










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkVaccimonitor
versión impresa ISSN 1025-028Xversión On-line ISSN 1025-0298
Vaccimonitor v.17 n.1 Ciudad de la Habana ene.-abr. 2008
Interacción del componente pertussis de células completas con los antígenos tetánico, Haemophilus influenzae tipo B y hepatitis B en ensayos de potencia para vacunas combinadas
Interaction of the whole-cell pertussis component with the the tetanus antigens, type b Haemophilus influenzae and hepatitis B in potency assays for combined vaccines
Mario Landys1, Dr. Juan Miguel Figueroa2, Ana Lara2, Vicente Perdomo2.
1 Instituto Finlay, Centro de Investigación-Producción de Vacunas. Ave. 27 No. 19805 e/198 y 202.
La Lisa, Ciudad de La Habana, Cuba.
2 Centro para el Control Estatal de la Calidad de los Medicamentos (CECMED). Calle 200, No 1706 entre 17 y 19. Siboney, Playa, Ciudad de La Habana, Cuba.
Correo electrónico: mlandys@finlay.edu.cu
RESUMEN
Las vacunas combinadas permiten un grado de aceptación mayor entre la población, dado que se requiere de menos inmunizaciones para proteger contra las enfermedades infecciosas. Sin embargo, esto ha generado nuevos retos, ya que se han reportado numerosas interacciones entre los diferentes antígenos que conforman estas vacunas. El propósito del presente trabajo fue evaluar la interferencia potencial del componente pertussis de células completas sobre los ensayos para determinar la actividad biológica de otros antígenos como toxoide tetánico, Haemophilus influenzae tipo b (Hib) y hepatitis B. Para ello se estudiaron mediante ensayos de potencia vacunas combinadas que contenían estos antígenos y se compararon con vacunas monovalentes. A su vez se prepararon adyuvaciones experimentales cuya composición permitió estimar adecuadamente la extensión y naturaleza de la interacción entre componentes. Se obtuvo que el componente pertussis incrementó significativamente la actividad biológica de Hib y tétanos, aunque esto puede depender mucho del modelo animal y el diseño experimental utilizado. En cuanto al antígeno de hepatitis B se demostró que pertussis inhibía la potencia de este antígeno, tanto in vitro como in vivo, aunque por mecanismos diferentes y de forma no significativa.
Palabras clave: Componente pertussis de células completas, interacciones, potencia de toxoide tetánico, antígeno de hepatitis B y Hib
ABSTRACT
Combined vaccines have a higher level of acceptance in the population because fewer immunisations are required for protection against infectious diseases. Nevertheless, new challenges have appeared due to the report of some interference among the antigens included in such vaccines. The aim of this paper was to evaluate whole-cell Pertussis (wP) interference in potency tests with other antigens like tetanus, Haemophilus influenzae type b (Hib) and hepatitis B. For that purpose, we evaluated combined vaccines and compared them with the respective monovalent vaccines. Likewise, we prepared experimental formulations with different antigen combinations to evaluate the extent and nature of the interference caused by wP. We obtained a significant increase of the tetanus and Hib biological activities when both antigens are combined with wP, although it depended on the animal model and the experimental design used. On the other hand, wP decreased both the in vivo and in vitro potency of HB in a non-significant way.
Keywords: Whole-cell pertussis, interference, hepatitis B, tetanus toxoid, hepatitis B and Hib antigen potency.
Introducción
El desarrollo y uso de vacunas combinadas ha estado dirigido a incrementar la cobertura de vacunación en todo el mundo, así como a elevar el acceso de la población a estos importantes productos inmunobiológicos. El hecho de que se requiera un número menor de inyecciones para proteger contra más enfermedades aumenta el nivel de aceptación de los programas nacionales de inmunización, no sólo entre la población sino también entre el personal de salud. Al mismo tiempo ello contribuye al ahorro de los presupuestos destinados a la salud pública en los diferentes países (1).
El concepto de vacunas combinadas no es nuevo, partiendo de que durante más de dos décadas se han empleado en el mercado combinaciones de antígenos bacterianos inacti-vados como los toxoides tétanico, diftérico y células completas inactivadas de Bordetella pertussis (DPwT), y virus vivos atenuados de rubéola, sarampión y parotiditis, a las cuales se han sumando combinaciones multivalentes de vacunas pneumo y meningocócicas con resultados altamente satisfactorios.Recientemente, a las habituales combinaciones de toxoide tetánico y diftérico y antígenos de pertussis acelular o de células completas se han ido adicionando otros componentes como las vacunas Haemophilus influenzae tipo b (Hib), antipoliomelitíca inactivada (IPV), antihepatitis B (HB) y antihepatitis A, además de otras vacunas en desarrollo como los conjugados polisacárido-proteína pneumo y meningocócicos, y las combinaciones de vacunas contra infecciones entéricas como el cólera, la fiebre tifoidea y las producidas por Shigella y rotavirus (2).
Sin embargo, contrario a las expectativas generadas por las nuevas combinaciones de vacunas, han surgido algunas complicaciones técnicas y científicas. En esta nueva generación de vacunas combinadas la presencia de más de un componente frecuentemente causa una interacción que lleva a la disminución o al incremento de la actividad biológica o a la transformación de una o más propiedades de algunos de los componentes de la combinación en relación con su comportamiento cuando son administrados solos. Los principales problemas asociados al desarrollo de vacunas combinadas pueden resumirse en el incremento de la reactogenicidad (por excesiva carga de endotoxina, carga excesiva de toxoides, efectos sinérgicos o aditivos, desplazamiento del antígeno del adyuvante o estimulación de respuestas inmunes excesivas o inapropiadas), la reducción de la inmunogenicidad, la excesiva variación lote a lote producto de complejas interacciones entre los componentes, así como estabilidad subóptima (1,2).
Tales interacciones son generalmente de naturaleza inmunológica, aunque también pueden existir interacciones químicas y físicas entre los diferentes antígenos. Las interacciones inmunológicas incluyen la competición antigénica, la supresión epitópica específica, el efecto adyuvante y la interacción adversa con el adyuvante. Es por ello que se recomienda que cada combinación tiene que ser desarrollada y estudiada individualmente en términos de calidad, seguridad, estabilidad, tolerabilidad clínica y eficacia e inmunogenicidad (2).
Entre las interacciones más conocidas en vacunas combinadas tenemos las que se producen entre los tres componentes de la vacuna trivalente antipoliomielítica oral, la reducción de la respuesta inmune al antígeno de la parotiditis inducida por sarampión en la vacuna triple viral, el incremento del contenido de endotoxinas en las combinaciones de vacunas DTP y antitifoídica y las diversas interacciones que tienen lugar en vacunas que contienen DTP y Hib (2).
En la clínica se han reportado algunas de estas interacciones producto de la inmunización con vacunas combinadas, en particular las relativas a combinaciones de vacunas Hib y DTP, tanto con pertussis acelular como de células completas, donde se ha observado que existe una marcada diferencia en los títulos anti-PRP (polirribosil ribitol fosfato), polisacárido utilizado como antígeno en combinaciones de vacunas Hib y DTP con relación a las vacunas monovalentes (3-6), aunque a su vez otros autores niegan que existan interferencias significativas entre dichos componentes (7-8). De cualquier modo, pareciera que tales interferencias dependen del número de dosis administradas en los niños (6).
Otro panorama más complejo se presenta en los ensayos de control para estas vacunas combinadas. Shams y colaboradores (9) evaluaron de forma experimental, tanto en ratones como en curieles, las interacciones que tienen lugar entre vacunas Hib conjugadas y una vacuna tetravalente DPaT-IPV. En dicho estudio la administración concomitante de vacuna Hib conjugada a toxoide tetánico y DPaT-IPV produjo una disminución significativa de la respuesta de anticuerpos anti-PRP en el modelo ratón, mientras que en curieles no hubo variación que resultara significativa, lo cual indica que el efecto de supresión antigénica pudiera deberse a la mayor simplicidad del sistema inmune del ratón. Más recientemente un documento del Departamento Europeo de Calidad de los Medicamentos (EDQM) reportaba un incremento de la respuesta al componente Hib producida en combinaciones con DTP, en comparación con la respuesta al evaluar vacunas Hib conjugadas exclusivamente. El incremento de la respuesta al componente tetánico en estas combinaciones fue igualmente reportado por la mayor parte de los Laboratorios Nacionales de Control Europeos (OMCL) (1). Una buena parte de estos efectos se atribuyen a la acción del adyuvante, bien sean las sales de fosfato o el gel de hidróxido de aluminio, y muy particularmente al componente de células completas de Pertussis (1-2).
Tomando en consideración estos antecedentes y el propio incremento de la fabricación de combinaciones de vacunas en Cuba, las cuales requieren de una rigurosa caracterización que incluya la evaluación de las potenciales interferencias entre los diferentes antígenos, el Laboratorio Nacional de Control de la Autoridad Reguladora de Medicamentos cubana (CECMED) se propuso como objetivo fundamental en este trabajo evaluar las interacciones del componente Pertussis de células completas con el Toxoide Tetánico y los antígenos Hib y Hepatitis B en ensayos biológicos para determinar potencia en las vacunas combinadas producidas en nuestro país.
Materiales y Métodos
Muestras
Se utilizaron muestras de vacunas combinadas: Trivac HB (DTP-HB) y Heberpenta (DTP-HB+Hib), suministradas por el Centro de Ingeniería Genética y Biotecnología (CIGB). Este centro proporcionó igualmente las vacunas monovalentes contra la hepatitis B (Heberbiovac HB ) y el Haemophilus influenzae tipo b (Quím.-Hib ), esta última la primera vacuna (conjugada a toxoide tetánico) cuyo antígeno polisacarídico se produce por vía sintética. Por su parte, el Instituto Finlay suministró las vacunas vax-TET (antitetánica), Va-DIFTET (antidiftérica y antitetánica) y DTP . Todas las vacunas del estudio, con excepción de Quimi-Hib que no tiene adyuvante, están adsorbidas en gel de hidróxido de aluminio. Los lotes de vacunas fueron liberados tanto por los respectivos fabricantes como por el CECMED.
Los Ingredientes Farmacéuticos Activos para la preparación de las adyuvaciones experimentales fueron igualmente suministrados por el CIGB y el Instituto Finlay.
Patrones y reactivos
Para la titulación de sueros de conejos para anticuerpos anti-PRP se utilizó el suero patrón de la FDA (Food and Drug Administration), también conocido por suero de Frasch, con actividad de 60,9 g/mL de IgG. Para la estimación de la potencia en vacunas antihepatitis B se empleó un material de referencia interno de vacuna, calibrado contra el patrón de trabajo previo. La asignación de títulos de anticuerpos antitetánicos se realizó utilizando un suero hiperinmune, al que se le asignaron 100 unidades de antitoxina tetánica por mL.
Los reactivos biológicos (Conjugados y Sustratos) para realizar el ELISA se adquirieron de fabricantes reconocidos y certificados (SIGMA y OXOID). Para la determinación de la potencia in vivo / in vitro del componente HB se adquirió el kit comercial Hepanostika anti-AgsHb (Organon Teknika, Holanda). En el caso del ELISA de inhibición utilizado para la determinación de la potencia in vitro de HB se empleó una inmunoglobulina antihepatitis B producido por el Centro de Hemoderivados (Empresa "Adalberto Pesant"), Cuba, a partir del suero de personas sanas inmunizadas con la vacuna Heberbiovac HB (CIGB). Todos los reactivos químicos utilizados en los inmunoensayos son de calidad p.a. (puro para análisis) e igualmente fueron adquiridos de proveedores certificados (Merck, Alemania).
Preparación de adyuvaciones experimentales de vacunas
Se prepararon las siguientes adyuvaciones experimentales: Toxoide Tetánico (TT), DT, DTwP, HB, wP-HB, DTwP-HB, las cuales se combinaron con el componente PRP-TT (Hib) que no es adyuvado. Las correspondientes adyuvacio-nes se realizaron siguiendo el mismo procedimiento utilizado por el CIGB para la adsorción de antígenos durante la fabricación: El gel de hidróxido de aluminio fue preparado en solución salina y los antígenos fueron adsorbidos por adición de cada antígeno de forma lenta, con permanente agitación. Con el fin de minimizar la influencia del antígeno de pertussis, este fue degradado antes de conformar algunas de las muestras experimentales, lo cual también se realizó con muestras adyuvadas del componente HB. En ambos casos se incubaron las muestras a 70 C durante una semana y luego se realizó la adsorción y combinación con otros antígenos en los casos que procedía. Todas las muestras fueron conservadas entre 2 y 8 C hasta su uso.
Animales
Para la evaluación de la actividad biológica del componente Hib se utilizaron conejos blancos raza Nueva Zelanda, hembras, con peso entre 2 y 3 \kg. Para la determinación de la actividad biológica de los componentes TT y HB se utilizaron ratones Balb/C hembras, con un peso entre 16 y 18 g. Todos los animales fueron adquiridos a través del Centro Nacional para la Producción de Animales de Laboratorio (CENPALAB), Cuba.
Determinación de la actividad biológica del componente PRP (Hib) en conejos
Las muestras de vacunas que contenían PRP fueron inoculadas a razón de 5 conejos por lote, bajo un esquema de dos inmunizaciones (0 y 14 días) con 0,5 mL. Los animales fueron desangrados a los 21 días de la primera inmunización por vía axilar bajo anestesia y los sueros se obtuvieron por centrifugación a 2 500 rpm durante 10 min, se mezclaron en partes iguales y se conservaron a -70 C hasta su uso.
Para la determinación de los títulos anti-PRP se realizó un ELISA antiespecie, desarrollado en nuestro laboratorio (10). Brevemente: se recubrieron las placas de microelisa con PRP acoplado a albúmina de suero humano en PBS durante 2 h a 37 C. Posteriormente se añadió como bloqueo una solución de seroalbúmina de suero bovino (BSA) al 1% en agua destilada y se incubó a 37 C durante media h. Concluido este tiempo se preparó una curva de calibración del suero estándar de FDA y se hicieron diluciones doble seriadas de las mezclas de suero para aplicar en la placa. Una vez finalizado el tiempo de incubación de las muestras, se aplicó el conjugado de IgG anti-conejo acoplado a peroxidasa de rábano picante a una dilución 1/10000. Como sustrato se utilizó ortofenilendiamina (OPD) en tampón fosfato citrato + peróxido de hidrógeno y se incubó a temperatura ambiente. Media hora más tarde, la reacción se detuvo con solución de ácido sulfúrico (2 mmol/L) y la absorbancia de los pocillos se lee a 492 nm en un lector de placas. Los resultados se procesan por el programa CDC para ELISA y los títulos se reportan en comparación con el suero estándar de la FDA. Los sueros deben tener una actividad biológica superior a los 10 g/mL.
Determinación de la actividad biológica del componente Tétano (TT) en ratones
Para la evaluación del componente TT en vacunas se siguió un esquema de dos inmunizaciones (0 y 28 días), con desangrado a los 42 días. Los ratones fueron desangrados por vía retroorbital y el suero se obtuvo por el mismo procedimiento descrito para Hib.
La evaluación de los títulos anti-TT se realizó por un ELISA desarrollado en el laboratorio (10). Las placas fueron incubadas toda la noche a 4 C con TT a 0,5 s.f. /Mª en 100 Mohs/L de tampón carbonato / bicarbonato (pH 9,6). El resto de los pasos del ELISA fueron muy similares al descrito para Hib, sólo que utilizando un suero hiperinmune antitetánico y conjugado anti-ratón. El criterio de aceptación utilizado fue el siguiente: actividad antitetánica en los sueros 2 UI de Antitoxina tetánica/mL.
Potencia in Vitro del componente HB
Se realizó mediante un ELISA de inhibición desarrollado en el laboratorio (11). Las muestras de vacuna y la referencia fueron diluidas en PBS que contenía albúmina de suero bovino al 1% (diluciones seriadas 1:2) e incubadas con una cantidad fija de inmunoglobulina anta-AGSO, a concentración neutralizante, a 37 C durante 1 h. Transcurrido el tiempo de incubación se centrifugó para eliminar el adyuvante y se realizó el ELISA para determinar los anticuerpos anti-AgsHb remanentes utilizando el kit Hepanostika. Las muestras se evaluaron por el método de líneas paralelas (12). La potencia relativa debe ser 0,75 en relación con la vacuna de referencia.
Potencia in vivo para el componente HB
La potencia del componente HB se evaluó a través del clásico ensayo de inmunogenicidad en ratones descrito por la OMS (13). Las muestras de vacuna y la referencia fueron diluidas en PBS (5 diluciones seriadas 1:2). Grupos de 10 ratones por dilución fueron inmunizados por vía intraperitoneal. El suero fue obtenido por punción retroorbital tras 28 días de observación de los animales. Posteriormente las muestras se evaluaron por el mismo ELISA para determinación de anticuerpos anti-AgsHB descrito para la potencia in vitro. Las muestras se evalúan por el método de las líneas paralelas Probit (12). La potencia relativa debe ser 0,5 en relación con la vacuna de referencia.
Evaluación de la interacción producida por wP
Todas las vacunas combinadas y adyuvaciones experimentales fueron evaluadas por los métodos de ensayo antes citados y comparados con las formulaciones monovalentes. En tales evaluaciones se incluyeron muestras experimentales que contenían antígeno de pertussis degradado para demostrar si este antígeno era el responsable o no de las interacciones observadas en los métodos de control.
En el caso del componente HB se realizaron adicionalmente dos estudios: uno de correlación entre los ensayos de potencia in vivo/in vitro y otro en el cual se evaluó si existía reacción cruzada entre el antígeno de pertussis y la inmunoglobulina antihepatitis B utilizada en el ELISA de inhibición.
Cada adyuvación fue evaluada en seis determinaciones independientes en el caso del ELISA de inhibición para hepatitis B y tres ensayos independientes para todas las determinaciones in vivo.
Procesamiento estadístico
El estudio de correlación fue evaluado utilizando los paquetes estadísticos de los Programas Excel y Statgraphics. La relación entre cada par de variables fue realizada por ajuste a un modelo lineal. La correlación de Pearson (12) fue calculada y su significación estadística se estimó a partir del valor de P. Se consideró que los valores de P por debajo de 0,05 indicaban una correlación estadísticamente significativa y diferente de cero, en un intervalo de confianza del 95%. Los resultados derivados de la comparación de las adyuvaciones fueron analizados para comprobar una distribución normal y la significación estadística de las diferencias entre medias fue determinada por el test de Student. Las diferencias fueron consideradas significativas para P 0,05 (12).
Resultados y Discusión
La inclusión de varias vacunas combinadas en el Programa Nacional de Inmunización de Cuba, los reportes de interacciones entre antígenos que conforman tales combinaciones y algunos antecedentes generados durante la estandarización y validación de ensayos de potencia en nuestro laboratorio, particularmente en aquellas vacunas que contienen wP, nos llevaron a evaluar el papel de este antígeno en la variación de la respuesta inmune producida por otros antígenos administrados de forma concomitante en relación con el comportamiento de los mismos en formulaciones monovalentes.
La Tabla 1 muestra el comportamiento de los títulos anti-PRP y anti-TT y de la potencia in vivo / in vitro del componente HB en vacunas combinadas en comparación con vacunas que contienen un solo antígeno. En el caso de la actividad biológica del componente Hib, se obtuvieron incrementos estadísticamente significativos en las vacunas que contenían PRP y wP de forma simultánea en comparación con la vacuna Hib monovalente, no ocurriendo así en otras combinaciones como la de HB+Hib.
Tabla 1. Actividad biológica de los componentes individuales HB, Hib y TT en vacunas combinadas en relación con las formulaciones monovalentes.
Algo similar ocurrió con los títulos anti-TT, los cuales tuvieron un incremento en vacunas combinadas cuya formulación incluía TT y wP en relación con la vacuna antitetánica, sólo que no en todos los casos este aumento fue significativo. Por ejemplo, la vacuna DTP tuvo un incremento de aproximadamente el doble de la vacuna TT, pero este no fue estadísticamente significativo, a diferencia de las que contenían D, T, wP junto al componente HB o Hib, cuyos títulos sí mostraron diferencias significativas en relación con la vacuna monovalente.
Ambos resultados indican que el antígeno wP contribuye al incremento de los títulos de los componentes Hib y TT cuando se encuentran estos en la misma formulación, al ser evaluados por los modelos experimentales utilizados en este estudio. A fin de verificar si verdaderamente es el componente wP, y no otro antígeno, el responsable de la interferencia observada en los ensayos de control, se prepararon y evaluaron diferentes adyuvaciones experimen-tales que permitieron discriminar si el efecto producido se debía al componente wP o no.
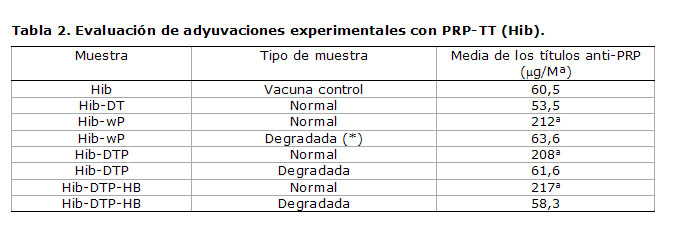
La Tabla 2 confirma que el incremento de la actividad biológica del componente Hib en vacunas combinadas se debe al efecto inducido por wP. En todas las adyuvaciones en las cuales se combinó wP y PRP se obtuvo un aumento significativo de los títulos anti-PRP, que existió cuando la adyuvación se realizó con el antígeno wP previamente degradado. En estos últimos casos los valores de actividad biológica fueron comparables, casi similares a los de la vacuna control, demostrando que en tales muestras sólo el componente Hib produce anticuerpos anti-PRP de forma equivalente a la vacuna monovalente. Resulta conveniente esclarecer que el componente wP no es capaz de producir anticuerpos anti-PRP por él mismo, sino que esto se produce por un efecto adyuvante que refuerza la producción de anticuerpos anti-Hib por parte de las células plasmáticas (1,2). Está reportado que el componente HB puede igualmente contribuir a este incremento de los títulos anti-PRP (2), pero su influencia no tiene la relevancia de la producida por wP.
Tabla 2. Evaluación de adyuvaciones experimentales con PRP-TT (Hib).
Los resultados están expresados como títulos en relación con el suero de referencia de la FDA (suero de Frasch). La comparación de las diferencias entre las adyuvaciones y la vacuna control se realizó por un t de Student. Se consideró que existían diferencias significativas cuando P 0,05.
a Representa las diferencias significativas en relación con la vacuna control Hib
(*) Siempre que aparezca se refiere a la degradación del componente wP (solamente) antes de la adyuvación experimental y la combinación con otros antígenos.
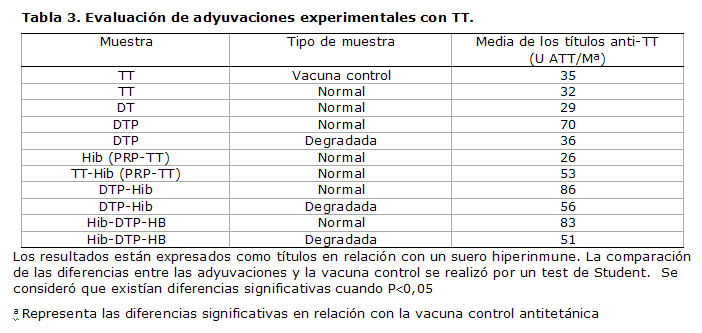
La Tabla 3 confirma que wP es también el responsable del incremento de los títulos anti-TT en las adyuvaciones que contenían ambos antígenos por el mismo efecto mitógeno de células plasmáticas antes mencionado, lo cual queda reforzado por la obtención de resultados menores cuando se degrada este componente, eliminando su influencia en la formulación. Es conocido que la especificación de la potencia del componente antitetánico en las vacunas DwPT es superior a la establecida para la vacuna antitetánica monovalente, según los requerimientos de la Organización Mundial de la Salud (14).
Sin embargo, de esta Tabla se derivan consideraciones adicionales. Desde la Tabla 1 se pudo observar que la vacuna monovalente Hib tenía una actividad antitetánica notable, comparable, aunque ligeramente menor, con la de la vacuna vax-TET®. Ello se debe a que la vacuna Quimi-Hib posee como proteína transportadora (carrier) precisamente TT a fin de incrementar la pobre inmunogenicidad del PRP (15), razón por la cual aparecen valores de incremento notables, aunque no significativos en el estudio de las adyuvaciones, cuando wP y Hib están combinados con TT. Incluso en aquellas adyuvaciones donde se degradó previamente wP, pero está presente el componente PRP-TT junto al TT, los títulos anti-TT son superiores a los de la vacuna control. Existen incluso autores que consideran que quizás sea innecesario incorporar TT como antígeno en vacunas combinadas que contengan PRP conjugado a TT (1, 2).
Sin embargo, no siempre estas interacciones halladas en métodos de potencia tienen correspondencia con los resultados en ensayos clínicos, de modo que, a pesar de las diferencias significativas encontradas en uno y otro caso y del indiscutible efecto inducido por wP, es necesario tomar los resultados obtenidos con precaución. La influencia del modelo animal y del esquema de inmunización, para sólo tomar dos ejemplos, pueden tener impacto en la estimación de la naturaleza y la extensión de la interferencia producida por el antígeno wP. Tanto para la evaluación del componente Hib como para el TT, se emplearon esquemas de dos dosis, que favorecen la respuesta inmune, pero que no resultan del todo adecuados para ensayos de control. En cuanto al modelo animal, está descrito que no existe un modelo único y del todo relevante para la evaluación de la respuesta contra el Hib, y la utilización de conejos posee un valor limitado. Con relació al empleo de ratones para tétanos, esto es válido mientras la evaluación se mantenga por debajo de tres antígenos, pues la simplicidad del sistema inmune de esta especie puede producir resultados de valor dudoso (16, 17).
Es por ello que resulta necesario explorar nuevos diseños experimentales y modelos animales (el curiel, entre ellos) (17) para conocer si la interacción inducida por wP en vacunas que contienen Hib y TT tiene el nivel de significación obtenido por nosotros en este trabajo.
Tabla 3. Evaluación de adyuvaciones experimentales con TT.
Un panorama distinto se presenta con la evaluación de la interferencia de wP sobre el componente hepatitis B: no existen reportes sobre la estimación de este efecto relacionado con la actividad biológica del antígeno de superficie de la hepatitis B en vacunas combinadas, se dispone de métodos reconocidos y correlacionados con los resultados en ensayos clínicos para evaluar la potencia de este componente de forma confiable y la interferencia producida por wP sobre HB, a diferencia de la observada para Hib y TT, es del tipo inhibición.
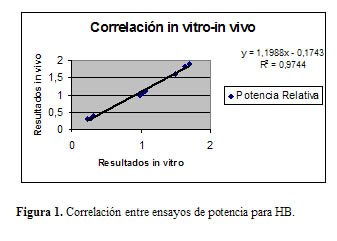
Partiendo de que nuestro laboratorio disponía de dos metodologías, una in vivo y otra in vitro, para estimar la potencia del componente HB, decidimos realizar un estudio de correlación para ver si la interferencia de wP era detectada por ambos métodos de forma equivalente. La Figura 1 muestra un coeficiente de correlación altamente satisfactorio (r = 0,997, P 0,05), el cual ya habíamos obtenido, pero sólo para vacunas monovalentes (11, 18).
Esta vez se demostró que la detección de la interferencia producida por wP era equivalente, a pesar de tratarse de métodos con principios diferentes.
Se diferencian claramente tres poblaciones: los puntos más a la derecha corresponden a las formulaciones HB monovalentes, los del centro a adyuvaciones de vacunas combinadas conteniendo wP y los de la izquierda corresponden a muestras de HB degradadas para no cumplir con las especificaciones de ninguno de los dos métodos. Como resulta evidente, wP produce disminución de la potencia in vivo / in vitro de HB (muestras del centro), pero no al punto de afectar el cumplimiento de las especifi-caciones establecidas por ambos ensayos, que son capaces de discriminar claramente entre muestras subestándar (las de la izquierda) y muestras con el nivel de interferencia inducido por wP.
Como se observa, la naturaleza de la interferencia de wP en cada uno de estos dos ensayos es bastante diferente. En el caso del ensayo de potencia in vivo, lejos de la capacidad adyuvante o activadora del complemento, el componente wP parece actuar sobre las células B específicas para HB por un mecanismo de competencia antigénica (2).
La interferencia sobre el ELISA de inhibición tiene una naturaleza bien diferente. En nuestra opinión, pensamos que podría tratarse de un bloqueo parcial de la unión entre el componente HB y la inmunoglobulina anti-HB, evitando la neutralización que se necesita para la estimación de potencia en nuestro método de potencia in vitro. Esto incrementaría el nivel de anticuerpos anti-HB sin neutralizar y disminuiría la potencia relativa de vacunas que contienen wP. Otro posible efecto sería una unión directa al antígeno de superficie de HB que recubre las placas de ELISA.
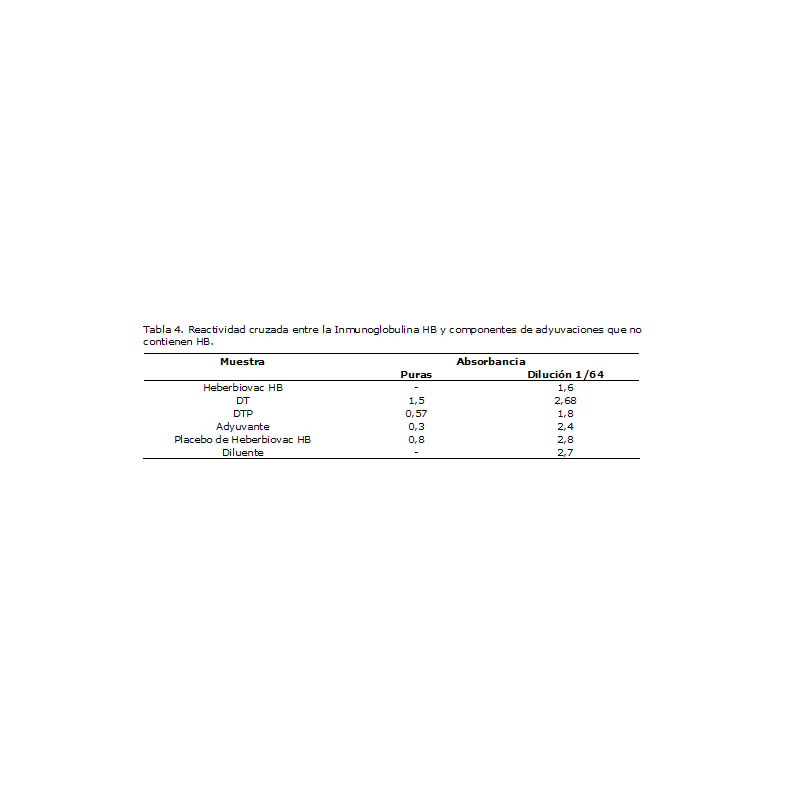
Tratando de identificar el mecanismo a través del cual el antígeno pertussis interfería en la potencia del componente HB, se evaluó la reactividad cruzada en relación con la inmunoglobulina HB. Como se observa en la Tabla 4, sólo la adyuvación que contenía wP fue capaz de mostrar una absorbancia comparable a la de la vacuna control a la mayor dilución de ensayo (1/64), efecto completamente inherente a wP si tomamos en cuenta que no sucedió lo mismo con la adyuvación DT. De igual forma, pudiera pensarse que en esta interferencia puede existir una contribución del adyuvante, pero resulta evidente de nuestros resultados que la interacción con el aluminio se produce solamente en muestras puras y desaparece por completo a la dilución de ensayo.
Tabla 4. Reactividad cruzada entre la Inmunoglobulina HB y componentes de adyuvaciones que no contienen HB.
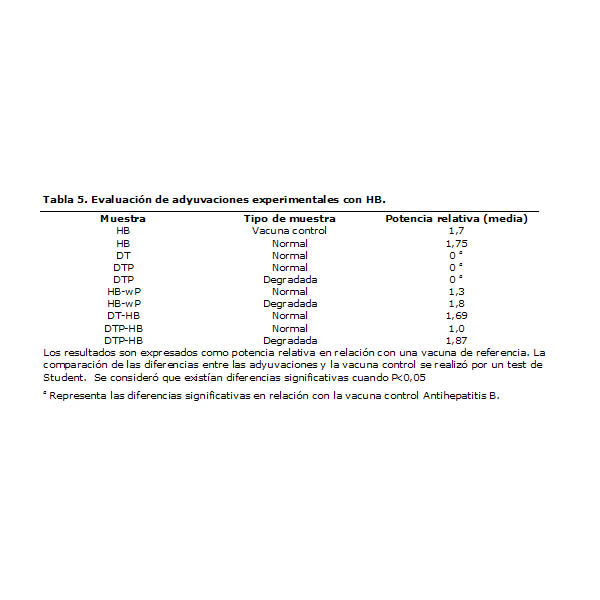
Igualmente evaluamos la potencia in vitro de las adyuvaciones experimentales (Tabla 5), que --como se esperaba--, aquellas que no contenían HB (DT and DTP) no tuvieron valores medibles. Ello demuestra que la reactividad cruzada de wP con la inmunoglobulina HB no es suficiente para rendir un valor de potencia in vitro, además de que demuestra que este es el mecanismo que prevalece y no otro.
Aún cuando no se obtuvieron diferencias significativas, se observó que las adyuvaciones que contenían wP degradado tenían valores de actividad similares a la vacuna control, mientras tanto los que tenían este antígeno de forma íntegra mostraban valores inferiores.
Nuestro laboratorio sólo utiliza para la liberación de lotes de vacunas HB el ensayo de potencia in vitro, razón por la cual debimos analizar el impacto que tiene la interferencia producida por wP. Afortunadamente, todos los lotes de vacunas combinadas con HB mostraron un cumplimiento ampliamente satisfactorio de la especificación definida para este ensayo, por lo que si consideramos que la variabilidad del mismo no excede el 20%, estamos convencidos que la interferencia de wP no es relevante para el proceso de liberación de lotes y se puede utilizar este ensayo en la evaluación de vacunas combinadas con HB sin riesgo de rechazar lotes con actividad biológica satisfactoria y sin necesidad de corregir la especificación.
No obstante, a fin de minimizar la interferencia de wP sobre este ensayo se podría pensar en la inclusión de una vacuna de referencia que combinara wP y HB, aunque se debe tener conciencia de que ello puede entrañar dificultades adicionales. Además, no debe perderse de vista que este es un ensayo para monitorear consistencia de fabricación y no para predecir actividad clínica, de modo que una vez demostrada que la interferencia de uno de los componentes de vacunas combinadas no es relevante, podemos proceder a utilizarlo con seguridad (19).
Conclusiones
Los resultados de este estudio demuestran que el wP es capaz de interactuar de forma estimulatoria o inhibitoria sobre otros antígenos en combinaciones de vacunas, razón por la cual debe disponerse de métodos de ensayo y diseños experimentales adecuados que permitan evaluar de una forma confiable el grado, extensión y naturaleza de esta interacción.
Tabla 5. Evaluación de adyuvaciones experimentales con HB.
Referencias
1. EDQM: Need for Interactivity between OMCLs and licensing bodies. Position Paper September 2004.
2. EMEA: Note for Guidance on Pharmaceutical and Biological aspects of combined vaccines. Committee for Proprietary Medicinal Products (CPMP) 1998.
3. Eskola J. Análisis of Haemophilus influenzae Type b Comjugate and Diphtheria-Tetanus-Pertussis Combination Vaccines. The Journal of Infectious Diseases 1996;174(Suppl 3): S302-5.
4. Mills E, Gold R, Thipphawong J, Barreto L, Guasparini R, Meekison W et al: Safety and immunogenicity of a combined five-component DPT-IPV-Hib vaccine administered to infants at two, four and six months of age. Vaccine 1998;16 (6):576-85.
5. Rennels M, Englund J, Bernstein D, Losonsky G, Anderson L, Pichichero M et al. Diminution of the anti-polyribosylribitol phosphate response to a combined diphtheria-tetanus-acellular pertussis/Haemophilus influenzae type b vaccine by concurrent inactivated poliovirus vaccination. Pediatr Infec Dis J 2000; 19: 417-23.
6. Daum RS, Zenko CE, Given GZ, Ballanco GA, Parikh H, Germino K. Magnitude of Interference after Diphtheria-Tetanus Toxoids-Acellular pertussis/Haemophilus influenzae type b capsular polysaccharide-Tetanus Vaccination is related to the number of doses administered. The Journal of Infectious Diseases 2001; 184:1293-1299.
7. Daum RS, Zenko CE, Given GZ, Ballanco GA, Parikh H, Vidor E et al: Absence of a significant interaction between a Hib conjugate vaccine combined with a DPaT vaccine in the same syringe and inactivated polio vaccine. Pediatr Infect Dis J 2000;19:710-17.
8. Kayhty H, Estola J, Peltola H, Scout MG, Samuelson JS, Gordon L. Immunogenicity in infants of a vaccine composed of Haemophilus influenzae type b capsular polysaccharide mixed with DPT or conjugated to Diphtheria Toxoid. The Journal of Infectious Diseases 1987; 155:100-105.
9. Shams H, Heron I. Mutual interactions between DTaP-IPV Haemophilus influenzae Type b (Hib)-conjugated Vaccines in Laboratory Animal Models. Biologicals 1999;27:227-240.
10. Chovel ML, Pérez LR, Perdomo V, Oviedo M, Costa L. Correlation among physico - chemical and biological tests for a synthetic Hib-TT conjugate vaccine. Vaccine 2007; 25:194-200.
11. Chovel ML., Reyes N: Validation of an in vitro potency test for Cuban Hepatitis B vaccine in the NCL. Development in Biologicals 2002;111:307-315.
12. Council of Europe: Statistical analysis of results of biological assays and tests. European Pharmacopoeia Commission 1997.
13. WHO. Requirements for Hepatitis B vaccines made by recombinant DNA techniques. WHO Technical Report Series 786 1989:38-71.
14. OMS. Normas para las vacunas antidiftéricas, atitetánicas y antipertussis y las vacunas combinadas. Serie de Informes Técnicos de la OMS No. 800, 1990.
15. Vérez-Bencomo V, Santana V, Hardy E, Toledo ME, Rodríguez MC, Heyggnezz L et al. A synthetic conjugate polysaccharide vaccine against Heamophilus influenzae type b. Science 2004 ;305:522-25.
16. Redhead K, Sesardic D, Yost SE et al: Interaction of Haemophilus influenzae type b conjugate vaccines with the diphtheria-tetanus-pertussis vaccine in control tests. Vaccine 1994; 12(15):1460-66.
17. Gupta RK, Anderson R, Cecchini D, Rost B, Xu J, Gendreau K et al. Evaluation of a Guinea Pig Model to assess interference in the immunogenicity of different components of a combination vaccine comprising Diphtheria, Tetanus and Acellular Pertussis (DTaP) vaccine and Haemophilus influenzae type b capsular polysaccharide conjugate vaccine. Biologicals 1999; 27:167-176.
18. Chovel ML, Figueroa JM, Perdomo V. Correlation between potency tests for Cuban recombinant Hepatitis B vaccine. Revista Cubana de Farmacia 2002;1: 66-70.
19. Chovel ML, et al: Comparison between in vitro potency tests for Cuban Hepatitis B vaccine: contribution to the standardization process. Biologicals 2004; 32(4):171-76.
Recibido: Marzo de 2008
Aceptado: Abril de 2008


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}