










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkVaccimonitor
versión On-line ISSN 1025-0298
Vaccimonitor v.18 n.2 Ciudad de la Habana Mayo-ago. 2009
ARTICULOS ORIGINALES
Respuesta inmune mucosal inducida por proteoliposoma y cocleato derivados de N. meningitidis serogrupo B.
Mucosal immune response induced by proteoliposome and cochleate derived from serogroups B N. meningitidis.
Judith del Campo1, Miriam Lastre1, Caridad Zayas1, Reinaldo Acevedo1, Elizabeth González1, Belkis Romeu1, Maribel Cuello1, Osmir Cabrera1, Julio Balboa, Ali M. Sarandi 2and Oliver Pérez1
1 Immunology Department, Research Vice-presidency, Finlay Institute, P.O. Box 16017, Havana, Cuba.
2 Department of Microbiology & Immunology, Institute of Biomedicine University of Gothenburg, Sweden.
email: judithc@finlay.edu.cu
Abstract
Mucosal vaccination offers attractive advantages to conventional systemic vaccination. Most pathogens enter or establish infection at mucosal surfaces. This represents an enormous challenge for vaccine development. Nevertheless, the availability of safe and effective adjuvants that function mucosally is the major limitation. Therefore, we investigated the impact of mucosal immunization with the Neisseria meningitidis B proteoliposome (AFPL1, Adjuvant Finlay Proteoliposome 1) and its-derived cochleate (Co, AFCo1). They contain multiple PAMPs as immunopotentiators and have delivery system ability as well as Th1 polarization activity. Groups of female mice were immunized by nasal, oral, intravaginal, or intramuscular routes with three doses with AFPL1/AFCo1 alone or containing ovalbumin or glycoprotein (g) D2 from Herpes Simplex Virus type 2 (HSV-2). High levels of specific IgG antibodies were detected in sera of mice vaccinated with either route. However, specific IgA antibodies were produced in saliva and vaginal wash only following mucosal delivering. The polarization to a Th1 pattern was confirmed by testing the induction of IgG2a/IgG2c antibody, positive delayed-type hypersensitivity reactions, and gIFN production. Additionally, AFCo1gD2 showed practically no vaginal HSV-2 replication and 100% protection against lethal vaginal HSV-2 challenge. In conclusion, the results support the use of AFCo1 as potent Th1 adjuvant for mucosal vaccines, particularly for nasal route.
keywords: Neisseria meningitidis, Adjuvants, Mucosal vaccines
Introduction
Mucosal surfaces are prominent in the gastrointestinal, urogenital, and respiratory tracts, and provide portals of entry for pathogens (1). There are also important logistic reasons that have made mucosal immunization attractive for public health services, especially in developing countries (2). Currently, a major obstacle to developing a mucosal vaccine in humans is finding a safe and effective adjuvant (2-6). Neisseria meningitidis is a major cause of bacterial meningitis in the human, especially among young children. Meningococcal serogroup B based in outer membrane vesicles (Proteoliposome, PL) vaccine induces protective response (7). An attractive vaccine strategy is therefore to induce immunological response at the mucosal surfaces, with the possibility of blocking microbial invasion, as well as to induce systemic immunity with the capacity of inactivating the microbes. AF (Adjuvant Finlay) PL1, is a nanoparticle and contains multiples pathogen-associated molecular patterns (PAMPs). Additionally, we have transformed AFPL1 into a Cochleate (AFCo1) microparticle, that showed high stability and antigen delivering capacity and that contain the same main components of AFPL1 including their PAMPs (8, 9).
Materials and Methods
PLs were obtained from the outer membranes of N. meningitidis serogroup B Cu 385-83 strain (B:4:P1.19.15; L3,7,9) and were produced on industrial scale under GMP conditions at the Finlay Institute, Havana, Cuba. Cochleate structures were obtained from this PL (8). Female Balb/c or C57BL6 mice were inoculated intranasally (IN), intragástrica (IG). Intravaginally routes (IVag) with AFPL1 or AFCo1 (50 µg per dose per mouse). Mice received three inoculations for mucosal routes, separated by a 7 days interval. For intramuscular (IM) immunization, groups of five mice received two doses of AFPL1 or AFCo1. Specific antibodies titers (IgG, IgA, and IgG subclasses) in sera, saliva, and vaginal secretions of immunized mice were determined by capture enzyme-linked immunosorbent assay (ELISA) (9). The ability of AFPL1 or AFCo1 to induce MIP-a, MIP-1b, gIFN or IL-5 productions was determinate on spleen cells from immunized mice. Concentrations of chemokines (MIP-1a and MIP-1b) or cytokine (IFN-g and IL-5) in the cell culture supernatants were determined using Douset chemokine ELISA kits (R&D Systems, Abingdon, United Kingdom). Challenge virus experiments were conducted as previously reported (10). Six days later, the mice were challenged by an intravaginal administration of 9x104 PFU (200 50% lethal doses [LD50]) of HSV-2 strain 333. Mice were examined daily for vaginal inflammation, neurological illness and death after HSV-2 infection. Following intravaginal HSV-2 challenge, vaginal fluids were collected. The differences between the groups were evaluated by a Tukey multiple comparison test and by a Student's t test using Graph Pad Prism 4 software (CA, USA).
Results
Notably, the IN, or IG immunization of mice with AFCo1 or AFPL1 induce IgA antibodies in saliva and vaginal washes.As expected, IN administration induced measurable levels of specific IgA (p<0.001). Neither of the AFCo1 or AFPL1 for IVag or IM immunized groups had not detectable levels of specific IgA in saliva, but for the vaginal washes the AFCo1 or AFPL1 groups had significant (p<0.05) levels of PL-specific IgA. Also, we examined anti PL IgG antibody responses in sera and vaginal lavages. Mice immunized with AFPL1 or AFCo1 by any routes developed high levels of systemic PL specific IgG antibody response. AFCo1 resulted in stronger anti PL IgG responses (p< 0.001) and longer persistence time in sera (up to 6 month) than AFPL1. (Tab. 1).
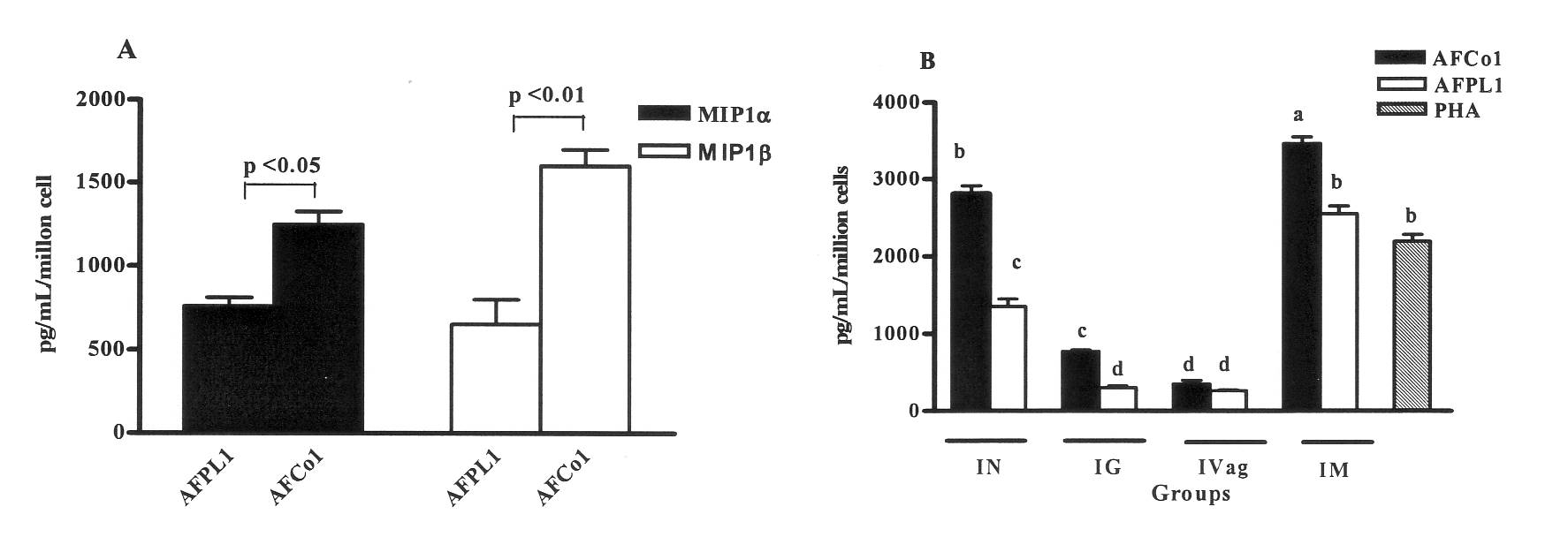
In addition, the presence of high levels of IgG2a in sera using IN, IG, IVag, or IM routes of immunization constitutes indirect evidence of Th1 pattern induction. The data demonstrates that those animals immunized with either AFCo1 or AFPL1 had the good bactericidal activity against the Cu385-83 strain. The sera from the group immunized with the AFCo1 for IN or IM were more effective at killing Cu385-83 serogroup B N. meningitidis strain than were the AFPL1 sera. Next, we evaluated, the MIP-1a levels for both adjuvants were significantly higher responses than control mice groups. Likewise, the MIP-1b production was significantly greater in the AFCo1 or AFPL1 immunized group. Interestingly, the AFCo1 produced higher levels of CC chemokines in spleen cells than AFPL1 (Figure 1A). We determined the gIFN and IL-5 production by spleen cells from immunized mice with AFCo1 or AFPL1 for IN, IG, IVag, or IM, and found these routes produced considerable amounts of gIFN (p< 0.001, p< 0.01, p< 0.05. But no detectable IL-5 compared with those of naive mice (Figure 1B).
This is a direct prove than AFCo1 induce a Th1 pattern. Overall, the AFCo1 immunized mice by IN or IM routes had significantly higher responses than those of the AFPL1 immunized mice or for IG and IVag routes.
The adjuvant capacity of AFCo1 and AFPL1 was also addressed by immunizing mice with a model antigen, a poorly immunogenic soluble antigen such as OVA and a viral antigen as by IVag and IG routes and we compared these results with IN and IM routes. The same result was obtained with either for all the routes immunized mice with OVA incorporated in AFCo1 or AFPL1.
The results showed that IN and IG administration of AFCo1-OVA and AFPL1-OVA also stimulate the production of OVA-specific IgA in saliva and in vaginal washes significantly higher than the groups immunized with OVA alone and enhanced the specific IgG response against OVA for all routes explored and also resulted in sustained systemic (anti-OVA IgG1 and IgG2a) antibody responses in the sera. As well as, we demonstrate for the first time that the application of AFPL1 or AFCo1 by nasal route, with a non replicating gD antigen included, elicited a strong antigen-specific Th1-type cell-mediated immunity and total protection against an otherwise lethal vaginal challenge with HSV-2. Interestingly, the intramuscular only confer a 60% of protection meanwhile the intravaginal route induce.
Discussion
Mucosal surfaces are enormous surface areas that are vulnerable to infection by pathogenic microorganisms. Several investigations indicated a slight reduction in carriage rate among the vaccinated individuals. Mucosal immunization would be a direct approach to stimulating a mucosal response, since the human nasopharyngeal region is the natural habitat for meningococci, and it is believed that nasopharyngeal carriage leads to natural immunization. In the present study we evaluated other mucosal vaccination routes to determine the most effective immune responses to AFPL1 or AFCo1, for this propose we compared different mucosal routes against N. meningitidis antigens on PLs.
This study suggested that IN immunization induced relatively higher levels of mucosal specific IgA and IgG in saliva, vaginal secretions, and sera than IG or IVag immunizations, which support previous results (6) showing that IG route induce potent response in the gastrointestinal-tract tissues and in IVag immunization had a local behavior of the immune response, which could be important for protection against pathogens that colonize vaginal mucosae and which is supported too, by animal studies using other antigens coupled to Cholera Toxin B given by various routes(13). We show here that the local mucosal administration of AFCo1 is associated with a strong increase in the CC chemokines MIP-1a, MIP-1b. In addition, CC-chemokines also link activation of the Th1 adaptive cellular immunity in the tissue to the innate immune response (11, 12). Also, Spleen cell from AFCo1 or AFPL1 immunized mice produced high levels of gIFN.
Table 1. PL specific antibody responses induced by different routes immunized mice. Groups of female Balb/c mice were vaccinated with AFPL1, AFCo1. PL specific IgA or IgG levels in their saliva, sera or vaginal washes, were determined using a specific ELISA. Significant differences between the means of different groups were determined by a Tukey multiple comparison test using the Graph Pad Prism 4 software (Calif.). A P-value of <0.05 was considered statistically significant and it is represent by different letters.
Figure 1. PL-specific cell-mediated immune response. Groups of female Balb/c were immunized with AFPL1, AFCo1. Four weeks after the last immunization, the mice were sacrificed and the spleen cells or purified CD4+ T spleen cells were co-cultured with CD11c+ dendritic cells pulsed with PL protein. (A) Shows PL-specific MIP-1a, MIP-1b contents by ELISA. (B) Shows IFN-g concentration in co-cultures' supernatants. Significant differences between the means of different groups were determined by a Tukey multiple comparison test using statistical analyses with Graph Pad Prism 4 software.
This result clearly indicates that in vivo AFCo1 induce the activation of T cells and their differentiation into Th1 cells. Thus, the presence of gIFN producing T cell clones is direct evidence of dendritic cells presentation of AFCo1 antigens. AFCo1 is a very effective adjuvant that may be useful for the development of such vaccines. The same result was obtained with either route immunized mice with OVA or gD2 incorporated into AFCo1 or AFPL1.
Previous studies have shown IN immunization to be an effective means for the induction of serum and mucosal antigen specific antibodies (11-14). On the other hand, IN route may be an effective alternative that is more convenient and acceptable, has been more promising in terms of eliciting genital-tract antigen-specific IgA and IgG in mice, primates, and human, too can avoid degradation of vaccine antigen caused by digestive enzymes, and so requires a smaller dose of antigen than oral immunization. In addition, the genital-tract antibodies generated as a function of IN immunization have been demonstrated to be long lasting in mice (11-14).
Finally, we observed promising results with IN administration of AFCo1 containing gD2 of HSV for induction of protective immunity against genital herpes, AFCo1gD confers a total protection measured by elimination of virus. It is important to highlight to know if both adjuvants are effective at inducing protection when we compare the classical route of immunization, because all currently vaccine candidate are parenteral vaccines. In summary, AFCo1 or AFPL1 have proved to function very well both as nasal vaccine per se and as a mucosal adjuvant for nasal vaccine such as HSV-2, human immunodeficiency virus, and human papilloma virus.
References
1. He Q, Mitchell A, Morcol T, and Bell SJD. Calcium Phosphate Nanoparticles Induce Mucosal Immunity and Protection against Herpes Simplex Virus Type 2. Clinical and Diagnostic Laboratory Immunology 2002;9:1021_24.
2. Neutra MR and Kozlowski PA. Nature Reviews. Immunology 2006; 6:148-58.
3. Mestecky J, Russell MW, and Elson CO. Perspectives on Mucosal Vaccines: Is Mucosal Tolerance a Barrier?. The Journal of Immunology: BRIEF REVIEW 2007: 5633-38.
4. Johansen FE, Baekkevold ES, Carlsen HS, Farstad IN, Soler D, and Brandtzaeg P. Regional induction of adhesion molecules and chemokine receptors explains disparate homing of human B cells to systemic and mucosal effector sites: dispersion from tonsils. Blood 2005;106:503-600.
5. Alcon V, Baca-Estrada M, Vega-López M, Willson P, Babiuk LA, Kumar P, Hecker R and Foldvari M. Mucosal Delivery of Bacterial Antigens and CpG Oligonucleotides Formulated in Biphasic Lipid Vesicles in Pigs. The AAPS Journal 2005;7 (3) Article 57 (http://www.aapsj.org).
6. Holmgren J, Czerkinsky C, Eriksson K, and Harandi A. Mucosal immunization and adjuvants: a brief overview of recent advances and challenges. Vaccine 2003;21(Suppl 2):89-95.
7. Sierra G, Campa HC, Varcacel NM, Izquierdo PL, Sotolongo PF, Casanueva GV, et al. Vaccine against group B Neisseria meningitidis: Protection trial and mass vaccination results in Cuba. NIPH. Ann. 1991;14.
8. Campa C, Sierra VG, Gutiérrez M M. et al. Method of producing Neisseria meningitidis B vaccine, and vaccine produced by method. United States Patent 1997. Patent number 5 597 572.
9. Pérez O, Lastre M, Cabrera O, del Campo J, Bracho G, Cuello M, Balboa J, Acevedo R, Zayas C, Gil D, Mora N, González D, Pérez R, González E, Barbera R, Fajardo ME, Sierra G, Solís RL, and Campa C. New Vaccines Require Potent Adjuvants like AFPL1 and AFCo1. Scandinavian Journal of Immunology 2007;66: 271-77.
10. Rodríguez T, Pérez O, Menager N, Ugrinovic S, Bracho G, and Mastroeni P. Interactions of proteoliposomes from serogroup B Neisseria meningitidis with bone marrow-derived dendritic cells and macrophages: adjuvant effects and antigen delivery. Vaccine 2005;26:1312-21.
11. S. Tengvall, A. Josefsson, J. Holmgren and A. M. Harandi. CpG oligodeoxynucleotide augments HSV-2 glycoprotein D DNA vaccine efficacy to generate T helper 1 response and subsequent protection against primary genital herpes infection in mice. J Reproduct Immunol 2005; 68:53_69.
12. Singh D, O'Hagan. Advances in vaccine adjuvants. Nat Biotechnol 1999;17:1075_81.
13. J. Holmgrem, C. Czerkinsky. Mucosal immunity and vaccines. Nature Medicine 2005;11(4):45-52.
14. Haneberg B, Dalseg R, Wedege E, Høiby EA, Haugen IL, Oftung F, Andersen SR, Naess LM, Aase A, Michaelsen TE, and Holst J. Intranasal administration of a meningococcal outer membrane vesicle vaccine induces persistent local mucosal antibodies and serum antibodies with strong bactericidal activity in humans. Infect Immun 1998;66: 1334_4.1.


{kind=link}
{kind=link}