










My SciELO
 Custom services
Custom servicesServices on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkVaccimonitor
Print version ISSN 1025-028XOn-line version ISSN 1025-0298
Vaccimonitor vol.17 no.1 Ciudad de la Habana Jan.-Apr. 2008
Biomodelo para la evaluación de cepas atenuadas como candidatos vacunales contra el cólera humano. I. Estudio de la virulencia, capacidad de colonización y adherencia a la mucosa intestinal
Biomodel for evaluating attenuated Vibrio cholerae strains as human cholera vaccine candidates. I. Virulence, colonizing capacity and adherence to the intestinal mucosa.
Reynaldo Oliva, Hilda García, Juan F. Infante, Luis García, José L. Pérez, Bárbara Cedré, Tania Balmaceda, Arturo Talavera, Gemma Año.
1 Instituto Finlay. Centro de Investigación-Producción de Vacunas. Ave 27 No. 19805, e/ 198 y 202.
La Lisa, Ciudad de La Habana. Cuba.
Correo electrónico: roh@finlay.edu.cu
RESUMEN
El cólera continúa siendo en muchos países un problema para la salud humana, manteniéndose como una enfermedad epidémica o endémica que afecta tanto a niños como adultos y causa la muerte en casos no tratados. Una vacuna viva oral contra esta enfermedad puede ser la solución. En el presente trabajo se seleccionó y aplicó un biomodelo para la evaluación de cepas atenuadas genéticamente de Vibrio cholerae como candidatas vacunales contra el cólera. La virulencia, capacidad de colonización y adherencia a la mucosa intestinal de las cepas fueron evaluadas mediante el uso de ratones neonatos de 2 a 4 días de nacidos de la línea Balb/c, con un peso entre 1,5-2 g. Los resultados obtenidos con este biomodelo demostraron que las cepas atenuadas genéticamente son no virulentas, colonizan y se adhieren a la mucosa intestinal. Se concluye que el biomodelo utilizado permite la evaluación y selección de cepas candidatas para vacunas vivas orales contra el cólera.
Palabras clave: Biomodelo; Vibrio cholerae, cepas atenuadas, ratón neonato.
ABSTRACT
Cholera is still a human health problem in many countries. It is an epidemic or endemic disease affecting both children and adults that causes death of untreated cases. A live oral vaccine could be the solution against this disease. In the present study a biomodel was selected and applied for the evaluation of genetically attenuated Vibrio cholerae strains as vaccine candidates. The virulence, colonizing capacity and adherence to the intestinal mucosa of the strains were evaluated using 2-4 day-old neonatal Balb/c mice, weighing from 1.5-2 g. The results obtained with this biomodel showed that genetically attenuated strains are not virulent, colonize and adhere to the intestinal mucosa. The conclusion was that the biomodel used allows the evaluation and selection of candidate strains for live oral cholera vaccines.
Keywords: biomodel, Vibrio cholerae, attenuated strains, neonatal mice.
Introducción
El cólera humano es una enfermedad diarreica aguda caracterizada por diarreas profusas, con apariencia de agua de arroz. La forma clínica más severa (cólera gravis) se caracteriza por una rápida pérdida de líquido y electrolitos a través del tracto gastrointestinal que de no tratarse adecuadamente conduce al shock hipovolémico, acidosis metabólica e incluso a la muerte en poco tiempo (1).
El agente causal del cólera es Vibrio cholerae de los serogrupos O1 y O139 (2). La toxina producida por este microorganismo (siglas en inglés CT) es la responsable de los principales síntomas clínicos de la enfermedad. CT es una proteína compuesta por una subunidad A (siglas en inglés CTA) (28 kDa) y cinco subunidades B (siglas en inglés CTB) (11,5 kDa cada una). CTA se une de forma covalente a la subunidad reguladora de la enzima adenilato ciclasa, la cual se activa, generándose un aumento intracelular de los niveles de AMP cíclico (AMPc), cuyo efecto final es la salida indiscriminada de agua y electrolitos de la célula. CTB se une de forma específica al gangliósido GM1 presente en la superficie de las células epiteliales del intestino (3).
Además de CT, ambos serogrupos de V. cholerae, O1 y O139, producen otros factores potencialmente tóxicos como la Accesory Cholera Enterotoxin (siglas en inglés ACE), Zónula Occludens Toxin (siglas en inglés ZOT), hemolisina y una hemaglutinina/proteasa (3). El papel de estos factores en la patogenia de la enfermedad es controversial y se encuentra bajo intensa investigación científica.
V. cholerae O1, biotipo El Tor, es el agente causal de la séptima pandemia, iniciada en 1961 y ha continuado su diseminación hacia todas las regiones del mundo (4, 5). La influencia del fenómeno "El Niño" en el clima y otros fenómenos meteorológicos, la insuficiencia en los servicios de salud y el desgaste ocasionado en la infraestructura económico-social por diversos desastres naturales y sociales, son factores que han influido de forma continua en el aumento de la incidencia de esta enfermedad desde finales del año 1998 (6).
La tasa de letalidad por V. cholerae es elevada, pero es menor de un 1% en la mayoría de los casos donde existen posibilidades de acceso a un tratamiento con sales de rehidratación oral (2). La administración por vía oral de cepas de V. cholerae atenuadas por técnicas de ingeniería genética a humanos es actualmente la variante más promisoria en la búsqueda de una vacuna efectiva y barata contra el cólera, debido a que las personas expuestas a la enfermedad raramente contraen cólera después de la convalecencia (7).
En el desarrollo de estas vacunas ha influido de manera notable el advenimiento de la biotecnología moderna y el incremento sustancial en el conocimiento de la inmunidad de las mucosas; en particular; el reconocimiento de que la producción de IgA es estimulada mayormente después de una correcta inmunización oral (8).
Como resultado de un proyecto de colaboración entre el Centro Nacional de Investigaciones Científicas y el Instituto Finlay, fueron atenuadas genéticamente un grupo de cepas de V. cholerae de los serogrupos O1 y O139, candidatas a vacunas orales contra el cólera. Estas cepas fueron manipuladas genéticamente mediante el empleo de vectores suicidas y técnicas de intercambio alélico, por lo que se logró la construcción de cepas desprovistas de los genes de CT y otros genes acompañantes en el genoma del fago CTX (9). En una segunda manipulación se le inactivó a las cepas atenuadas, el gen de la hemaglutinina proteasa (hap) y en esa posición se insertó el gen de la endoglucanasa A (celA) de Clostridium thermocellum (10).
El estudio de la patogénesis del cólera ha sido posible gracias al establecimiento de biomodelos y la posibilidad de generar técnicas quirúrgicas que facilitan la localización del ileum y el ciego del tracto gastrointestinal de conejos, ocasionando una similitud de la infección provocada en el hombre (11).
Los biomodelos han permitido caracterizar candidatos vacunales vivos orales contra el cólera, donde existen diversos puntos de vista sobre la utilidad de los mismos para los ensayos de efectividad; no obstante, perros, ratas, ratones y conejos se utilizan con estos fines (11-15), pero los de mayor relevancia son ratones y conejos (11, 12).
Debido a la ausencia de un biomodelo animal que reproduzca fielmente la patogenia del cólera, el objetivo de este trabajo fue seleccionar y utilizar uno para evaluar la virulencia, capacidad de colonización y adherencia a la mucosa intestinal, en la fase de investigación básica, para seleccionar candidatos vacunales antes de su desarrollo tecnológico y clínico.
Materiales y Métodos
Todas las operaciones fueron ejecutadas ajustándose a las Buenas Prácticas de Laboratorio (BPL) establecidas en los Procedimientos Normalizados de Operaciones (PNO).
Animales
Los animales utilizados en este trabajo, procedentes del Centro Nacional para la Producción de Animales de Laboratorio (CENPALAB), fueron ratones hembras adultas de la línea consanguínea Balb/c, con sus crías o neonatos de 2 a 4 días de nacidos, con un peso vivo entre 1,5 y 2 g. Los mismos de categoría convencional y recepcionados con sus correspondientes certificados de la calidad higiénico-sanitaria y genética.
Tenencia y manejo de los animales
Los animales se mantuvieron bajo condiciones ambientales convencionales con un régimen de iluminación de 12 h luz y 12 h oscuridad. La temperatura ambiental, así como la humedad relativa fueron controladas y registradas en la mañana y durante la noche a través de un equipo de medición habilitado en cada habitación; siendo de 21 ± 2 oC y 60 ± 10%, respectivamente. El alimento fue suministrado por el CENPALAB que consistió en pienso concentrado para roedores con sus correspondientes certificados de calidad bromatológica e higiénico-sanitaria; tanto este como el agua acidulada con ácido clorhídrico, ajustada a un pH de 2,5, fueron suministradas ad libitum.
Los ratones fueron alojados en cajas modelo T2 (Policarbonato, TECNIPLAS, Italia); el encamado fue cambiado dos veces por semana, consistió en bagazo de caña de azúcar desmeollado y esterilizado en autoclave a 121 oC y 1,5 atmósferas de presión durante 25 min.
Todos los diseños experimentales y protocolos de ensayos, incluyendo los métodos de eutanasia, se sometieron a la consideración, análisis y aprobación de la Comisión de Ética de la institución, donde se respetó lo establecido por las regulaciones de seguridad biológica.
Estudios anatomopatológicos
Los estudios anatomopatológicos se realizaron por métodos convencionales, utilizando la técnica descrita para la eutanasia por Capó (16) y la necropsia según Nevalainen (17); las alteraciones macroscópicas se recogieron en protocolos establecidos al efecto; se fijaron los fragmentos seleccionados de órganos y tejidos en formalina neutra al 10% para los estudios histopatológicos y se procesaron por la técnica de inclusión y cortes en parafina, siendo coloreados con Hematoxilina Eosina (H.E).
Cepas
Las características de las cepas evaluadas en este trabajo así como los controles se resumen en la Tabla 1. Se utilizaron cepas de V. cholerae del serogrupo O1, serotipos Inaba y Ogawa, de los biotipos clásicos y El Tor y del serogrupo O139. Las cepas se conservaron a una temperatura de -70 °C en Caldo Tripitona Soya (TSB) con Leche descremada al 10% y Glicerol al 20%.
Tabla 1. Características de las cepas de V. cholerae utilizadas en los ensayos.
Nomenclatura de los genes que codifican para: ctxA, subunidad de la toxina A de la toxina colérica, ctxB, subunidad B de la toxina colérica; zot, toxina de la zona ocluida; ace, enterotoxina colérica accesoria; cep, core encoded pilus; orfU, marco de lectura abierto U; mshA, hemaglutinina sensible a manosa, thy, auxotrofía la timidina; hap, hemaglutinina/proteasa; celA, endogluconasa A y me, mutante espontáneo (los genes comprendidos en el cassette de virulencia están entre corchetes).
Análisis estadístico
Se utilizó el paquete estadístico SPSS sobre Windows, versión 10.0, así como el paquete estadístico R, versión 1.4.1, con un nivel de significación del 0,05 para todas las comparaciones. Para el análisis de la DL50 y de colonización intestinal se usó la prueba de la suma de rangos de Wilcoxon con la finalidad de determinar si existían diferencias estadísticamente significativas entre las cepas.
Experimento 1: Biomodelo de ratón neonato para evaluar la virulencia de cepas de V. cholerae
La toxina de cólera constituye uno de los factores más importantes dentro de los mecanismos de la patogenia de V. cholerae y está directamente relacionada con la virulencia. Es la responsable de los eventos determinantes en la diarrea característica de la enfermedad (18, 19).
Para comprobar la atenuación y virulencia de cada cepa se determinó la dosis letal media (DL50), según el método descrito por Ujille y colaboradores (20).
Se conformaron 5 grupos de 10 animales por dilución (para cada una de las 10 cepas), los cuales se separaron de sus madres 6 h antes de la inoculación para asegurar que sus estómagos estuvieran vacíos en el momento de la administración de las células y no hubiera nada que impidiera la adherencia de las mismas al epitelio intestinal.
La preparación de la suspensión bacteriana se realizó a partir del cultivo de las cepas en caldo de ácidos casamino y extracto de levadura (CAYE) con glucosa al 2% durante 8 h a 37 oC y 200 rpm. Se realizaron diluciones convenientes del cultivo hasta obtener una suspensión de 105 células en PBS-azul de Evans al 0,01%, el cual también cumple con el objetivo de garantizar una correcta inoculación.
A cada grupo de animales se le administró por vía oral a través de una cánula, 50 µL de las diluciones seriadas desde 10-1 hasta 10-5 del cultivo de cada una de las cepas. El grupo control recibió sólo PBS-azul de Evans al 0,01%; 6 h después se colocaron las madres y se registró el número de muertos en cada dilución a las 72 h de inoculados. El experimento se realizó por triplicado con cada cepa. Los cálculos de la DL50 se realizaron según el método descrito por Reed y Muench (21).
Para considerar que las cepas modificadas genéticamente estaban atenuadas y no virulentas se tomó como criterio la de aceptación que deben cumplir con que el valor de la DL50 fuera > 106 UFC (19).
Experimento 2: Biomodelo de ratón neonato para evaluar la capacidad colonizadora y adherencia de cepas de V. cholerae en la mucosa intestinal
La colonización intestinal en humanos por cepas de V. cholerae constituye el evento primario más importante para el desarrollo de los mecanismos de patogenicidad, además de lograr una respuesta inmune efectiva (23).
El ensayo de colonización en el biomodelo del ratón neonato es una de las pruebas que determina la potencia de cepas de V. cholerae modificadas genéticamente (24).
Los ratones fueron agrupados en un número de 10 (para cada una de las 10 cepas) y separados de sus madres 6 h antes de la inoculación, con el mismo objetivo que en el ensayo de virulencia.
La preparación de la suspensión bacteriana se realizó a partir del cultivo de las cepas en Caldo Triptona Soyas (TSB) durante 6 h a 200 rpm, en una zaranda orbital a una temperatura de 37 oC. Se realizaron diluciones convenientes del cultivo hasta obtener una suspensión de 105 células en PBS azul de Evans al 0,01%.
La prueba se desarrolló de la siguiente forma: A cada grupo de ratones se le administró 50 µL de esta suspensión. Los animales fueron sacrificados 24 h después de la inoculación. Posteriormente se realiza la extracción de los intestinos que fueron pesados en forma de mezcla (10 intestinos por cada mezcla) y lavados con el objetivo de eliminar las células que no se encontraban adheridas al epitelio intestinal. Por último se homogeneizaron en un ULTRA-TURRAX T-25 (Janke & Kunkel, IKA) a 13 500 rpm durante 15 s en PBS. El experimento se realizó por triplicado con el objetivo de garantizar la repetibilidad y confiabilidad de los resultados.
Para cuantificar el número de células adheridas se realizó conteo en placas de Agar-Tiosulfato-Citrato-Sales Biliares-Sacarosa (TCBS) a los homogenatos de los intestinos que provenían de los animales inoculados y de los controles. El índice de colonización se expresó como el número de unidades formadoras de colonias por gramo de intestino (UFC/g) (25).
El criterio de aceptación que deben cumplir las cepas de V. cholerae cuando son modificadas para esta prueba es que incrementen su concentración entre 102-103 UFC/g más, con relación al inóculo inicial (105 UFC/mL) o semejante a la cepa control (virulenta) (26).
Para evaluar la capacidad de las cepas modificadas de adherirse a la mucosa intestinal se utilizó el mismo biomodelo y metodología que en la prueba de colonización y se realizaron las tomas de muestras de intestino para estudios de identificación microbiológica de V. cholerae y para el estudio anatomopatológico mediante la inclusión y cortes en parafina por la técnica H.E, así como para el análisis inmunohistoquímico mediante la técnica de inmunopero-xidasa indirecta (27).
Se hizo una dilución 1/10 del anticuerpo policlonal anticélulas enteras de los serogrupos O1 y O139, obtenidos en conejos. Para ello se empleó un conjugado anti IgG-peroxidasa de conejo (Sigma Chemical Co., St. Louis, MO), a una dilución 1/250.
Resultados y Discusión
Experimento 1: Evaluación de la virulencia de cepas de V. cholerae
Los resultados del cálculo de la DL50 en la evaluación de la virulencia de las cepas modificadas genéticamente de primera, segunda y tercera generación difirieron significativamente con respecto a sus cepas parentales (p < 0,05), lo que nos permitió comprobar que las cepas atenuadas resultaron no virulentas, pues al comparar los valores de dosis letal media, las concentraciones celulares que provocan la muerte fueron significativamente altas en estas con relación a las concentraciones que necesitan sus correspondientes cepas parentales. Tanto es así que las cepas modificadas alcanzaron valores de dos a tres órdenes superiores (Figura 1). Estas cepas cumplieron con los criterios de aceptación para esta prueba, ya que los valores de DL50 fueron mayores de 106 UFC.
Los animales inoculados con los cultivos que se obtuvieron a partir de las cepas atenuadas sobrevivieron a la dosis de inóculo más alta, aún después de las 72 h de inoculación, lo que corrobora la atenuación de las cepas por la eliminación de sus principales determinantes de la patogenicidad (toxina de cólera y otras toxinas), las cuales están ubicadas en el genoma del fago filamentoso lisogénico CTX , presente en el cromosoma de cepas toxigénicas de V. cholerae.
Klose (23) plantea que las cepas de V. cholerae que carecen de estos genes tienen una dosis letal media en ratones neonatos mucho mayor que las cepas virulentas, lo que coincide con nuestros resultados, ya que las cepas parentales virulentas (C7258, C6706 y SG251) tienen una DL50 =103 UFC.
Experimento 2: Evaluación de la colonización y la adherencia
Independientemente de la modificación genética realizada a las cepas de V. cholerae de ambos serogrupos O1 y O139, no se afectó la capacidad de colonización del intestino de ratones neonatos, no existiendo diferencias significativas entre las cepas parentales y las modificadas genéticamente (p > 0,05) (Figura 2).
El hecho de que las cepas parentales y las atenuadas presenten índices de colonización similares coincide con lo reportado por Silva y colaboradores (28), cuando señaló que en la transformación genética realizada no se modifican los genes que regulan la capacidad de colonización y adherencia en el intestino del modelo de ratón neonato.
La adherencia, al igual que la colonización, es un factor crítico en la patogénesis del cólera. A pesar de que se ha investigado mucho sobre el mecanismo de colonización de V. cholerae, todavía se conoce poco acerca de cuáles son los atributos que facilitan la colonización y la adherencia de este microorganismo, aunque existe consenso en que es un proceso multifactorial en el que intervienen varios compuestos y que están favorecidos por determinadas características ambientales (29, 30).
Adicionalmente a las cepas atenuadas de segunda generación (638, 1333 y L911) se les inactivó el gen hap (que codifica para la hemaglutinina proteasa, una enzima que desempeña un papel importante en la colonización intestinal) y en su lugar se le insertó el gen celA, lo que no afectó la capacidad de colonización de las cepas. De hecho, la inactivación del gen hap incrementó la colonización en los ratones neonatos.
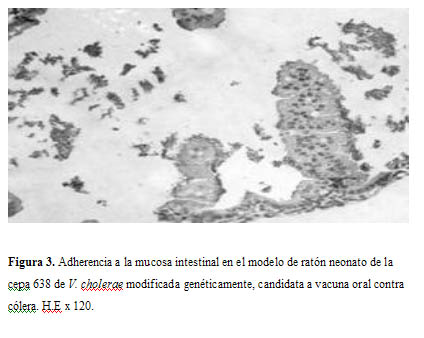
A la observación microscópica de los cortes histológicos realizados a la mucosa intestinal de ratones inoculados por vía oral con cepas de V. cholerae virulentas y genéticamente modificadas, mediante la técnica de inmunoperoxidasa, utilizando anticuerpos policlonales anticélulas enteras de los serogrupos O1 y O139, se evidenció una reacción positiva frente a los anticuerpos específicos de ambos serogrupos, apreciándose reacciones positivas en la luz del órgano y encima de la capa superficial del epitelio intestinal, con la presencia de abundantes bacterias adheridas a la superficie de las células del intestino delgado, siendo estos resultados similares a los alcanzados por Kirn y Taylor (11).
Los estudios histopatológicos guardan relación con los aislamientos micro-biológicos obtenidos en los cultivos realizados, en los que se identificó por bioquímica y serología la presencia de V. cholerae de los serogrupos O1 y O139, coincidiendo con los resultados obtenidos por Cedré y colaboradores (26).
Conclusión
El biomodelo aplicado del ratón neonato permitió la evaluación y selección de cepas candidatas para vacunas vivas orales contra el cólera humano, en cuanto a virulencia, colonización y adherencia a la mucosa intestinal.
Referencias
1. Bajaj JK, Baradker VP, Joshi SG, Damle AS, Kayararte RP, Deshmukh AB. Epidemiology of cholera a five year study. J Commun Dis 2001; 33(4):5-282.
2. CDC/NCID.OPS. Métodos de laboratorio para el diagnóstico de Vibrio cholerae. En Programa especial de publicaciones; 1994.
3. Waldor MK, Mekalanos JJ. Lysogenic conversion by a filamentous phage encoding cholera toxin. Science 1996; 272:1910-14.
4. WHO. World Health Organization. Alerta y Respuesta ante Epidemias y Pandemias (EPR). [en línea] junio 2006. Disponible en:
http://www.who.int/csr/don/2006_06_21a/es/index.html [Consultado: 12 abril 2007].
5. ISID. International Society for Infectious Diseases. Cholera, diarrhea & dysentery. [en línea] ProMED-mail 2007 (21):1804. Disponible en:
http://www.promedmail.org/pls/promed/f?p=2400:1001:2329690040162723291::NO::F2400_P1001_BACK_PAGE,F2400_P1001_PUB_MAIL_ID:1000,37699. [Consultado: 4 junio 2007].
6. Rodríguez R. Reporte Técnico de Vigilancia (versión ASCII) 2000; 5.
7. WHO. World Health Organization. Meeting on the potential role of new cholera vaccine in the prevention and control of cholera outbreaks during acute emergencies. Document CDR/GPV/95.1, 1995. World Health Organization, Geneva, Switzerland.
8. Levine MM and Tacket Co. Recombinant live cholera vaccines. En: Wachsmuth IK, Blake PA, Olsvik O, eds. Vibrio cholerae and cholera: molecular and global perspectives. 1994; Washington DC. American Society for Microbiology: 395-414.
9. Benítez JA, Silva A, Rodríguez B, Fando R, Campos J, Robert A, et al. Genetic manipulation of Vibrio cholerae for vaccine development: construction of live attenuated El Tor candidate vaccine strains. Archives of Medical Research 1996; 27(3):275-283.
10. Robert A, Silva A, Benítez JA, Rodríguez BL, Fando R, Campos J, et al. Taggin a Vibrio cholerae El Tor candidate vaccine strain by disruption of its hemagglutinin protease gene using a novel reporter enzime, Clostridium thermocellone endogluconase A. Vaccine 1996; 14:1517-22.
11. Kirn TJ and Taylor RK. TcpF is a soluble colonization factor and protective antigen secreted by El Tor and classical O1 and O139 Vibrio cholerae serogroups. Infect Immun 2005; 73(8):4461-70.
12. Liang W, Want S, Yu F, Zhang L, Qi G, Liu Y., et al. Construction and evaluation of a safe, live, oral Vibrio cholerae vaccine candidate, IEM108. Infect Immun 2003; 71(10):5498-504.
13. Sack RB and Carpenter CJ. Experimental canine cholera. I. Development of the model. J Infect Dis. 1969; 119:138-149.
14. Sack RB, Johnson J, Pierce NF, Keren DF, Yardley JH. Challenge of dogs with live enterotoxigenic Escherichia coli and effects of repeated challenges on fluid secretion in jejunal Thiry-Vella loops. J Infect Dis. 1976; 134(1):15-24.
15. Sifontes S. Consideraciones éticas en el desarrollo de modelos animales para el cólera. VI Congreso Internacional de Ciencias Veterinarias. (10 al 13 de abril de 2007. Palacio de Convenciones, La Habana. Cuba.) Programa Científico. 66.
16. Capó MA. Bioética Animal: Desarrollo de un Concepto. Animales de Experimentación. La Revista Hispanoamericana 1999; 5(2):30-40.
17. Nevalainen T, Berge E, Gallix P, Jilge B, Melloni E, et al. FELASA guidelines for education of specialists in laboratory animal science (Category D). Laboratory Animals 1999; 33: 1-15.
18. García L y Benítez J. Hacia el desarrollo de una vacuna eficaz contra el cólera. VacciMonitor 1996;5(2):2-6.
19. Finkelstein RA. Cholera enterotoxin (choleragen): a historical perspective. En: Barua D, Greenough III WB, editores. Cholera. Edition. New York: Plenum Medical Book Company; 1992.
20. Heidelberg JF, Eisen JA, Nelson WC, Clayton RA, Gwin ML, Dodson RJ, et al. DNA sequence of both chromosomes of the cholera pathogen Vibrio cholerae. Nature 2000; 406:477-483.
21. Ujiiye A, Nakatomi M, Utsunomiya A, Mitsui K. Experimental cholera mice. First report on the oral infection. Trop Med. 1968; 10:65-71.
22. Reed LJ and Muench H. A simple method for estimating fifty percent endopoints. Am. J. Hyg. 1938; 27:493-497.
23. Klose KE. The suckling mouse model of cholera. Trends Microbiol 2000; 8(4):189-91.
24. Pearson GD, Woods A, Chiang SL, Mekalanos JJ. CTX genetic element encodes site-especific recombination system and an intestinal colonization factor. Proc Natl Acad Sci USA. 1993; 90:3750-54.
25. Brandtzaeng P. Role of secretory antibodies in the defence against infections. Int J Med Microbiol 2003; 293(1):3-15.
26. Cedré B, García LG, García HM, Fariñas M, Talavera A and Infante JF. Intestinal colonization of the infant mouse model by attenuated and virulent Vibrio cholerae strains. Archives of Medical Research 1998; 29(3):231-234.
27. Vacca LL. Laboratory Manual of histochemistry. En: Raven Press Books, editors. ed. New York; 1985; p. 27-32, 118-119, 541-549.
28. Silva AJ, Pham K, Benítez JA. Haemagglutining/portease expresión and mucin gel penetration in El Tor biotype Vibrio cholerae. Microbiology 2003; 149(7):1883-91.
29. Chiavelli DA, Marsh JW, Taylor RK. The mannose-sensitive hemagglutinin of Vibrio cholerae promotes adherence to zooplankton. Appl Environ Microbiol 2001; 67(7):3220-25.
30. Merrell DS, Hava DL, Camilli A. Identification of novel involved in colonization and acid tolerance of Vibrio cholerae. Mol Microbiol 2002; 43(6):1471-91.
Recibido: Febrero de 2008
Aceptado: Marzo de 2008


{kind=link}
{kind=link}
{kind=link}
{kind=link}