










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkVaccimonitor
versión On-line ISSN 1025-0298
Vaccimonitor v.17 n.1 Ciudad de la Habana ene.-abr. 2008
Purificación de lipopolisacárido de Neisseria meningitidis a partir de una fracción colateral del proceso de producción de VA-MENGOC-BC®
Purification of Neisseria meningitidis lipopolysaccharide starting from a collateral fraction of the process of the antimeningococcal vaccine, VA-MENGOC-BC® production process
Julio A. Balboa, Jorgelina Estrada, Liliam D. Nápoles, Susan Aguilar, Humberto González, Dairis Hernández , Yisabel Aranguren, Yoanna Garrido, Marlenis Cardoso, Gisela Puentes, Ramón Barberá, Gustavo Sierra.
Instituto Finlay. Centro de Investigación - Producción de Vacunas. Ave.27 No. 19805. La Lisa, Ciudad de La Habana, Cuba.
Correo electrónico: jbalboa@finlay.edu.cu
RESUMEN
El trabajo tuvo como objetivo purificar lipopolisacáridos (LPS) de Neisseria meningitidis a partir de una fracción colateral del proceso de producción de la vacuna antimeningocócica VA-MENGOC-BC®, el sobrenadante que se obtiene del paso de ultracentrifugación durante el proceso de extracción de las proteínas de membrana externa del meningococo. La purificación se realizó mediante precipitación con etanol al 80%, extracción de las proteínas con fenol al 90% entre 65-70 ºC y ultracentrifugación fraccionada a 105,000 g. Se obtuvieron tres lotes de LPS, en total 1,069 g, con un contenido de proteínas, ácidos nucleicos y ácido sálico respecto al LPS de 0,5%, 0,3% y 2,2% (m/m) respectivamente. La evaluación por cromatografía mostró una alta integridad molecular, con valores de constante de distribución reproducibles (0,36-0,38) y una posible asociación del ácido siálico al LPS. Se apreció homogeneidad en el perfil electroforético de los tres lotes y alta actividad endotóxica. El LPS purificado fue identificado fundamentalmente como del inmunotipo L3,7,9. El procedimiento de purificación empleado permite aprovechar una fracción colateral del proceso de producción de la vacuna, es escalable, no incluye métodos cromatográficos, y posibilita la obtención de gran cantidad de LPS de Neisseria meningitidis, no disponible en el mercado, con elevada pureza y alta actividad endotóxica.
Palabras clave: Lipopolisácarido, lipooligosacárido, purificación, Neisseria meningitidis, vacuna antimeningocócica.
ABSTRACT
The work aimed at purifying lipopolysaccharides (LPS) of Neisseria meningitidis from a collateral fraction of the antimeningococcal BC vaccine, VAMENGOC-BC®. production process, the supernatant obtained from the ultra centrifugation stage during the proteins extraction process of the meningococcus outer membrane. The purification was carried out by precipitation 80% ethanol, protein extraction with 90% phenol from 65-70 ºC and fractional ultra centrifugation at 105 000 g. Three lots of LPS were obtained, in total 1.069 g, with a content of proteins, nucleic acids and sialic acid in respect to the LPS of 0.5%, 0.3% and 2.2% (m/m) respectively. The assessment by chromatography showed a high molecular integrity. with constant valves of reproducible distribution (Kd 0.36 - 0.38) and a possible sialic acid association to the LPS. Homogeneity was observed in the electrophoretic profile of the three lots and a high endotoxic activity. The purified LPS was mainly identified as the inmunotype L3,7,9. The purification procedure used allows making use of a collateral fraction of the vaccine production process, it is scalable, it does not include chromatographic methods and makes easy the obtainment of large quantity of LPS of Neisseria meningitidis, wihich it is non available on the market, with high purity and high endotoxic activity.
Keywords: lipopolysaccharide, lipooligosaccharide, purification, Neisseria meningitidis, meningococcal vaccine.
Introducción
El lipopolisacárido (LPS) de Neisseria meningitidis desempeña un papel clave en la enfermedad meningocócica sistémica fulminante y su nivel plasmático en pacientes se correlaciona con la severidad de la enfermedad (1). Está formado por una porción hidrofóbica residente en la membrana externa y altamente endotóxica, denominada lípido A y una porción hidrofílica compuesta por carbohidratos, expuesta a la superficie de la membrana externa y constituida a su vez por una región central o núcleo que se une al lípido A por una cadena de azúcares antigénicamente variable (2). Se divide en once inmunotipos de acuerdo con las diferencias estructurales de la porción sacarídica (3,4), y se ha encontrado que la estructura de la columna de carbohidratos del inmunotipo L3 es idéntica a la de los inmunotipos L7 y L9 (5).
El LPS es también señalado como uno de los principales patrones moleculares asociados a patógenos que son reconocidos por receptores de células del sistema inmunitario innato como señales de peligro, e interactúa con los macrófagos y otras células a través del complejo formado por el receptor semejante a Toll tipo 4, (TLR4, del inglés Toll-like receptor 4) y una proteína accesoria denominada MD-2 (6). Entre los LPS de varias especies de bacterias gramnegativas, el de N. meningitidis resultó ser el más potente agonista del complejo receptor TLR4-MD2 por la vía dependiente del factor de diferenciación mieloide 88 (MyD88, del inglés Myeloid diferentiation factor 88) y el más activo inductor de óxido nítrico por la vía TLR4 independiente de MyD88 (7).
El LPS es un componente del proteoliposoma de N. meningitidis B, uno de los ingredientes farmacéuticos activos de la vacuna antimeningocócica cubana VA-MENGOC-BC® (8), así como de estructuras cocleares derivadas del proteoliposoma (9). Tanto la vacuna como el proteoliposoma y los cocleatos inducen una polarización de la respuesta inmunológica hacia un patrón Th1 (9,10).
Es importante continuar estudiando el papel del LPS, como parte de estas estructuras en la polarización de la respuesta y en la capacidad adyuvante; también es de interés determinar la inducción de anticuerpos anti LPS por la vacuna y su posible acción protectora, así como evaluar su presencia en la Inmunoglobulina Humana Antimeningocócica obtenida a partir del plasma de voluntarios vacunados. Para todo esto se requiere disponer de cantidades suficientes del LPS de N. meningitidis de la cepa Cu385/83, con capacidad antigénica y actividad endotóxica.
El procedimiento más utilizado para la extracción de LPS de N. meningitidis es el método de Westphal (11), basado en la extracción con agua y fenol caliente, después del tratamiento con lizosima (12). Este se obtiene a partir de células (13), o de las vesículas de membrana externa (14).
El sobrenadante que se obtiene del paso de ultra- centrifugación del proceso de producción de VA-MENGOC-BC©, mediante el cual se separan las proteínas que son purificadas por cromatografía para conformar el proteoliposoma de la vacuna, constituye una fuente apreciable, como material de partida, para la obtención de LPS, debido a los volúmenes que de él se obtienen y a su composición. Por ello, resulta de interés práctico y metodológico la purificación de LPS a partir de este material, mediante los métodos usuales de extracción con fenol, cromatografía y ultracentrifugación.
Materiales y Métodos
Extracción y semipurificación de LPS
Se colectó el sobrenadante de ultracentrifugación correspondiente a cuatro procesos, se midió el volumen, se homogenizó y se tomó un litro de cada uno. Se precipitó con etanol al 80% (m/v) y se conservó a -20 ºC durante toda la noche. Se centrifugó a 4,000 g durante 30 min para colectar el precipitado y conservarlo a -20 ºC. Se resuspendió a 4 ºC con agitación, en tampón fosfato de sodio 40 mM, EDTA 5 mM, pH 7, a una relación de 3 mL de tampón por cada gramo de precipitado. Se realizó la extracción de las proteínas con fenol al 90% (v/v), previamente calentado entre 65-70 ºC y se incubó a 6 ºC en un baño termostatado, durante 15 min, con agitación (12). La emulsión fue inmediatamente enfriada en baño de hielo y centrifugada a 10,000 g durante 20 min, para colectar la fase acuosa superior, según lo descrito por Wu, Tsai y Frasch (13). La interfase y parte de la fase fenólica fueron reextraídas con igual volumen de agua. Todo el procedimiento de extracción y reextracción fue repetido una vez. La solución acuosa de LPS recuperada, se precipitó con 10 volúmenes de etanol al 96% (m/v) durante toda la noche a -20 ºC; el precipitado fue obtenido mediante centrifugación a 4,000 g por 20 min y conservado a -20 oC. Posteriormente se resuspendió el precipitado en agua destilada, se ajustó a pH 7 y se trató con ribonucleasa A y desoxirribonuclea A (SIGMA), ambas a 50 g/mL, durante tres horas a 37 oC (13). Se realizó una extracción con fenol frío al 90% (m/v) durante 15 min, se centrifugó a 10,000 g durante 30 min, se colectó la fase acuosa y se dializó contra agua destilada hasta comprobarse la eliminación de todo el fenol mediante lecturas consecutivas de la absorbancia a 260 nm. Se añadió solución de NaCl 5 M para una concentración final de NaCl 0,5 M y se precipitó nuevamente con etanol al 96% (m/v) según fue descrito anteriormente. El precipitado de LPS se conservó a -20 oC.
Procedimiento de purificación por cromatografía
Se descongeló un lote del extracto de LPS semipurificado, se disolvió en 200 mL de agua destilada y se dividió en alícuotas de 50 mL. Se añadió EDTA 2 mM, pH 8,5, se ajustó el pH a 8,5 y se añadió desoxicolato de sodio a una concentración final del 2% (m/v). Se incubó a 37 oC durante 10 min para aumentar la solubilidad y eliminar la turbidez. Se utilizó para la separación por filtración, Sephacril S 300 en una columna de 5 X 100 cm, calibrada previamente con una mezcla de azul dextrana y 2,4 dinitrofenilserina en una relación de 4 mg/0,25 mg, en la solución tampón de elusión (Tris 20 mM, EDTA 2 mM, Deoxicolato de Sodio 10% (m/v) y Azida Sódica 0,2% (m/v), pH 8,5). Se colectaron fracciones de 20 mL y de cada una se tomó una alícuota de 10 mL, se le añadió NaCl 5 M hasta una concentración final de 0,25 M, se precipitó con cuatro volúmenes de etanol al 95%, a -20 oC durante toda la noche, se recogió el precipitado por centrifugación a 4,000 g, 20 min y se disolvió en agua destilada para determinar el contenido de LPS y polisacárido.
Procedimiento de purificación por ultracentrifugación
Se descongeló el extracto de LPS parcialmente purificado, se disolvió en agua y se centrifugó a 10,5000 g durante 8 h (15). El sedimento obtenido fue disuelto y sometido al mismo procedimiento dos veces más.
Controles de la calidad
Contenido de LPS: Se evaluó el contenido de LPS mediante la determinación del ácido 2-ceto-3-deoxioctonoico (KDO) por el método del ácido tiobarbitúrico (TBA) descrito por Osborn (16).
Contenido de proteínas: La concentración de proteínas totales se determinó por el método de Lowry con el empleo de albúmina bovina como patrón, según el procedimiento de Chávez y colaboradores (17, 18) o por espectrofotometría a 280 nm.
Contenido de ácidos nucleicos: Se estimó por adsorción a 260 nm, teniendo en cuenta que 50 g/mL de ácidos nucleicos equivalen aproximadamente a una unidad de absorbancia (14, 19).
Contenido de polisacárido: Se obtuvo mediante la determinación de ácido siálico (AS) por el método del Resorcinol, según Svennnerholm (20).
Evaluación electroforética. Se realizó electroforesis en gel de poliacrilamida con dodecil sulfato de sodio (SDS-PAGE), empleando gel concentrador al 5% (m/v) y gel separador al 15% (m/v). Se utilizó tinción con plata según Tsai Ch y Frasch (21) y Tsai CM y colaboradores (22), con la utilización de dicromato de potasio 3 mM en el paso de oxidación.
Evaluación cromatográfica. Se determinó la constante de distribución (Kd) por cromatografía en columna XK-26/100 con Sephacril-S300 (23). Se aplicó 1 mL de LPS con una concentración de 5 mg/mL y se utilizó un tampón de elusión con pH 8,5; compuesto por Tris 20 mM, EDTA 2 mM, desoxicolato de sodio al 2% (m/v) y azida sódica al 0,2% (m/v). En cada fracción se evaluó la presencia de proteínas y ácidos nucleicos mediante la lectura de absorbancia a 280 nm y 260 nm respectivamente. Para determinar el contenido de LPS y polisacáridos cada fracción fue precipitada previamente en frío con alcohol como se describió anteriormente en la etapa final del procedimiento de purificación por cromatografía.
Evaluación de la pureza química del LPS: Se realizó mediante un sistema de FPLC en Superosa 6 y Sephadex G-25. Para Superosa 6, las muestras de LPS se disolvieron en buffer fosfato de sodio 25 mM, EDTA 2,5 mM y SDS 0,75 mM y pH 8,6. Se ajustó la concentración de la muestra a 2 mg/mL en buffer de elusión y se aplicaron 100 uL con un flujo de 0,8 cm3/min para lograr su elusión. El LPS fue detectado a una longitud de onda de 215 nm. Con Sephadex G 25, las muestras de LPS fueron disueltas a una concentración de 2 mg/mL, en buffer de elusión compuesto por Tris-HCl 50 mM, EDTA 50 mM, DOC-Sodio 1,5% (m/v), pH de 8,5. Se inyectaron 200 l y se eluyó con un flujo de 0,8 cm3/min constante durante toda la corrida. El LPS fue detectado a una longitud de onda de 254 nm y un índice de refracción de 32.
Determinación de inmunotipo del LPS. Se realizó mediante un ensayo inmunoenzimático en fase sólida en placas de poliestireno de alta capacidad de acoplamiento (Costar) recubiertas con Poly-L Lisina, a la concentración de 50 µg/mL, en solución salina tamponada con fosfatos (SSTF) y LPS a la concentración de 2 µg/mL en SSTF. Para la detección se utilizaron los anticuerpos monoclonales anti-L3,7,9 (NIBSC hibridoma stock # 4047), anti-L8 (NIBSC hibridoma stock # 4048) o anti-L10 (NIBSC hibridoma stock # 4049) obtenidos a partir de las líneas de hibridoma W.D. Zollinger 9-2-L379, 2-1-L8 y 14-1-L10, respectivamente, en diluciones de 1/5000. Los anticuerpos monoclonales fueron incubados 2 h a temperatura ambiente, al igual que 100 µL por pocillo de conjugado Proteína A-Peroxidasa (Amersham) en dilución de 1/500. Se añadieron 100 µL de sustrato ortofenilendiamina (OPD) en tampón citrato más peróxido de hidrógeno, se incubó 20 min a temperatura ambiente y finalmente se detuvo la reacción con H2SO4 2N. Se leyó absorbancia a 492 nm en un lector de microplacas Multiskan Plus (Labsystem).
Actividad endotóxica. La potencia del LPS, se determinó por el ensayo del lisado de amebocitos de limulos (LAL), con el juego de reactivos para determinaciones cuantitativas cromogénicas, COATEST Endotoxin (Cromogenix AB) según las indicaciones del fabricante. Este ensayo utiliza un método cinético con un rango de detección para la cuantificación de endotoxinas de 0,005-50 unidades endo-tóxicas por mililitros (UE/mL).
Resultados
Extracción y semipurificación de LPS
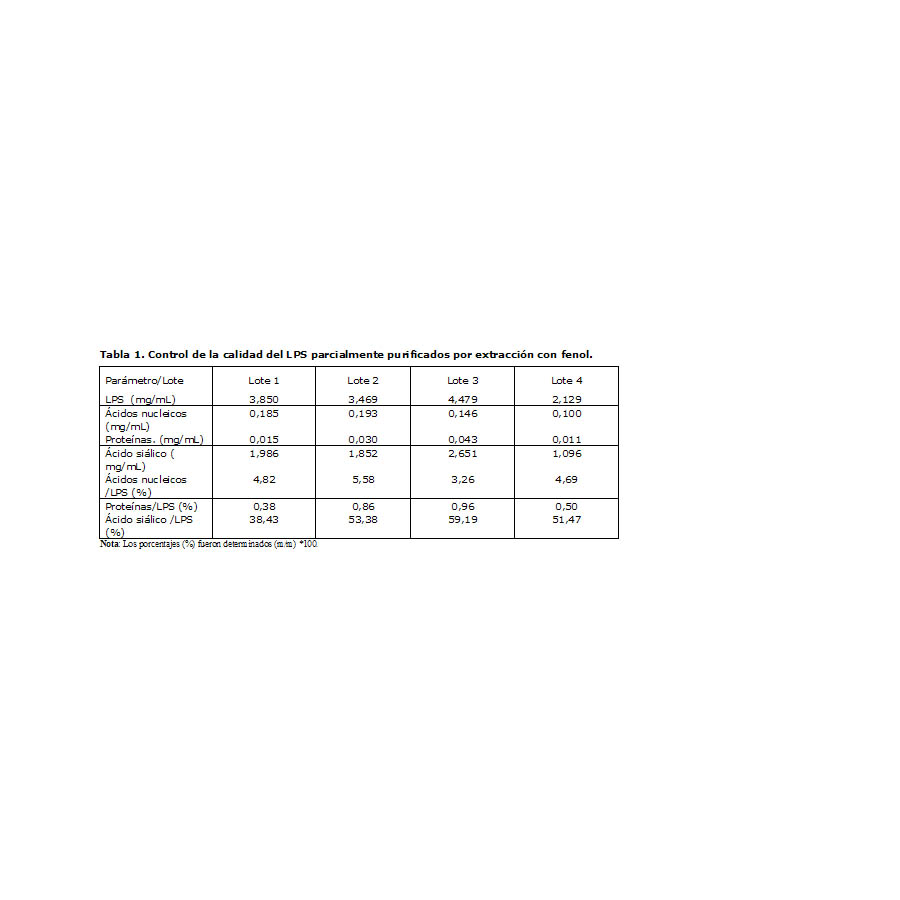
La Tabla 1 muestra los resultados de la purificación parcial de 4 lotes de LPS, a partir del sobrenadante de ultracentrifugación de cuatro procesos de obtención de la vacuna VA-MENGOC-BC®.
Tabla 1. Control de la calidad del LPS parcialmente purificados por extracción con fenol.
El tratamiento enzimático pudo degradar gran parte de los ácidos nucleicos hasta oligonucleótidos y redujo de forma considerable su presencia hasta niveles aceptables para una purificación parcial, con un valor promedio de 4,6% respecto al LPS. Los niveles de proteína fueron inferiores al 1%, lo que demostró la efectividad del método de extracción con fenol para eliminar casi en su totalidad las proteínas contaminantes. El ácido siálico resultó ser el contaminante principal, con un valor promedio de 50,6%.
El análisis comparativo de las fases acuosas de los pasos de extracción y reextracción con fenol (Tabla 2), mostró cómo la reextracción logra recuperar una cantidad elevada de LPS.
La presencia de proteínas fue similar para las dos recuperaciones, y la reextracción no aportó contaminación adicional importante. Sin embargo, la fase acuosa de la re-extracción presentó niveles menores de ácido siálico.
Tabla 2. Control de calidad de LPS obtenidos por extracción o reextracción con fenol caliente.
Purificación por Cromatografía
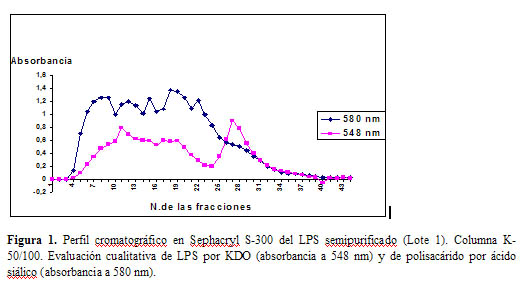
En la Figura 1 se muestra el perfil de elusión cromatográfico del lote 1 de LPS obtenido después de la extracción de proteínas con fenol. Se muestran los resultados de la determi-nación de LPS y de polisacáridos, expresados como valores de absorbancia a 548 nm y 580 nm, respectivamente.
Se observa interferencia entre ambas técnicas, debido a la naturaleza química de ambas moléculas, que están constituidas por unidades de monosacáridos con similitud. Sin embargo, se pudo reconocer en el perfil de elusión detectado a 548 nm, un primer pico casi en forma de meseta y un segundo pico más estrecho y simétrico y mejor definido. Para poder discernir el pico de interés, correspondiente al LPS, se recopilaron las fracciones correspondientes a los dos picos, se centrifugaron y se purificaron por el método de ultracentrifugación. Se evaluó el contenido de LPS, polisacáridos y ácidos nucleicos; antes y después de la centrifugación. Los resultados que aparecen en la Tabla 3, muestran que el segundo pico corresponde mayoritariamente al LPS, con presencia mínima del resto de los contaminantes (ácidos nucleicos y polisacáridos expresados como ácido siálico).
Tabla 3. Caracterización de picos obtenidos por cromatografía en Sephacril S-300 del LPS después de las extracciones con fenol.
Purificación por ultracentrifugación
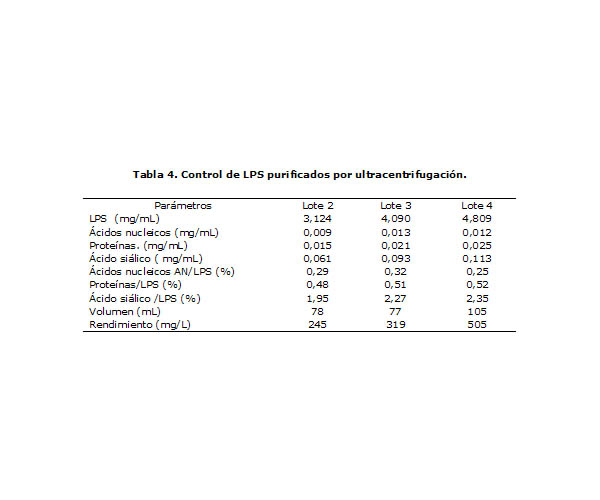
En la Tabla 4 se muestran los resultados del control de calidad de 3 lotes de LPS purificados por ultracentrifugación. Como se puede observar, se obtuvo en todos los casos cantidades apreciables de LPS, con cantidades mínimas de contaminantes (proteínas y ácidos nucleicos < 1% y polisacáridos ˜ 2 %).
Generalmente, para la purificación de LPS se parte de un cultivo, y el recobrado se expresa como la cantidad de LPS purificado a partir de 1g de células de cultivo, en este caso se expresó la cantidad de material purificado como la cantidad de LPS obtenida a partir de 1 L de sobrenadante de ultracentrifugación. En total se obtuvieron 1,069 g de LPS.
Tabla 4. Control de LPS purificados por ultracentrifugación.
Evaluación electroforética
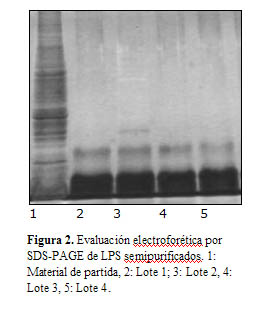
En la Figura 2, se muestra el resultado del análisis por SDS-PAGE de los cuatro lotes de LPS semipurificados y el material de partida de uno de los lotes. Se pudo observar una gran contaminación en el precipitado obtenido del sobrenadante de ultra centrifugación (primera línea) y prácticamente ausencia de estos contaminantes en los 4 lotes de LPS semipurificados.
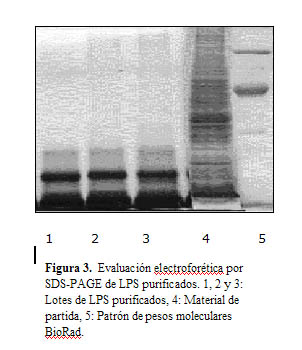
La electroforesis en gel de poliacrilamida con dodecil sulfato de sodio de los lotes de LPS purificados se muestra en Figura 3. Se observa homogeneidad en el patrón electroforético de los tres lotes de LPS, con igual peso molecular. Se observan dos bandas mayoritarias hacia el límite inferior de la electroforesis y una banda de menor intensidad de menor movilidad relativa como igualmente se observó en la electroforesis de los lotes semipurificados.
La densitometría que se realizó a los tres lotes de LPS purificados mostró para estas tres bandas los siguientes resultados: La banda de mayor expresión electroforética fue la de mayor movilidad relativa (Rf) con un valor promedio de 0,90, el porcentaje promedio del área de la banda fue de 58,69% y alcanzó el mayor valor de IR (190), correspondiendo al menor valor de talla molecular, que fue de aproximadamente 7 104,33 Da. El segundo componente presentó una movilidad relativa de 0,75 el cual correspondió a un 18,18 % del área de la banda ocupando un volumen mucho menor que los otros dos componentes y una talla molecular promedio de 9639,33 Da, con el menor valor de IR de 100. El tercer componente fue el de menor movilidad, entre 0,65 y 0,70, con un porcentaje promedio de 23,12%, un valor medio del peso molecular de 11157 Da y una IR de 140. La densitometría fue homogénea para los tres lotes de LPS purificados evaluados.
Evaluación cromatográfica
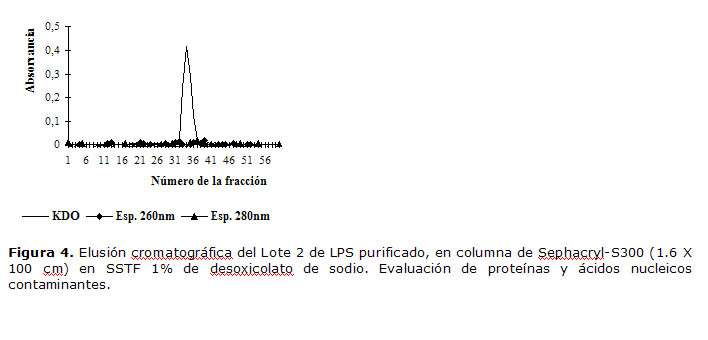
En la Figura 4 se muestra a modo de ejemplo el resultado de la cromatografía en Sephacril S-300 del lote 2 de LPS purificado. Por este método se calculó la constante de distribución mediante la cual se determinó la integridad molecular de los LPS purificados.
Figura 4. Elusión cromatográfica del Lote 2 de LPS purificado, en columna de Sephacryl-S300 (1.6 X 100 cm) en SSTF 1% de desoxicolato de sodio. Evaluación de proteínas y ácidos nucleicos contaminantes.
En el perfil cromatográfico se obtuvo un pico único, simétrico y estrecho, cuyo volumen de elusión fue similar para todos los lotes purificados y evaluados, con un valor de 117,3 mL que correspondió siempre a la fracción número 34, lo que denotó resolución, reproducibilidad y exactitud alta para el sistema de evaluación y por otra parte demostró elevada consistencia en el proceso de obtención y control de la calidad de los lotes purificados. La constante de distribución (Kd) se calculó entre 0,36 y 0,38. Las mediciones espectrofotométricas a 260 nm y 280 nm de las fracciones, permitieron evaluar la contaminación por proteínas y ácidos nucleicos como sustancias en forma de trazas casi indetectables.
Los resultados de la determinación de ácido siálico (polisacárido) en las fracciones mostraron un aumento en la zona del pico correspondiente al LPS, lo cual indica que el ácido siálico podría estar formando parte de su estructura (Figura 5). Los tres lotes de LPS purificado presentaron un comportamiento similar.
Figura 5. Elusión cromatográfica del Lote 2 de LPS purificado, en columna de Sephacryl-S300 (1.6 X 100 cm) en SSTF 1% de desoxicolato de sodio. Evaluación de ácido siálico contaminante.
Evaluación de la pureza química del LPS
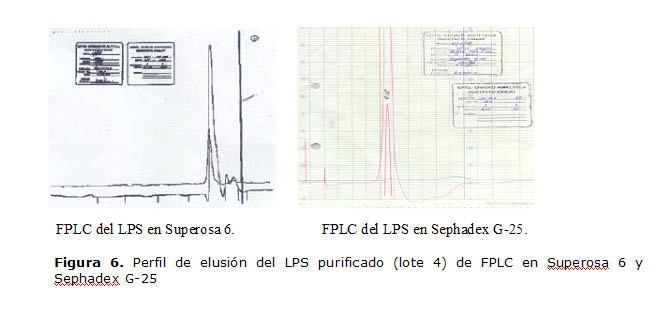
En la Figura 6, se muestran los resultados de FPLC en Superosa 6 y Sephadex G-25 del lote 4 de LPS purificado, la elusión para los otros lotes fue similar. En la cromatografía con Superosa 6 se observó un pico simétrico, con un tiempo de retención de 23,48 min como promedio para todos los lotes. La elusión fue cercana al volumen total de la columna debido al bajo peso molecular que presentan los LPS, no se obtuvo ninguna molécula antes del volumen de elusión del LPS. En la cromatografía en Sephadex G-25, se pudo apreciar un comportamiento similar respecto a la elusión del LPS y no se observó contaminante alguno antes o después. El tiempo de retención promedio fue de 8,53 min, similar para todos los lotes purificados, y no se observó elusión de contaminantes después de la del LPS, cuyo volumen de elusión fue cercano al volumen muerto de la columna.
Figura 6. Perfil de elusión del LPS purificado (lote 4) de FPLC en Superosa 6 y Sephadex G-25
Determinación de inmunotipo del LPS
Mediante el ensayo inmunoenzimático se corroboró que el LPS purificado es fundamentalmente del inmunotipo L3,7,9. La medición con el AcM anti-L3,7,9 fue la mayor.
El ensayo fue negativo para el AcM anti-L10, pero se obtuvo un valor positivo, más de seis veces el valor del blanco con el AcM anti-L8, lo cual indica el reconocimiento también de este inmunotipo aunque en mucha menor cantidad que el L3,7,9 (Figura 7).
Figura 7. Reconocimiento del LPS purificado por los AcM, anti L3,7,9, L8 y L10 (diluciones 1:5000) en el ensayo inmunoenzimático. Se expresa la media y desviación estándar de tres réplicas.
Actividad endotóxicaLos lotes de LPS purificados y liofilizados, mostraron una elevada actividad endotóxica, (Tabla 5).
Tabla 5.Actividad endotóxica por LAL en lotes de LPS purificado.
Discusión
Los cuatro principales componentes en la vesícula de membrana externa son: polisacárido capsular, proteínas, fosfolípidos y LPS; estos se encuentran en una proporción 1:5:2:2 (24). El material de partida utilizado en el presente trabajo difiere de esta proporción, donde el polisacárido aparece en una mayor concentración que las proteínas. Esto pudiera deberse a que en el paso de ultracentrifugación se extraen proteínas que son utilizadas en la producción de la vacuna. Los procedimientos para la purificación de LPS utilizados al inicio (precipitación con etanol y extracción con fenol caliente), no incluyen ningún método que permita eliminar el polisacárido contaminante aunque éste debe haber sido degradado por el calor, por lo cual el LPS parcialmente purificado resultante de las extracciones con fenol tiene una elevada cantidad de polisacáridos.
El paso de reextracción logra aumentar el porcentaje de recuperación del LPS y no aporta contaminación, incluso el LPS que se obtiene, tiene un menor contenido de polisacárido contaminante en comparación con el de la primera extracción; esto se explica por el menor contenido de polisacárido capsular en la interfase debido a su alta solubilidad por ser una molécula altamente hidrofílica.
La cromatografía de exclusión molecular no resultó un método adecuado para la purificación de LPS a partir del sobrenadante de ultracentrifugación en las condiciones empleadas, debido al alto contenido de polisacárido, se requeriría un paso previo para su eliminación. En cambio, mediante la ultracentrifugación, se logró en buena medida eliminar el polisacárido, pues redujo el contenido total de ácido siálico en los lotes purificados a un porcentaje entre 1.9- 2,3 % (m/m) respecto al LPS, mientras que el análisis de las fracciones cromatográficas que se muestran en la Figura 5, indicó que la mayor parte del ácido siálico se encontraba probablemente asociado al LPS, es decir formando parte de su estructura.
El LPS de la cepa que se utiliza en la producción de VA-MENGOC-BC® corresponde al inmunotipo L3,7,9, como se comprobó mediante el ensayo inmunoenzimático con AcM. Los LPS del inmunotipo L3,7,9 asociados a cepas provenientes de enfermos se encuentran usualmente sialilados, mientras que los LPS de cadena corta del los inmunotipos L1, L8 y L10, que se encuentran asociados a cepas de portadores, no están sialilados (25).
Los valores de ácido siálico contaminante pueden además estar sobreestimados debido a la baja especificidad del método empleado que no permite discriminar entre el AS del polisacárido y el LPS, aún así los niveles de AS contaminante son inferiores a los de otros autores. Se han informado valores de hasta el 5% (m/m) de ácido siálico cuando se utiliza el método de extracción con fenol (13, 19). Por otra parte, otros autores han encontrado valores de contaminación de polisacárido que no sobrepasan el 0,3% (m/m) en un LPS purificado solamente por el método de extracción con fenol caliente. Cuando determinaron el contenido de polisacárido B por un ensayo mucho más específico (marcaje isotópico de inhibición de un monoclonal contra el polisacárido B) demostraron que el LPS de N. meningitidis puede contener ácido siálico como parte de su estructura (26). Esto indica la posibilidad de que los niveles de polisacáridos contaminantes en los LPS purificados pudieran ser realmente menores si hubieran sido estimados por un método más específico que la determinación de AS.
Igualmente, los niveles de contaminación con proteínas pudieran estar sobrestimados debido a que las hexosaminas, lípidos presentes en los LPS, pueden generar interferencia en la detección de proteínas (27,28). No obstante los valores de concentración de proteína determinados por Lowry en los lotes de LPS purificado representan sólo el 0,5% (m/m) del contenido de LPS.
Las cromatografías de exclusión molecular de los LPS purificados a baja presión y en FPLC mostraron igualmente el elevado grado de pureza alcanzado.
Además del inmunotipo L3,7,9 que constituye el mayoritario, los resultados de la medición obtenida por el método inmunoenzimático con el monoclonal anti L8, indican la posible presencia de algunas cantidades de este inmunotipo. Andersen (29) obtuvo resultados similares con el LPS que preparó para emplear como antígeno en los inmunoensayos enzimáticos, además del inmunotipo L3,7,9, contenía otros de cadena corta.
El perfil de bandas observado en el análisis electroforético, teniendo en cuenta la presencia de diferentes inmunotipos en el LPS purificado, es similar al obtenido por Tsai, Mocca y Frasch (15), en LPS de N. meningitidis. Ellos caracterizaron por SDS-PAGE que los LPS pertenecían a los inmunotipos L1 a L8 y concluyeron que en general muestran dos bandas mayoritarias bien separadas hacia el final de la corrida junto a otras de menor intensidad con diferencias entre los inmunotipos, las dos bandas principales pueden aparecer como dobletes escasamente separados, que son probablemente la misma molécula con pequeñas diferencias de carga sin embargo, los valores de peso molecular aparente calculados en la electroforesis para el LPS purificado, son mayores que los informados por ellos. Las condiciones de corrida y los patrones de peso molecular utilizados pudieron influir en estas diferencias.
La evaluación electroforética se realizó para comparar el perfil del LPS purificado, mientras que la evaluación cromatográfica por FPLC tuvo como objetivo la evaluación de la pureza. No fue objetivo de este trabajo la caracterización molecular, lo cual sería interesante realizar en trabajos futuros.
La actividad biológica de los LPS purificados, medida por el método de LAL, puede considerarse alta, pero resulta difícil compararla con los resultados de LAL obtenidos por otros autores. En general, existe una correlación moderada entre los métodos cuando el reactivo del LAL es producido por el mismo fabricante e incluso mejor si pertenece al mismo lote, mientras que los resultados pueden ser muy diferentes, hasta para el mismo método, cuando el reactivo proviene de diferentes fabricantes (30).
Wu, Tsai y Frasch informaron valores de actividad endotóxica 10 veces superiores para los LPS purificados por cromatografía que para aquellos obtenidos por extracción con fenol y ultracentrifugación, con valores de concentración mínima para la gelificación de 0,01ng/mL y 0,10 ng/mL respectivamente y apuntaron como posible causa, la diferencia de solubilidad de los LPS purificados por ultracentrifugación y por métodos cromatográficos, superior en estos últimos debido al uso de detergentes (13). Otros autores refieren valores de actividad endotóxica para LPS purificados por técnicas de FPLC que oscilan entre 10 y 17 x 106UE/ng (31). En nuestro caso se obtuvo un LPS con una buena solubilidad.
El método de extracción con fenol brinda la posibilidad de aprovechar las diferencias de solubilidades de los LPS en agua y solventes orgánicos, especialmente para LPS pequeños e hidrofílicos (14). Continua siendo muy utilizado para la extracción de LPS (32), incluso para eliminar la presencia de proteínas contaminantes con actividad endotóxica de preparaciones de LPS comerciales (33). La ultracentrifugación ha sido el método más empleado para la eliminación de contaminantes (34, 35), aunque se ha descrito un aumento en el rendimiento de más del 50% en la purificación de LPS de N. meningitidis con el empleo de la cromatografía de exclusión molecular con respecto a la ultracentrifugación (13), incluso se ha empleado la cromatografía por FPLC con estos fines (31).
En nuestro caso al final de la purificación por el método de ultracentrifugación se logran alcanzar buenos rendimientos, ya que si se tiene en cuenta un volumen promedio de sobrenadante por proceso igual a 3,7 L, así como la obtención de 356 mg de LPS como promedio, a partir de un litro de sobrenadante de ultracentrifugación, se puede calcular la obtención de alrededor de 1,3 g de LPS por proceso.
Los resultados de pureza obtenidos (proteína < 1% y ácidos nucleicos <1%) son similares a los referidos en LPS de otras especies que se comercializan (36).
Conclusiones
Se logró aislar y purificar con elevado rendimiento, LPS a partir de una fracción del proceso de purificación de vesícula de membrana externa de N. meningitidis serogrupo B, que habitualmente se desecha. El LPS obtenido posee alta pureza y actividad endotóxica elevada. Los altos rendimientos y pureza logrados mediante el método de purificación empleado permiten recomendar el uso del sobrenadante de ultracentrifugación como material de partida para la obtención de LPS purificado y demuestran la posibilidad del escalado de este proceso.
Agradecimientos
Los autores agradecemos la revisión crítica de este trabajo al Dr. C. Oliver Pérez así como el apoyo del Dr. C. Luis Riverón y del Dr. Cs. Arturo Talavera.
Referencias
1. Brandtzaeg P, Kierulf P, Gaustad P, Skulberg A, Bruun JN, Halvorsen S, et al. Plasma endotoxin as a predictor of multiple organ failure and death in systemic meningococcal disease. J Infect Dis. 1989;159:195-204.
2. Kulshing VA, Zähringer U, Lindner B, Frasch CE, Tsai CM, Dimitriev BA, et al. Structural characterization of the lipid A component of pathogenic Neisseria meningitidis. J Bacteriol. 1992;174:1793-800.
3. Mandrell RE, Zollinger WD. Lipopolysaccharide serotyping of Neisseria meningitidis by hemagglutination inhibition. Infect Immun. 1977;16:471-5.
4. Zollinger WD, Mandrell RE. Outer-membrane protein and lipopolysaccharide serotyping of Neisseria meningitidis by inhibition of a solid-phase radioimmunoassay. Indect immune 1977;18:424-33.
5. Jennings HJ, Johnson KJ, Kenne L. The structue of an R-type oligosaccharide core obtained from some lipopolysaccharide of Neisseria meningitidis. Carbohydr Res. 1983;121:233-41.
6. Carrillo-Esper R. Inmunidad innata, receptors Toll y sepsis. Cir Ciruj. 2003;71:252-8.
7. Zughaier SM, Zimmer SM, Datta A, Carlson RW, Stephens DS. Diferential induction of the Toll-like receptor 4-MyD88-dependent and -independent signaling pathways by endotoxins. Infect Immun. 2005;73:2940-50.
8. Campa-Huergo C, Sierra-González, VG, Gutiérrez-Vázquez MM, Bisset-Jorrin G, García-Imia LG, Puentes-Rizo, G, et al. Method of producing Neisseria meningitidis B vaccine and vaccine produced by method. United States Patent 1997. Patent number 5 597 572.
9. Pérez O, Bracho G, Lastre M, Mora N, del Campo J, Gil D, et al. Novel adjuvant based on proteoliposome-derived Cochleate structure containing native lipopolysaccharide as pathogen-associated molecular pattern. Immunol Cell Biol. 2004;82:603-10.
10. Pérez O, Lastre M, Lapinet J, Bracho G, Díaz M, Zayas C, et al. Immune response induction and new effectors mechanisms possibly involved in protection conferred by Cuban anti-meningococcal BC vaccine. Infect Immun. 2001;69:4502-8.
11. Westphal, O, Jann K. Bacterial lipopolysaccharides extraction with phenol-water and further application of the procedure. Methods Carbohydr Chem. 1965;5:83-91.
12. Jhonson,KG, Perry MB. Improved Techniques for the preparation of bacterial lipopolysaccharides. Can J. Microbiol. 1976;22:29-34.
13. Wu L, Tsai CM, Frasch CE. A method for purification of bacterial R-Type Lipopolysaccharides (lipooligo-saccharides). Anal Biochem 1987;160:281-9.
14. Gu XX, Tsai, CM. Purification of Rough-Type Lipopolysaccharides of Neisseria meningitidis from Cells and Outer Membrane Vesicles in Spent Media. Analytical Biotech. 1991; 196: 311-8.
15. Tsai CM, Mocca LF, Frasch CE. Immunotype epitopes of Neisseria meningitidis lipopooligosaccharide types 1 through 8. Infect Immun. 1987;55:1652-6.
16. Osborn MJ. Studies on the gram-negative cell in the lipopolysaccharide of Salmonella typhimurium. Proc. Natl. Acad. Sci. USA. 1963; 50: 499-506.
17. Lowry OH, Rosebrough NJ, Farr AL, Randoll RJ. Proteín measurement whith the Folin Phenol reagent. J. Biol. Chem. 1951; 193: 265-75.
18. Chávez MA, Pérez U, Delfín J, Márquez M, Romero M. Manual de Prácticas de Laboratorio de Enzimología. Editorial ENPES. Ciudad de la Habana. 1988[ STANDARDIZEDENDPARAG]
19. Gu XX, Tsai, CM. Purification of Rough-Type Lipopolysaccharides of Neisseria meningitidis from Cells and Outer Membrane Vesicles in Spent Media. Analytical Biotech. 1991; 196: 311-8.
20. Svernerholm, L. Quantitativa estimation of silic acids. II. A colorimetric resorcinol-hydrochloride acid method. Biochim. Biophys. Acta 1957; 24: 604-11.
21. Tsai Ch, Frasch C.E. A sensitive Silver stain for detecting lipopolysaccharides in polyacrilamide gels. Anal Biochem 1982;119:115-9.
22. Tsai CM, Frasch CE, Rivera E, Hochstein HD, Measurements meningococcal protein and polysaccaride preparations for vaccine usage. J Biol Stand. 1989;17:249-58.
23. Chávez MA, Díaz J, Pérez U, Delfín J. Temas de enzimología. Tomo II. Universidad de la Habana, Facultad de Biología 1990:78-80.
24. Zollinger WD, Kasper DL, Veltri BJ, Artenstein MS. Isolation and characterization of a native cell wall complex from Neisseria meningitidis. Infect. Immun. 1972;6:835-51.
25. Jones DD, Borrow R, Fox AJ, Gray S, Cartwright KA, Poolman JT. The lipooligosaccharide immunotype as a virulence determinutosant in Neisseria meningitidis. Microb Pathog. 1992;13:219-24.
26. Mandrell RE, Kim JJ, John CM, Gibson BW, Sugal JV, Apicella MA, et al. Endogenous sialylation of the lipooligosaccharides of Neisseria meningitidis. J Bacteriol. 1991;173:2823-32.
27. Herd, JK. Interference of hexosaminutoses in the Lowry reaction. Anal Biochem. 1971;44:404-10.
28. Eichberg, J, Mokrash LC. Interference by oxidised lipids in the determinutosation of protein by the Lowry procedure. Anal. Biochem. 1969;30:386-90.
29. Andersen SR. Lipopolysaccharide as component in an outer membrane vesicle vaccine against meningococcal disease caused by Neisseria Meningitidis of serogroup B. Thesis submitted for the Degree of Doctor Scientiarum. University of Oslo.1997:24-32.
30. Perdomo Morales R. Ensayo del lisado de amebocitos del Limulus (LAL). Rev Cubana Farm, 2004;38:1-11.
31. Evans JS, Maiden MCJ. Purification of meningococcal lipo-oligosaccharide by FPLC techniques. Microbiology. 1996;142:57-62.
32. Post D, Zhang D, Eastvold J, Teghanemt A, Gibsom BW, Weiss JP. Biochemical and functional characterization of membrane blebs purified from Neisseria meningitidis serogroup B. J Biol Chem. 2005;280:38383-94
33. Hirschfeld M, Ma Y, Weis JH, Vogel SN, Weis J. Cutting edge: Reunification of lipopolysaccharide eliminates signalling through both human and murine Toll-like receptor 2. J Immunol. 2000;165:618-22[ STANDARDIZEDENDPARAG]
34. Hancock I, Poxton I. Separation and purification of surface components. In Hancock I, Poxton I, editors. Bacterial Cell Surface Techniques. Chichester-New York-Brisbane-Toronto-Singapore: John Willey and Sons 1988:91.
35. Tanamoto K-I. Induction of prostaglandin release from macrofages by bacterial endotoxin. Methods in Enzimology. 1994;234:31-41.
36. Catálogo Sigma-Aldrich. 2004-2005:1202-5.
Recibido: Enero de 2008
Aceptado: Abril de 2008


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}