










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkVaccimonitor
versión impresa ISSN 1025-028Xversión On-line ISSN 1025-0298
Vaccimonitor v.17 n.3 Ciudad de la Habana sep.-dic. 2008
Selection of Shigella sonnei strains to develop an effective shigellosis vaccine.
Olga M. Martínez1, Luis Riverón2, Ainel Alemán2, Arturo Talavera2, Isis Montano2, Carmen Soto2, Ranset Diez2, Leopoldo Araujo3, Magaly Suárez2.
1.Centro de Investigaciones Clínicas. Calle 34 Rpto. Coly. Playa, Ciudad de La Habana, Cuba.
2. Instituto Finlay. Centro de Investigación-Producción de Vacunas. Ave. 27 No. 19805 e/ 198 y 202. La Lisa, Ciudad de La Habana, Cuba.
3. Hospital "Hermanos Ameijeiras". San Lázaro y Belascoaín. Centro Habana, Ciudad de La Habana, Cuba.
Correo electrónico: lriveron@finlay.edu.cu
RESUMEN
En el Instituto Finlay se desarrolló una metodología de trabajo que contribuyó a la selección de cepas de Shigella sonnei como posibles candidatos vacunales contra la shigellosis. Las cepas investigadas, donadas por el Centro Provincial de Higiene y Epidemiología de Ciudad de La Habana, se caracterizaron según los métodos convencionales. La identificación del serogrupo y serotipo se realizó por aglutinación en láminas portaobjetos con antisueros comerciales; mientras que para el estudio de la susceptibilidad antimicrobiana se utilizó el DIRAMIC 10, un equipo semiautomatizado que proporcionó los resultados 4 horas después de su realización. Se investigó también la presencia de plásmidos de virulencia, por el crecimiento de Shigella spp. en medio de agar Triptona Soya con Rojo Congo al 0,025%, así como la expresión de las proteínas de la membrana externa en SDS-PAGE; para las pruebas de virulencia y potencia se emplearon los modelos animales (modelo ratón-pulmón y Test de Sereny). Los resultados obtenidos con la metodología utilizada permitieron la selección de la cepa de S. sonnei A-04 como la más adecuada para la obtención del posible candidato vacunal.
Palabras clave: Shigella sonnei, candidato vacunal, modelos animales.
ABSTRACT
A work methodology was developed at Finlay Institute that contributed to the selection of Shigella sonnei strains as possible vaccinal candidates against shigellosis. Strains under study , were donated by the provincial Centre of Hygiene and Epidemiology in the City of Havana and were characterized using traditional methods. The identification and of the serogroup and serotype was performed by agglutination in slides with commercial antisera. Whereas DIRAMIC 10, a semi automated equipment was used to study antimicrobial susceptibility. Results were obtained four hours after. The presence of virulence plasmid for the growth of Shigella spp. In Agar Soy Triptone with Congo Red at 0.025%, as well as outer membrane protein expression in of SDS-PAGE. In addition, animal models mice-lung and the test of Sereny were used for virulence and potency tests. Results obtained allowed the selection of the strain S. sonnei A-04 as the most adequate for the obtainment of a possible vaccinal candidate.
Keywords: Shigella sonnei, vaccinal candidate, animal models.
Introducción
La enfermedad diarreica aguda (EDA), producida por Shigella, es una importante causa de morbimortalidad en los países en vías de desarrollo, lo que representa alrededor del 25% del total de las defunciones que ocurren en los niños menores de 5 años en el mundo (1, 2). La shigellosis es una enfermedad infecciosa cuyo tratamiento con antimicrobianos constituye una medida efectiva para su control, disminuye también la severidad de la infección y previene las complicaciones (3).
Sin embargo, la extraordinaria capacidad que tienen algunas cepas epidémicas de Shigella spp. para crear resistencia a los antimicrobianos, complica la evolución y el tratamiento de la misma (4).
El porcentaje de las cepas multidrogorresistentes (MDR) asciende cada vez más, con variaciones importantes en algunas regiones. La incidencia elevada de cepas resistentes estimula el interés por el desarrollo y obtención de una vacuna segura y efectiva para la prevención de esta entidad clínica (4).
Dentro de la información básica necesaria para acometer una investigación de este tipo es imprescindible el estudio y la caracterización de los serogrupos y serotipos de las cepas de Shigella ssp circulantes, medida basada fundamentalmente en el carácter serotipo específico de la respuesta inmune generada por este microorganismo (5, 6).
Los estudios microbiológicos realizados en Cuba confirman que las especies del género Shigella constituyen un agente etiológico importante de la EDA, con cifras de aislamiento que fluctúan entre 2%-17% y en algunos casos se asocian con complicaciones graves (7).
El presente estudio propone una metodología de trabajo útil para la selección de cepas de Shigella spp, tomando en consideración su caracterización microbiológica, la susceptibilidad frente a los antimicrobianos, la presencia de plásmidos de virulencia, así como la identificación del perfil plasmídico y las pruebas biológicas en modelos animales (modelo ratón-pulmón y Test de Sereny), resultados cuyo análisis servirán como punto de partida para la elección de posibles candidatos vacunales contra la shigellosis.
Materiales y Métodos
Cepas bacterianas
Se investigaron 34 cepas de Shigella spp., 30 se aislaron en Ciudad de La Habana durante los años 1999 y 2000, a partir de cultivos de heces obtenidas de pacientes con el diagnóstico clínico de shigellosis; las cuatro restantes correspondieron a las cepas de referencia utilizadas en este trabajo: Shigella dysenteriae (ATCC 13313), Shigella flexneri (ATCC 12022), Shigella boydii (ATCC 8700) y Shigella sonnei (ATCC 25931).
Identificación fenotípica
Las cepas de Shigella spp. se sembraron en tubos con caldo Triptona Soya, se incubaron entre 18-24 h a 37 0C y posteriormente se inocularon en el medio agar MacConkey, manteniéndose iguales condiciones de temperatura y tiempo de incubación. Al día siguiente las colonias con las características culturales propias del género Shigella (blanquecinas, pequeñas y ligeramente abultadas) se sembraron en agar Hierro Kligler (KIA) y agar Hierro Lisina (LIA); además, se les realizaron las siguientes pruebas bioquímicas y enzimáticas: catalasa, oxidasa, urea, citrato, indol, ornitina, dulcitol, fenil alanina y motilidad.
Caracterización serológica de las especies
La clasificación serológica se realizó por aglutinación en láminas portaobjetos con antisueros comerciales de los siguientes grupos: S. dysenteriae (A), S. flexneri (B), S. boydii (C) y S. sonnei (D), todos producidos por el Centro de Producción de Biológicos "Carlos J. Finlay", de Cuba. Se utilizaron como controles las cepas ATCC, descritas anteriormente.
Prueba de susceptibilidad antimicrobiana
La prueba de susceptibilidad antimicrobiana se realizó por el sistema DIRAMIC 10, un método semiautomatizado y computadorizado, que calculó el índice de inhibición (en porcentajes) que cada antibacteriano fue capaz de producir y en dependencia del valor de esos índices se establecieron los siguientes criterios cualitativos: resistente (R) = 0-70%, intermedio (I) = 71-79% y sensible (S) = 80-100% (8). Los antimicrobianos evaluados y la carga de los mismos se enumeran en la Tabla 1.
Determinación de la presencia del plásmido de virulencia
Para evaluar la presencia del plásmido de virulencia se siguió la metodología descrita por Sasakawa y col. (9). Las colonias aisladas se sembraron en agar Triptona Soya con 0,025% de Rojo Congo (RC) a 37 0C durante 18-24 h; se interpretó como prueba positiva aquella donde se observó el cambio de coloración de las colonias de aspecto blanquecino a colonias rosadas y rojas.
Determinación de la expresión de las proteínas de membrana externa (PME)
El extracto de las PME se obtuvo según la metodología descrita por Frasch (10) y la concentración de las mismas se determinó por la técnica de Lowry (11), utilizándose como estándar Sero Albúmina Bovina (BSA). Las muestras a investigar se prepararon en condiciones desnaturalizantes con 2ß-Mercaptoetanol, posteriormente se les realizó una electroforesis en gel de poliacrilamida con SDS (SDS-PAGE) y, finalmente, las bandas se visualizaron después de teñir los geles con Azul Brillante de Comassie (12).
Pruebas Biológicas
Test de Sereny: También conocida como prueba de la keratoconjuntivitis purulenta en curieles, se realizó según la metodología descrita en la literatura (13), cumpliéndose las regulaciones bioéticas establecidas para los ensayos con animales.
El saco conjuntival de uno de los dos ojos de los curieles utilizados se inoculó con 50 µl de una concentración bacteriana de 7x107-109 UFC; el otro ojo se dejó como control y se le aplicó una solución estéril de Cloruro de Sodio al 0,85%. El grado de keratoconjuntivitis se determinó tomando en cuenta el tiempo que demoró la aparición de los síntomas, así como la severidad y regresión de los mismos. Se consideró la intensidad de la inflamación y se incluyeron valores que oscilaron desde cero hasta tres cruces (+++) (13).
Inoculación intranasal de ratones: Se emplearon ratones Blab/c machos de 18 semanas de edad y con un peso entre 20-25 g, todos obtenidos en el Centro Nacional para Animales de Laboratorio de Cuba (CENPALAB). Los ratones se anestesiaron por vía intraperitoneal con Pentobarbital Sódico y se les inoculó 30 µl de una concentración bacteriana de 5x107-109 UFC de las cepas a investigar (14).
Procesamiento estadístico
El análisis estadístico se realizó mediante el paquete STAT GRAPHICS PLUS (Versión 2.1) a través de las pruebas de comparaciones múltiples de Duncan, con un 95% de nivel de confianza. El análisis de correlación entre los parámetros, la presencia de plásmido de virulencia y la expresión de PME se realizó a través de un análisis de regresión múltiple contenido en este paquete.
Resultados y Discusión
Las cepas aisladas de los pacientes con shigellosis mostraron las mismas características morfológicas, culturales y bioquímicas que las de referencia. En el medio agar MacConkey crecieron colonias pequeñas y blanquecinas, similares a las descritas para el género Shigella (15). Al microscopio óptico todas las cepas se correspondieron con bacilos gramnegativos, rectos y pequeños, corroborándose también la ausencia de movilidad.
En el medio de KIA y LIA se constató la imagen típica de Shigella, características que coincidieron con las observadas en las cepas de referencia. En la prueba de KIA todos los aislamientos mostraron un color amarillo en el fondo del tubo, al igual que en el medio de LIA, la cuña mantuvo su color rojo y el fondo se tornó amarillo. No se observó producción de gas ni H2S.
Todas las cepas fueron catalasa positiva y oxidasa negativa, no utilizaron el citrato ni el dulcitol como fuente de carbono y no hidrolizaron la urea. Además, fueron fenil alanina negativas, ratificándose la ausencia de la enzima fenil alanina de aminasa, así como la no-formación de indol. Todos los resultados se correspondieron con el patrón bioquímico del género Shigella (2).
Nueve cepas fueron ornitina descarboxilasa positiva y el resto negativas, prueba que indicó la presencia de S. sonnei o S. boydii serotipo 13, únicas especies donde es positiva. Al realizar la caracterización de especie, 21 cepas se aglutinaron frente al antisuero de Shigella grupo B y nueve lo hicieron con el D.
No se observó reacción con los antisueros correspondientes al resto de las especies, confirmándose que aquellas cepas que poseían la enzima ornitina descarboxilasa, pertenecieron a la especie S. sonnei.
Susceptibilidad antimicrobiana
Se evaluó el comportamiento de las cepas frente a 10 antimicrobianos. Se observaron aislamientos resistentes a la ampicilina (36,66%), tetraciclina (73,33%), trimetoprim-sulfametoxazol (76,66%) y estreptomicina (63,33%). Sin embargo, se constató buena sensibilidad al cloranfenicol (100%), rifampicina (93,33%, kanamicina (96,66%) y todas fueron sensibles al ácido nalidíxico. Frente a la ciprofloxacina se observó una cepa con sensibilidad intermedia (8,33%) y otra resistente a la ceftriaxona (3,33%).
Plásmido de virulencia
El crecimiento de las cepas en las placas de agar Triptona Soya con 0,025% de RC identificó los fenotipos invasivos y virulentos (colonias que tomaron un color rojo), así como las no invasivas o avirulentas (colonias que mantuvieron su color blanquecino), comportamiento que se relacionó con la pérdida del fenotipo de virulencia acompañado de las alteraciones moleculares en estos plásmidos (9).
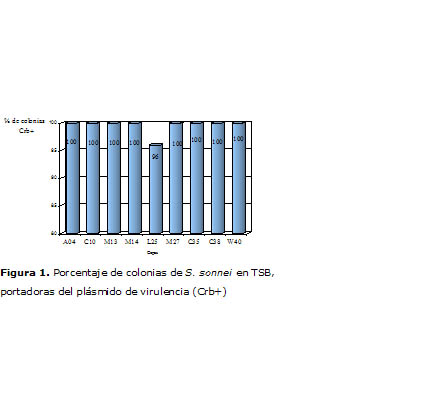
Se observó un porcentaje elevado de cepas que mantuvo su condición de portadoras del plásmido de virulencia íntegro y funcional; no obstante, en el caso de S. sonnei, la cepa L25 manifestó un 96% de prevalencia del plásmido, aunque sin diferencias estadísticamente significativa con respecto al resto de la misma especie (Figura 1).
Determinación de la expresión de las PME
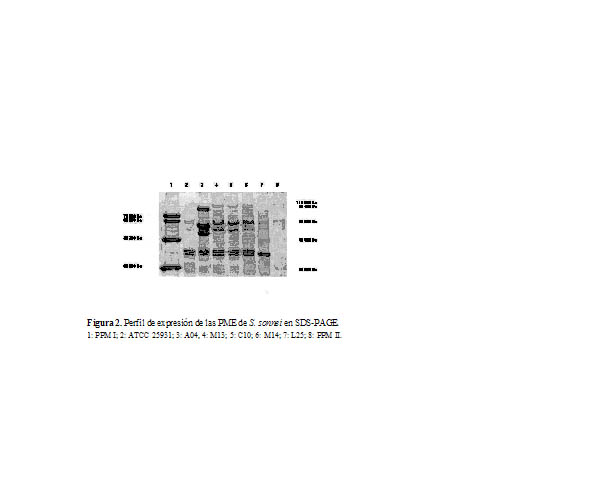
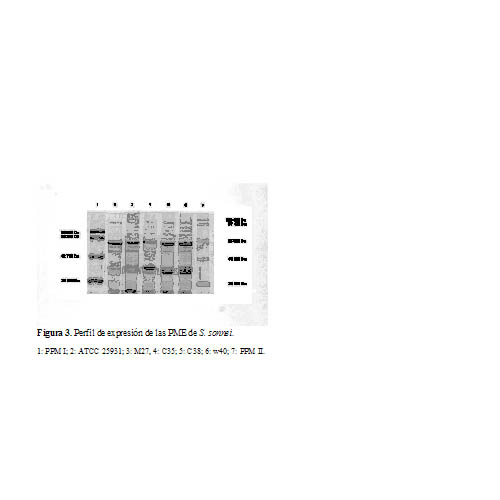
A través de la corrida electroforética en SDS-PAGE se observó el perfil de las proteínas de membrana externa mayoritarias (PMEMs) de cada cepa de S. sonnei (Figuras 2 y 3). Se observó una banda proteica de baja intensidad relativa de expresión, banda que migró a la altura de los 29 KDa y que pudo corresponder con la de 30 KDa.
Cuando se analizó la densitometría de cada una de las bandas del patrón, se identificó el peso molecular (PM) de las proteínas, así como su porcentaje con respecto al total y la intensidad relativa (IR). Al comparar los diferentes niveles de expresión de las PMEMs en las cepas estudiadas se seleccionaron aquellas que mostraron los valores más altos de IR y que se conservaron de manera homogénea, seleccionándose de S. sonnei las bandas con los siguientes PM: 35 KDa, 38 KDa, 58 KDa, 65KDa y 101 KDa.
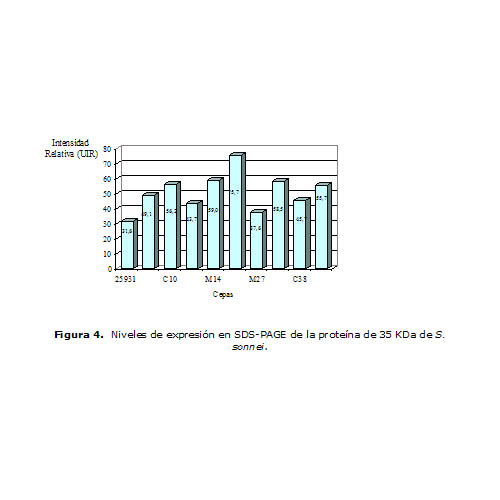
En la Figura 4 se presentan los valores de IR asociados con la expresión de la proteína que migró a la altura de 35 KDa. Entre las cepas de S. sonnei estudiadas, la cepa L25 expresó esta banda con mayor intensidad, mientras que en la cepa M27 la expresión fue menor.
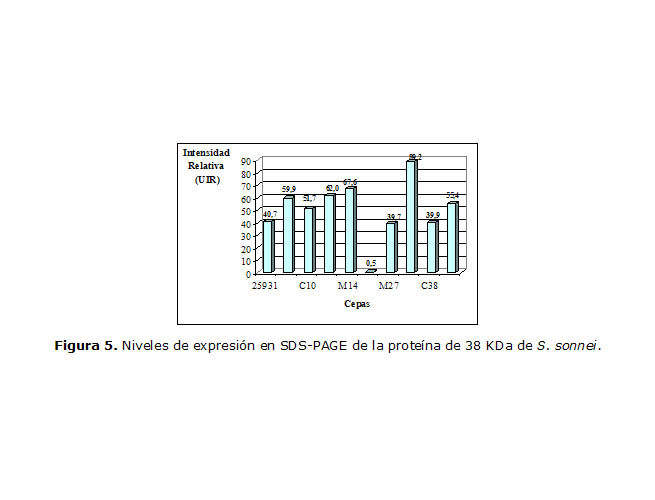
Para la proteína que migró a la altura de las 38 KDa (Figura 5) hubo resultados inversos a los anteriores. La cepa L25 mostró la banda menos intensa y la cepa C35 tuvo una mayor expresión.
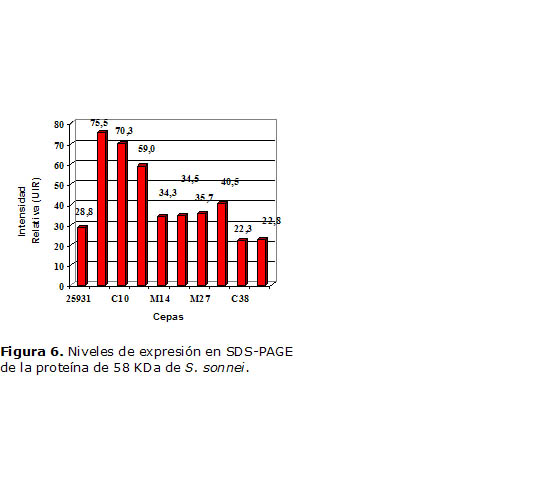
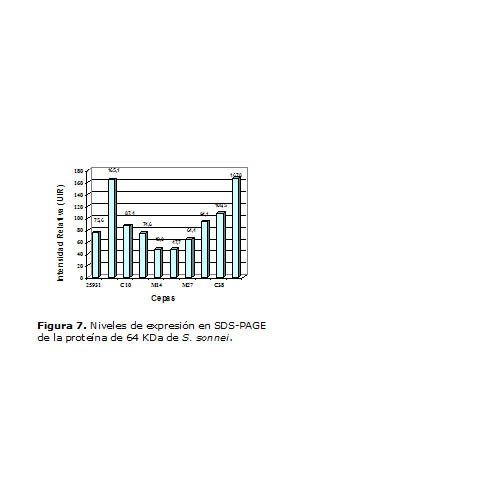
Para la proteína 58 KDa (Figura 6) las cepas A-04 y C10 mostraron los mejores niveles de expresión, mientras que la proteína que migró en el entorno de los 64 KDa (Figura 7) en la cepa A04 presentó nuevamente los mayores niveles de expresión, junto con W40.
Para determinar en cada una de las cepas estudiadas el mayor rango de secreción de estos antígenos se evaluó a través de IR la integración de todos los niveles de expresión, con el objetivo de obtener potenciales candidatos vacunales, sobre todo de cepas para utilizar en la obtención y purificación de vesículas de membrana externa o proteoliposomas bacterianos.
El promedio de los valores de IR asociados con la expresión de las 5 proteínas analizadas en las cepas de S. sonnei A-04, W40, C35, C10, M13, C18 y M4 (Figura 8), no presentaron diferencias estadísticas significativas (p<0,05), presentando los mayores niveles de síntesis, elemento importante para proponer las cepas A-04 y W40 como posibles cepas vacunales.
Se incluyó también el análisis de la proteína de 101 KDa de S. sonnei cuya mayor expresión se logró en la cepa A04 (Figura 9). Aunque los niveles alcanzados para esta proteína no fueron elevados cuando se comparó con el resto, se decidió mantenerla en el estudio por su posible papel en la inmunogenicidad.
Pruebas biológicas
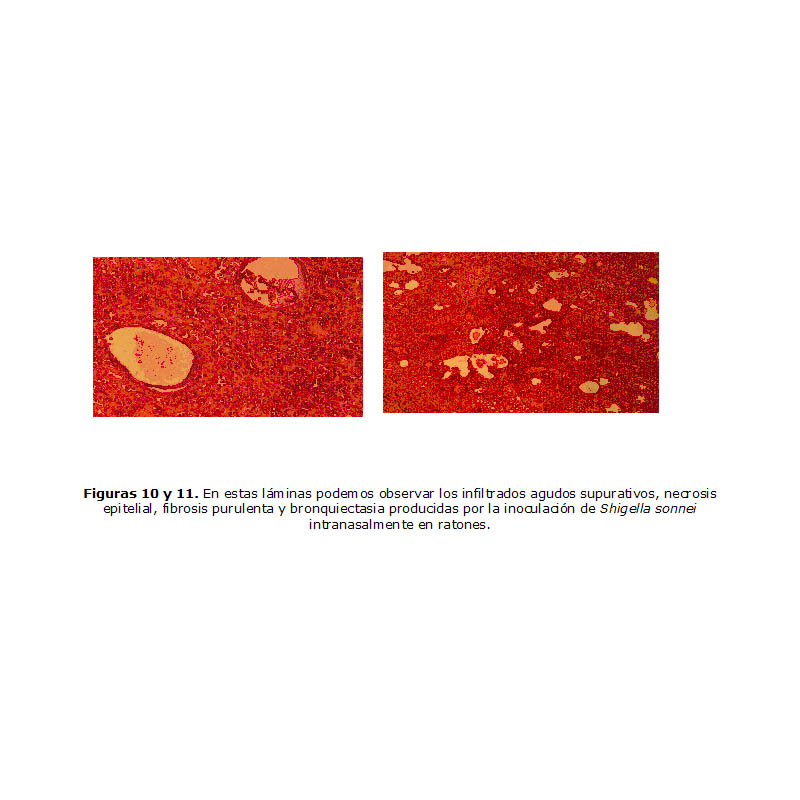
Al realizar el Test de Sereny, la cepa A-04 ratificó su virulencia por producir una keratoconjuntivitis clasificada de tercer grado (+++) (13) y en el modelo ratón pulmón, los animales inoculados presentaron la sintomatología descrita caracterizada por: decaimiento, disminución de su actividad, pérdida de apetito, fiebre, erizamiento de los pelos, hasta el shock séptico mortal. En el estudio histopatológico de los ratones se observaron hemorragias, bronquiectasias, infiltrado inflamatorio con polimorfonucleares y abundantes depósitos de fibrina (Figuras 10 y 11), resultados que sugirieron a la cepa A-04 como la más adecuada para su selección.
Figuras 10 y 11. En estas láminas podemos observar los infiltrados agudos supurativos, necrosis epitelial, fibrosis purulenta y bronquiectasia producidas por la inoculación de Shigella sonnei intranasalmente en ratones.
La EDA, ocasionada por Shigella en los países desarrollados o en vías de desarrollo, tiene dos aspectos importantes: el clínico y el epidemiológico. El primero se relaciona con el comportamiento de la morbimortalidad y el segundo con la distribución de los serogrupos y serotipos presentes en diferentes áreas geográficas. La circulación de los serogrupos parece estar relacionada con el nivel socioeconómico de las comunidades, observándose que en la mayoría de los países vías de desarrollo predominan las cepas de S. flexneri y S. sonnei (19).
Aunque en este trabajo el número de cepas que se estudiaron fue bajo, los resultados coinciden con los notificados por el Laboratorio Nacional de Referencia de Enfermedades Diarreicas Agudas del Instituto "Pedro Kouri" (IPK). En 1998, al evaluar cepas investigadas durante cinco años, esa Institución señala 62,5% y 29,03% de S. flexneri y S. sonnei, respectivamente (20, 21).
Las pruebas de caracterización utilizadas en este estudio fueron suficientes y adecuadas para confirmar la identidad de las cepas candidatas, no fue necesario emplear pruebas bioquímicas adicionales.
Se empleó, por primera vez, para determinar la susceptibilidad de Shigella spp. frente a diferentes antimicrobianos el sistema DIRAMIC 10 en un estudio de investigación. Este sistema calcula el índice de inhibición que produjo cada fármaco, con la ventaja de hacer el diagnóstico en 4 h, disminuyéndose el período requerido para la lectura de los resultados. Se observó un porcentaje elevado de cepas de Shigella resistentes frente al trimetroprim-sulfametaxazol. En Argentina, Uruguay y España aunque no emplearon el DIRAMIC 10, los resultados obtenidos por el método de Kirby-Bauer fueron similares (22).
El cambio de coloración de las colonias en el medio de agar Triptona Soya con RC al 0,025% permitió la identificación de los plásmidos de virulencia. Algunas cepas de S. flexneri mostraron cifras de plásmidos que desde el punto de vista estadístico fueron significativamente diferentes al resto de las cepas de esta especie. Sin embargo, en el caso de S. sonnei, el comportamiento fue favorable.
Es importante señalar que, a diferencia del experimento de estabilidad desarrollado por Maurelli (23), las cepas de este estudio se mantuvieron en un medio de conservación sólido, no se partió de una colonia seleccionada como RC (+) sino de un conjunto de células entre las que pudieron estar incluidas algunas RC (-). Con estas condiciones se conservó la presencia del plásmido de manera aceptable, corroborándose que aún partiendo de una biomasa determinada, al analizar el cultivo total, parece baja la pérdida del plásmido de virulencia.
El análisis de las cepas de S. sonnei se basó en sus características fenotípicas, su susceptibilidad a los antimicrobianos, así como en la presencia del plásmido de virulencia y la expresión de las PME, aunque esta última fue la más discutida. La presencia de algunas bandas importantes en la imunogenicidad y protección de Shigella fueron las que condujeron a la selección final de la cepa propuesta.
Como se explicó anteriormente, la expresión de las PME ayudó a seleccionar la cepa A-04 como la candidata para la obtención de una vacuna. Esta cepa, además de cumplir con todos los criterios de los prototipos vacunales, no presentó diferencias estadísticamente significativas entre los valores de IR; no obstante, mostró un elevado nivel de expresión de la proteína 58 KDa, incluida en un interesante estudio donde señalan a las cepas virulentas de S. flexneri y que crecen en presencia de RC con un nivel mayor de secreción de proteínas con PM de 43, 58 y 63 KDa (23), antígenos con un alto reconocimiento en sueros de humanos y monos.
Se debe señalar que dentro de las cepas de este trabajo, la cepa A-04 coincidió con los resultados obtenidos en la expresión de las proteínas de 58 y 63 KDa (24).
No pueden pasar desapercibidas las proteínas de alto PM, estas constituyen antígenos importantes para la obtención de preparados vacunales de proteoliposomas, ya que está bien documentada su eficiencia en la presentación antigénica a las células inmunocompetentes y el desencadenamiento de la inmunidad adaptativa (24). Entre las proteínas descritas, las codificadas en el segmento génico ipa del plásmido de virulencia de 200 kb de Shigella, tienen un papel protagónico en los procesos de invasividad, así como en el impacto inmunogénico, motivo por el que se decidió poner dentro de los patrones de PM el de 101 KDa, pudiendo diferenciar a la cepa A-04 como la de mayor expresión.
Para estudiar la shigellosis se desarrollan dos modelos de animales, ambos proporcionan una información importante a pesar de sus limitaciones intrínsecas, restricciones dadas porque hasta estos momentos, la infección natural con Shigella no existe en los animales comunes de experimentación y es solo en el mono donde se reproducen los signos y síntomas de la enfermedad. Los modelos empleados: ratón-pulmón y el Test de Sereny, permitieron evaluar el diagnóstico y la sintomatología de la infección por Shigella con sus características propias, esclareciendo la selección de la cepa y su correspondencia con el criterio de virulencia fundamental establecido para la selección de una cepa vacunal.
Sobre la base de todos los criterios y resultados referidos, así como la estabilidad del plásmido de virulencia manifestado a través de su determinación o presencia en el momento de la extracción proteica, se propuso que la cepa A-04 se utilizara como cepa vacunal para la obtención de un proteoliposoma con potencialidad de antígeno vacunal contra la shigellosis.
Referencias
1. Mota MI, Varela G, Godea MP, Caffer MP, Sirok A, Sahelotto F. Serotipos, perfil plasmídico y antibiotipos de cepas de Shigella flexneri aisladas en niños menores de 5 años con diarrea sanguinolenta. Usuarios de los servicios de Salud Pública. Rev Med. Uruguay 2005;21:30-36.
2. Brooks GF, Butel JS, Morse SA. Medical Microbiology, 23 rd edition. McGraw-Hill/Appleton and Lange; 2004.
3. Lépori Luis R. Infecciones bacterianas, miniatlas. Informédica. Contenidos Médicos. Barcelona España; 2007.
4. Gilbert DN, Moellring RC, Eliopolos G, Sande M. The Sandorf Guide to Antimicrobial Therapy 2004. 34 edition, 2004.
5. Paswell JH, Harley E, Ashkenazi S, Chu C, Miron D, Ramón R. Safety and inmunogenicity of improved Shigella O-specific polysaccharide-protein conjugate vaccines in adults in Israel. Infect Inmun 2001;69(3):1351-1357.
6. Fries LF, Montemarano AD, Malelet CP, Taylos DN, Hale TL, Lowell GH. Safety and imminogenicity of a proteosome-Shigella flexneri 2a lipopolysaccharide vaccine administered intranasally to healthy adults. Infect Inmun 2001;69(9):5230-5234.
7. Ramírez M, Valdés N, Bravo L, Fernández A, Castañeda N. Perfil plasmídico y resistencia antimicrobiana en cepas de Shigella aisladas en Cuba. Rev. Cub Med Trop 2004;56(3):178-185.
8. Álvarez-Varela E, Espino-Hernández M, Contreras-Alarcón R, Álvarez-Pineda AB. Evaluación de la resistencia a los antimicrobianos por el sistema DIRAMIC. Rev Panam Infectol 2005;7(4):28-32.
9. Sasakawa C, Kamata K, Sakai T, Murayama SY, Makino S, Toshikawa M. Molecular alteration of the 140-megadalton plasmad associated with loss of virulence and Congo Red binding activity in Shigella flexneri. Infect Immun 1996;51:470-475.
10. Frasch CE. Development and importance of meningococcal serotyping. Division of Products. Office of Biologics Research and Review, FED. Bethesda, p. 42 (trabajo inédito), 1985.
11. Lowry OH, Rosebroug NJ, Farr AL, Randall RJ. Protein measuren twith the folin phenol reagent. J Biol Clin1951;193: 265-275.
12. Laemmli VK. Métodos de laboratorio para el diagnóstico de V. cholerae. Programa Especial de Publicaciones. Washington DC. Estados Unidos. Nature 1970;227:680-685.
13. Sereny B. Experimental keratoconjuntivitis shigellosa. Acta Microbiol. Acad. Sci. Hung. 1957;4:367-376.
14. Noriega FR, Losonsky C, Lauderbaugh F, Liao M, Wang MS, Levin MM. Engineered ?gua BA, ?vir G Shigella flexneri 2dª strain CVD 1205: construction, safety, immunogenicity and potencial efficacy as a mucosal vaccine. Infect Immun 1996; 64:3055-3061.
15. Zwadyk P. Enterobacteriacea: Salmonella and Shigella. Intestinal Pathogens. En: Joklik WK, Willet HP, Amos DB and Wilfred CM. Zinsser Microbiology 20 th edition Appleton & Lange, Norwalk, Connecticult/San Mateo, California, USA; 1992: 556-565.
16. Brudry B, Kaezarik M, Sansonetti PJ. Nucleotide sequence of the invasión plasmad antigen B and C genes (ipaB and ipaC) of Shigella flexneri. Microb Pathog 1998; 4:345-357.
17. Chen Y, Smith K, Zychlisnk A. A bacterial invasión induces macrophage apoptosis by binding directly to ICE. EMBO J 1996.15: 3853-3860.
18. Mallett CP, Van De Verg LL, Collins HH, Hale TL. Evaluationof Shigella vaccine safety and efficacy in an intranasal challenge mouse model. Vaccine 1993;11:190-196.
19. Herrera S, Cabrera R, Ramírez MM, Usera MA, Echeita A. Use of AFLP, plasmad typing and phrnotyping in a comparative study to asses genetic diversity of Shigella flexneri strains. Epidemiiol Infect 2002; 29:1-6.
20. Ramírez M. Comunicación Personal del Laboratorio Nacional de Referencia de EDA del IPK. Ciudad de la Habana, Cuba; 1998.
21. Ramírez M, Valdés N, Bravo L, Fernández A, Castañeda N. Perfil plasmídica y resistencia antimicrobiana en cepas de Shigella aisladas en Cuba. Rev Cub Med Trop 2004; 56(3):178-85.
22. Rossi MA, Galas M, Terragno R, Tokumoto M, Gualfand L, Caffer MI, et al. Multiple Antibiotic Resistance of Shigella isolates; Results of 2 years (1998-1999) Surveillance in Argentina. Abstracts of International Congress of Infectious Diseases 2000;1:89.
23. Maurelli AT, Blackmon B, Curtis R. Loss of pigmentation of S. flexneri 2a is correlated with loss of virulence and virulence-associated plasmid. Infect Inmun 1984;43:397-401.
24. Broca A. Biology of Microorganims. 1994 7th edition: 456-461.
Recibido: Agosto de 2008
Aceptado: Noviembre de 2008


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}