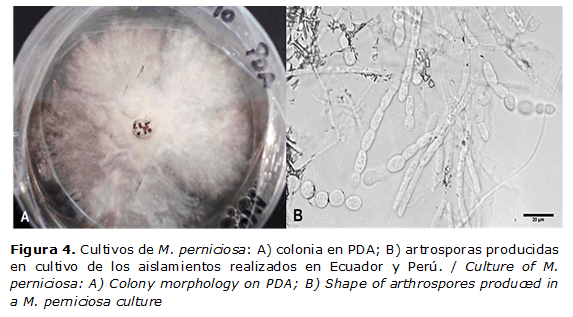
Los basidiocarpos pueden producirse en laboratorio después de ocho semanas de incubación sobre fragmentos de escobas de cacao autoclavadas, colocadas sobre agar agua o agar jugo V-8, e inoculados con micelio (24). Las basidiosporas se pueden dispersar por el viento a largas distancias entre 60 y 150 km (21), aunque el movimiento antropogénico en mazorcas, semillas y esquejes con infecciones latentes ha sido el responsable de los grandes saltos en América. La penetración del hospedante puede ocurrir en un tiempo mínimo de seis horas, pero el porcentaje máximo ocurre entre 12 -15 horas a 25ºC en 100 % HR (27). Se examinaron basidios, basidiosporas y micelio para determinar la condición nuclear en diferentes estados del crecimiento de M. perniciosa. Los basidios jóvenes fueron uninucleados, de los cuales se desarrollan cuatro basidiosporas mononucleadas después de la meiosis. Las hifas primarias derivadas de las basidiosporas son septadas, con células uninucleadas donde persiste un corto tiempo para dar lugar a un micelio denso. La aparición posterior de un micelio denso, septado binucleado con fíbulas, sugiere que el hongo es homotálico. Las artrosporas que se formaron a partir de aislamientos en cultivo en medio de agar malta y PDA obtenidos de escobas de Ecuador y Perú son binucleadas, pero no son infectivas (28). En la Figura 4 aparecen imágenes de las colonias y las artrosporas obtenidas de los aislamientos realizados en Ecuador y Perú.
Variabilidad genética de M. perniciosa
El rango de hospedantes de M. perniciosa incluye a Theobroma spp., Herrania spp., Solanum spp. y Bixa orellana L. Se documentó, por diferentes autores, la existencia de una considerable variabilidad patogénica entre los aislamientos de M. perniciosa (13, 20, 25, 29, 30).
Diagnóstico. M. perniciosa puede ser rápidamente aislado de escobas en diversos medios de cultivo, donde da lugar a colonias típicas y producción de artrosporas (Figura 4). Se ha desarrollado el diagnóstico por PCR mediante cebadores del gen actin de M. perniciosa a partir de ADN aislado de hifas del patógeno (31, 32).
Manejo de la escoba de brujas. Existe alguna resistencia genética, aunque no necesariamente durable. La poda sanitaria y la destrucción sistemática de residuos a nivel regional, aunque laboriosa y costosa, es rentable. Se requiere regular la altura de las plantas para hacer posible el saneamiento; este, combinado con la aplicación sistemática de Trichoderma spp. han resultado efectivos, aunque demanda trabajo intenso (33, 34, 35).
La moniliasis por M.roreri
Distribución. La enfermedad fue reconocida tan tempranamente como en 1817 en el noreste de Colombia (37). El patógeno se encuentra presente en Belice, Bolivia, Colombia, Costa Rica, Ecuador, Nicaragua, Panamá, Perú y el noreste de Venezuela (Figura 5).


El primer reporte en las Antillas se hizo en Jamaica en septiembre de 2016 (38). El centro de origen del patógeno parece estar ubicado en el noreste de Colombia, donde fue encontrada la mayor diversidad de genotipos (39). Es sorprendente el hecho de que no tenga una distribución mayor dado a la gran capacidad de producción de esporas, la resiliencia de las mismas a las condiciones ambientales, la facilidad con que se dispersan, el largo periodo
de latencia en mazorcas infectadas que facilitan el movimiento a grandes distancias antes de que aparezcan síntomas y la gran susceptibilidad del cacao y otras especies.
Síntomas e impacto. M. roreri afecta los géneros afines de Theobroma y Herrania. La moniliasis es la enfermedad más seria del cultivo, debido a las pérdidas potenciales que es capaz de producir y la dificultad para su manejo (9,24). Las medidas intensivas requeridas para el manejo de la enfermedad están más allá de las posibilidades de muchos productores, porque dependen de un saneamiento eficiente, actividad que encarece el costo de producción (24,40). Las pérdidas informadas en 2001 fueron de 30 % de la producción mundial, con un costo de 51 millones de USD (19); mientras que en 2013 estas ascendieron a 50 mil t (21). En Ecuador, después de la aparición de la moniliasis, la producción declinó de 50,000 a 30,000 t; en Costa Rica, de 12 mil t en 1962 a 708 en 2001 por diferentes causas, entre ellas, la influencia de la enfermedad (21). La aparición de la moniliasis fue la causa de, al menos, el abandono del 50 % de las plantaciones de cacao en diversas áreas afectadas (35).
Se tomaron muestras y fotos de síntomas de la enfermedad en plantaciones de cacao de Guayaquil y Los Ríos en el Ecuador y del jardín clonal de T. cacao y Herrania sp., del programa de mejoramiento del CATIE en Turrialba, Costa Rica. El patógeno infecta los frutos en desarrollo y causa una secuencia de síntomas (Figura 7): la aparición de decoloraciones acuosas y grasientas (hidrosis), la deformación de los tejidos de los frutos (tumefacción), el desarrollo de manchas irregulares pardas (necrosis) en los tejidos afectados, la maduración precoz de las mazorcas, la aparición durante tiempo favorable de grandes masas blancas de esporas, la momificación de los frutos y las pudriciones acuosas interiores del fruto y de las semillas del cacao.
Ciclo de la enfermedad. El hongo fue primeramente descrito como Monilia roreri (Ciferri y Parodi), un género discomiceto anamórfico en los Ascomycota. La clasificación se basó en la producción de conidios en cadenas largas. Más tarde, se documentó que realmente son probasidios dicaríoticos que han sufrido la primera división meiótica, aunque no la segunda (9,11), y que posee los locus bialelicos de apareamiento A y B (41); por lo tanto, se considera que M. roreri lleva a cabo una reproducción meiótica sin formar basidiocarpos. Ambos tipos de apareamiento se encontraron en América del Sur, pero solo uno en América Central (41), lo que indica una dispersión clonal en esta última. Estas meiosporas se producen en masas polvorientas blancas sobre un pseudostroma carnoso en la superficie del fruto afectado. (Figura 7)

Las esporas son rápidamente dispersadas por el viento y la lluvia o acarreadas en la ropa de trabajadores y animales, estas infectan mazorcas de cacao no maduras y causan la hinchazón y la maduración prematura del área infectada, seguido de la producción de inóculo abundante (hasta 44 millones de esporas/cm2) para nuevas infecciones (24). (Figura 8)

Los frutos más susceptibles a la enfermedad son los de menos de tres meses de edad. Un factor importante de distribución de la enfermedad ha sido el movimiento de mazorcas infectadas entre países.
Diagnóstico. Los síntomas de las plantas y la observación de las meiosporas constituyen un diagnóstico seguro de la enfermedad. De mazorcas afectadas de cacao del CATIE en Turrialba, se transfirieron esporas a PDA, V-8 y agar malta. En estos medios las esporas del patógeno germinan y dan lugar a colonias típicas que permiten la identificación. (Figura 7)
Prevención y manejo de la moniliasis
- Prevención: M. roreri solo infecta Theobroma y Herrania spp. i) Los países libres deben prohibir transportar mazorcas de áreas infestadas, sobre todo de campos de producción. El patógeno tiene un largo periodo de latencia que puede burlar la vigilancia; ii) preferiblemente debe usarse una cuarentena intermedia y tratar mazorcas con fungicidas; iii) entrenar inspectores de frontera e investigadores en detección.
- Manejo: i) saneamiento y destrucción semanal de mazorcas infectadas antes de que comiencen a esporular (fase hidrosis Fig.6A; tumefacción y manchas pardas). El corte debe ser hecho al pedúnculo del fruto; ii) realizar podas para mantener follaje con porte bajo y poder desarrollar saneamientos; iii) recoger, en los lugares donde hay picos de cosecha, todas las mazorcas sanas y enfermas al final de la cosecha para interrumpir ciclo; iv) aplicaciones de fungicidas (cobre de forma frecuente; sin embargo, los costos son un problema); v) uso de antagonistas en conjunto con las podas; vi) variedades resistentes: la mayor promesa para el manejo de la enfermedad es la obtención de líneas híbridas de cacao tolerantes. El CATIE desarrolló varias líneas tolerantes, entre ellas el CATIE-R4 y CATIE-R6 que poseen, además, tolerancia a Phytophthora (22).
El éxito del saneamiento depende de la sistematicidad en la eliminación de mazorcas infectadas antes de la aparición de la esporulación y su subsecuente destrucción. Para los epidemiólogos experimentados en el manejo de la moniliasis parece ser fácil, debido a que se transmite solo por esporas producidas en las mazorcas infectadas; sin embargo, en la práctica es sumamente difícil, bordeando a la calamidad. Entre los factores que impiden un adecuado control se encuentran: a) la capacidad de producción y las vías de dispersión de las esporas; b) la existencia de plantaciones que reciben poco cuidado cultural; c) la presencia de árboles altos que no permiten saneamiento de mazorcas infectadas; d) la pobre eficiencia del uso de los equipos de aspersión; e) la negativa de gobiernos de establecer un control químico sistemático en las plantaciones.
CONSIDERACIONES FINALES
- La escoba de brujas y la moniliasis son enfermedades de alto impacto en la producción de cacao, especialmente en áreas con pluviometría entre 1200 y 4000 mm anuales de lluvia. Estas condiciones son comunes en la principal área productora de cacao en Cuba.
- La presencia de M. roreri en Jamaica y el consecuente paso del huracán Matthew por la región en septiembre de 2016, pudo distribuir esporas del patógeno sobre la zona oriental, particularmente Baracoa, la principal zona de producción. Es imperativo desarrollar la vigilancia fitosanitaria y la capacitación de productores para un reconocimiento temprano de brotes potenciales.
- Dado el impacto de estas enfermedades en la producción y los costos, la prevención y las medidas de cuarentena con relación al manejo de germoplasma es esencial.
- La adquisición de germoplasma resistente a ambas enfermedades, siguiendo estrictos protocolos de bioseguridad, resulta estratégico para la sostenibilidad de la producción de cacao en Cuba.
1. Bartley BGD. The genetic diversity of cacao and its utilization. Wallingford: CABI Publishing.2005.341pp
2. The Guardian. The future of chocolate: Why cocoa production is at risk. Accessed: February 26, 2015, from http://www.theguardian.com/sustainable-business/fairtrade-partnerzone/chocolate-cocoa-production-risk.
3. Gray A. The World Cocoa Market Outlook. London: LMC International, Ltd. 2001.29p.
4. Fulton RH. The Cacao Disease Trilogy: Black Pod, Monilia Pod Rot, and Witches'-Broom. Plant Disease.1998; 73:601-603
5. Appiah AA, Flood J, Archer SA, Bridge PD. Molecular analysis of the major Phytophthora species on cocoa. Plant Pathology. 2004; 53:209-219.
6. FAOStat (2017). http://www.fao.org/faostat/en/#data/QC
7. Arnold GRW. Lista de Hongos Fitopatógenos de Cuba. Ministerio de Cultura. Editorial Científico-Técnica, 1986. 207 pp.
8. Farr DF, Rossman AY. Fungal Databases, U.S. National Fungus Collections, ARS, USDA. Retrieved March 15, 2017, from https://nt.ars-grin.gov/fungaldatabases
9. Evans HC. Pod rot of cacao by Moniliophthora roreri. Phytopathological Paper.1981; 24:1-44.
10. Evans HC, Stalpers JA, Samson RA, Benny GL. On the taxonomy of Monilia roreri, an important pathogen of Theobroma cacao in South America. Canad. J. Bot. 1978; 56:2528-2532
11. Evans HC, Holmes K, Phillips-Mora W, Wilkinson JM. What's in a name: Crinipellis, the final resting place for the frosty pod rot pathogen of cocoa?Mycologist. 2002; 16:148-152.
12. Evans HC, Holmes KA, Reid AP. Phylogeny of the frosty pod rot pathogen of cocoa. Plant Pathology. 2003; 52:476-485.
13. Evans HC, Bezerra JL, Barreto RW. Of mushrooms of chocolate trees: Aetiology and phenology of witches broom and frosty pod diseases of cacao. Plant Pathology. 2013; 62:728-74
14. Aime MC, Phillips-Mora W. The causal agent of witches’ broom and frosty pod rot of cacao (chocolate, Theobroma cacao) form a new lineage of Marasmiaceae. Mycologia. 2005; 97:1012–1022.
15. Pérez-Vicente L, Martínez-de La Parte E, Cantillo T. First report of cushion gall or green point ball of cocoa by Albonectria rigidiuscula (Fusarium decemcellulare) in Cuba. Fitosanidad. 2012; 16(1):19-25
16. Purdy LH. Assessment of the Presence of Witches’ Broom of Cacao. In: Matlick, B. K., Purdy, L. H. and Stevenson, C. (Ed.). Technical Support to Haitian Cacao. Productive land use system (PLUS) Haití. South East Consortium for International Development (SECID) and Auburn University. SECID/Auburn PLUS Report No. 48. USAID/ Haiti Economic Growth Office.1999. 31pp.
17. Pegus JE. Aspects of the host–parasite relationship in the Theobroma cacao L./Marasmius perniciosus Stahel disease complex. M.Sc. thesis. 1972. University of West Indies, St. Augustine, Trinidad.18 pp
18. Purdy LH, Schmidt, RA. Status of cacao witches' broom: biology, epidemiology and management. Annu. Rev. Phytopathol. 1996: 34:573–594.
19. Bowers JH, Bailey BA, Hebbar PK, Sanogo S, Lumsden RD. The impact of plant diseases on world chocolate production. Plant Health Progress. 2001. 17pp. doi:10.1094/PHP-2001-0709-01-RV. Published online.
20. Griffith GW, Nicholson J, Nenninger A, Birch RN, Hedger JN. Witches’ brooms and frosty pods: Two major pathogens of cacao. New Zealand Journal of Botany. 2003; 41:423–435.
21. Ploetz RC. The Impact of Diseases on Cacao Production: A Global Overview. (Chapter 2) Cacao Diseases. © Springer International Publishing Switzerland. 2016: 33-59. DOI 10.1007/978-3-319-24789-2_2
22. Evans HC. Cacao diseases-the trilogy revisited. Phytopathology2007. 97:1640-1643.
23. Teixeira PJPL, Thomazella DPT, Vidal RO, Prado PFV do, Reis O, et al. The fungal pathogen Moniliophthora perniciosa has genes similar to plant PR-1 that are highly expressed during its interaction with cacao. PLoSONE.2012;7(9): e45929. doi:10.1371/journal.pone.0045929.
24. Purdy LH, Dickstein ER. Basidiocarp development on micelial mats of Crinipellis perniciosa. Plant Disease. 1990; 74:493-496.
25. Meinhardt LW, Rincones J, Bailey BA, Aime MC, Griffith GW, et al. Moniliophthora perniciosa, the causal agent of witches’ broom disease of cacao: what’s new from this old foe? Mol. Plant Pathol. 2008; 9:577–588.
26. Rocha HM, Wheeler, BEJ. Factors influencing the production of basidiocarps and the deposition and germination of basidiospores of Crinipellis perniciosa the causal agent of whitches’ broom disease of cocoa. Plant Pathology.1985; 34: 319-328.
27. Frias GA, Purdy LH, Schmidt RA. An inoculation method for evaluating resistance of cacao to Crinipellis perniciosa. Plant Disease. 1995; 79:787–91.
28. Delgado JC, Cook AA. Nuclear condition of the basidia, basidiospores and mycelia of Marasmius perniciosus. Can. J. Bot. 1976. 54:66–72.
29. Wheeler BJE, Mepsted R. Pathogenic variability among isolates of Crinipellis perniciosa from cocoa. Plant Pathology.1988; 37:475-478.
30. Osorio-Solano C, Orozco-Castaño CA, López-Gartner G, Rivera-Páez FA. Genetic variability of Moniliophthora perniciosa (Stahel) Aime & Phillips-Mora, comb. nov. (Agaricales – Marasmiaceae) in varieties of cocoa (Theobroma cacao L.). Acta Agronómica. 2012; 61(2):85-93.
31. Melo SO, Pungartnik C, Cascardo JCM, Brendel M. Rapid and efficient protocol for DNA extraction and molecular identification of the basidiomycete Crinipellis perniciosa. Genet. Mol. Res. 2006; 5(4):851-855.
32. Tarnoski TLB. Using molecular analysis to investigate phylogenetic relationships in two tropical pathosystems: witches’ broom of cacao, caused by Moniliophthora perniciosa, and mango anthracnose, caused by Colletotrichum spp. [Ph.D. dissertation]. 2009. Univ. Florida, Gainesville, USA. 236 pp.
33. Sanogo S, Pomella A, Hebbar PK, Bailey B, Costa JCB, Samuels GJ, Lumsden RD. Production and germination of conidia of Trichoderma stromaticum, a mycoparasite of Crinipellis perniciosa on cacao. Phytopathology. 2002; 92:1032-1037.
34. De Arruda MCC, Sepulveda GF, Miller RNG, Ferreira MASV, Santiago DVR, Resende MLV, Dianese JC, Felipe MSS. Crinipellis brasiliensis, a new species based on morphological and molecular data. Mycologia. 2005; 97(6):1348–1361.
35. Drenth A, Guest D. Fungal and Oomycete diseases of tropical tree fruit crops. AnnualReview of Phytopathology. 2016; 54:373–395.
36. RiveraFernández RDO, ValerezoBeltron L, VerasMacías J, ChavarríaPárraga E, GuzmánCedeño AM. Efecto de la poda fitosanitaria sobre la enfermedad escoba de brujas en el cultivo de cacao. Revista Intropica. 2014; 9:129136.
37. Phillips-Mora W, Wilkinson MJ. Frosty pod, a disease of limited geographic distribution but unlimited potential for damage. Phytopathology. 2007; 97:1644–1647.
38. CIPP. Detection of Frosty Pod Rot in Jamaica. 29 Sep 2016. https://www.ippc.int/es/countries/jamaica/pestreports/2016/09/detection-of-frosty-pod-rot-in-jamaica/
39. Phillips-Mora W, Ortiz CF, Aime MC. Fifty years of frosty pod rot in Central America: Chronology of its spread and impact from Panama to Mexico. In: Proceedings of the 15th International Cocoa Research Conference, San José, Costa Rica. 2007. Vol. I, pp. 1039–1047.
40. Krauss U, Hidalgo E, Bateman R, Adonijah V, Arroyo C, Garcıa J, et al. Improving the formulation and timing of application of endophytic biocontrol and chemical agents against frosty pod rot (Moniliophthora roreri) in cocoa (Theobroma cacao). Biological Control. 2010; 54:230–240.
41. Díaz-Valderrama JR, Aime MC. The cacao pathogen Moniliophthora roreri (Marasmiaceae) produces rhexolytic thallic conidia and their size is influenced by nuclear condition. Mycoscience. 2016; 57:208–16.
42. Phillips-Mora W, Castillo J, Krauss U, Rodríguez E, Wilkinson MJ. Evaluation of cacao (Theobroma cacao) clones against seven Colombian isolates of Moniliophthora roreri from four pathogen genetic groups. Plant Pathology. 2005; 54:483–490
43. Phillips-Mora W, Arciniegas-Leal A, Mata-Quirós A. Catálogo de clones de cacao seleccionados por el CATIE para siembras comerciales. Centro Agronómico Tropical de Investigación y Enseñanza, CATIE. 2012. 66 pp.
44. Campuzano H. Fluctuación de poblaciones de esporas de Monilia roreri Cif. et Par., y viabilidad durante un ciclo completo de afección. Noticias Fitopatológicas. 1976; 5:107.
Recibido: 4/10/2017
Aceptado: 9/10/2017
*Autor para correspondencia: Luis Pérez-Vicente. E-mail: lperezvicente@inisav.cu

